Impact of agricultural landscape structure on the patterns of bird species diversity at a regional scale
2024-01-22DenisaDvokovJanipoJosefSuchomel
Denisa Dvoṙáková, Jan ŠipoŠ, Josef Suchomel
Department of Zoology,Fisheries,Hydrobiology and Apiculture,Mendel University in Brno,Zemˇedˇelsk´a 1,613 00,Brno,Czech Republic
Keywords:Bird occurrence Citizen science Conservation Landscape influence Phylogenetic diversity Spatial heterogeneity
ABSTRACTThe loss of bird species diversity is a crucial problem in the European agricultural landscape.Change in the area coverage of major land cover types has been mentioned as one of the main factors responsible for bird biodiversity impoverishment.In this study, we focused on the impact of landscape matrix characteristics on bird species richness and on Faith’s phylogenetic diversity index on a spatial scale of 1000-m radius around the measured occurrence points.We investigated how land cover composition affects bird diversity on the landscape scale using nationwide citizen science data.In total, 168,739 records of bird occurrence in the South Moravian Region of the Czech Republic during growing season from 2009 to 2019 were evaluated.We found that the presence of water bodies and wetlands significantly corresponded to the areas of highest bird species richness.We also revealed that the presence of forests (~60% of the forest in the Czech Republic is occupied by commercial forests), urban areas and arable land were negatively associated with bird species richness and phylogenetic diversity.Forests (both coniferous and deciduous) and urban habitats were found to have a tendency to host a clustered phylogenetic community structure in comparison with wetland and arable land.A strong negative association between forest proportion and bird diversity led us to conclude that the expansion of the forest (with simple species composition, horizontal and vertical structure) could be one of the critical drivers of the decline of bird species diversity in the European agricultural landscape.On the other hand, our results also pointed out that small woody features (i.e., woodlots) and scattered woodland shrub vegetation were one of the main landscape characteristics supporting a bird diversity in rural landscape.This is in concordance with other studies which mention these landscape structures as important elements for nesting and foraging of farmland birds.We thus recommend to maintain and restore scattered trees or woodlots with complex structure in agricultural landscape.
1.Introduction
Bird communities are good indicators of environmental changes that shape biodiversity at the landscape scale (Donald et al., 2001; Gregory et al., 2003; Fraixedas et al., 2020; Šálek et al., 2022).Populations of a large number of bird species have declined since the second half of the 20th century in Europe (Reif, 2013; Burns et al., 2021).The results of many research studies indicate that intensive farming and forestry,landscape abandonment and homogenisation have been the main drivers of the bird decline in the European landscape (Guilherme and Miguel Pereira, 2013; Reif and Hanzelka, 2016; Šálek et al., 2021,2022).Habitat loss caused by agricultural and forest monoculture expansion affect not only alpha but also beta diversity by increasing landscape homogeneity (Tscharntke et al., 2008).The decline in the overall species diversity of birds is also a reflection of the declining population trend among farmland birds driven by land use intensification (Donald et al., 2001; Reif and Hanzelka, 2016).The decline in bird diversity may also have another explanation, that is, changes in the use of agricultural land.Reif and Hanzelka (2016) argue that the changes in the areas of major land cover types may be a major factor in bird population change.
The species richness within given areas depends not only on habitat quality but also on the structure of the surrounding landscape.Connected landscape features are commonly considered together as a landscape matrix (Jonsen and Fahrig, 1997).It is well known that the composition of the matrix is one of the key factors influencing species richness at the regional scale (Hendrickx et al., 2007; Klingbeil and Willig, 2016; Reider et al., 2018).The composition and spatial configuration of the landscape features in the matrix may also influence the effect of habitat heterogeneity on species richness (Hendrickx et al.,2007; Šálek et al., 2018).Agriculture intensification has led to the disappearance of diverse mosaics of fields, grasslands, patchy landscapes and old roads.Scattered greenery has also disappeared from the landscape, and the overall structure of crops has changed to a more homogeneous species composition (Zámeˇcník, 2013).These changes in landscape negatively affect many bird species, such as shrub-rich grassland species (e.g.,Lanius collurio), farmland birds (e.g.,Perdix perdix), forest birds (e.g.,Streptopelia turtur) (Zámeˇcník, 2013) and wetland birds (e.g.,Crex crexandVanellus vanellus) (Cherkaoui and Hanane,2011; Pykal et al., 2021).Landscape elements such as linear patches,hedgerows, headlands, copses and tree rows are important factors in maintaining the complexity and connectivity of agricultural land (Tiang et al., 2021; Anderson et al., 2022).The presence of these areas within a landscape offers the opportunity for higher species diversity and richness (Norton et al., 2009; Morelli, 2013).The benefits to bird species include providing places for nesting, roosting and hiding.Linear and patchy areas also increase the connectivity of the landscape and often allow forest and forest-edge bird species to expand their presence into the agricultural land (Wilson et al., 2017; Jungandreas et al., 2022).In a study by Mulwa et al.(2012), bird density and richness were found to be higher in areas with diverse structural vegetation.Bird richness was also higher in and near natural forest types and on self-sufficient subsistent farmland than in secondary forests and plantations, indicating the type of vegetation also determines the bird species.A study by Šálek et al.(2018) demonstrated that habitat specialist species were most severely influenced by the heterogeneous landscape with higher cover of non-cropped elements and smaller field sizes.Understanding how habitat heterogeneity and composition affect species diversity is crucial for restoring countryside (Tscharntke et al., 2005).
Citizen scientists play a key role in providing the necessary data for species distribution and diversity (Bowler et al., 2022).However, there are still debates about the quality of citizen science data and whether they provide data of sufficient quality to be worth the cost (Ganzevoort et al., 2017).The citizen science differs from each other in the structuring of the data (structured, semi-structured or unstructured) and the knowledge of the observers, which can consequently create bias (Shen et al., 2023).Differences in the detectability of different bird species and variable identification skills among volunteers leads to underestimation of true species richness.Therefore, species richness estimators (i.e.,Chao 1 or 2) should be used to estimate true species richness in a community (Shen et al., 2023).Callaghan et al.(2021) in their work pointed out, that biases exist in unstructured citizen science data when compared with semi-structured data, predominantly as a result of the detectability.Their results indicated that large-bodied birds were over-represented in the unstructured citizen science dataset (Callaghan et al., 2021).Based on these results body size as a fundamental trait should be used in the ecological modelling as a covariate (Callaghan et al., 2021).
In this study we used semi-structured citizen science data to explain the pattern of bird species diversity distribution within an agricultural landscape.In general, we hypothesize that land cover types in an agricultural landscape that increase the diversity of its landscape matrix,while being able to maintain dominant ecological groups of birds (e.g.,farmland specialist or generalist species), will be positively associated with bird richness and diversity.In a landscape dominated by arable land, we assume that the forest patches and edge habitats (e.g., wood lots and headlands) will be important elements in maintaining bird diversity within this landscape context.We based this prediction on the results which revealed that scattered greenery and woodland edges provide important nesting and foraging habitats especially if they are rich in shrubs and deciduous trees for farmland birds (Berg, 2002;Schmidt and Tscharntke, 2005; Sasaki et al., 2020).The accumulation of different ecological groups of species (e.g., edge and open-habitat species) in these patches will also cause an increase in phylogenetically diverse assemblages.We also address the questions: a) how land cover composition influence bird diversity; b) the impact of small woody features in relation to the species richness of birds in the landscape; and c) which types of land cover dominantly affect bird community characteristics and how they affect its structure.
2.Materials and methods
2.1.Study area
Our study focused on an area in the South Moravian Region in the Czech Republic (Fig.1), which is defined by the districts of Blansko,Brno city, the Brno surroundings, Bṙeclav, Hodonín, VyŠkov and Znojmo.The region is divided into 21 administrative districts or municipalities.Overall, it has an area of 718,783 ha and a population of more than 1,195,000.This region ranks fourth place in the republic in terms of area.Due to its geomorphological, soil and climatic characteristics, the region of South Moravia belongs to the warmest regions of the Czech Republic with a large representation of lowlands and fertile black soils.Within the Czech Republic, it is therefore burdened by an intensive agricultural activity (~60% is represented by agricultural land, of which ~84% is arable land).Production of the region’s agriculture is mainly focused on the cultivation of cereals, rape (Brassica napusL.) and maize for green and silage.The region is also specific in that vineyards in the region area occupy 90% of the area in the Czech Republic.The highest share of arable land is in the Znojmo and VyŠkov districts (CZSO, 2021).The area of forest proportion in the South Moravian region represent a 28.3% of region area (MZe, 2022) and a share of 7.5% of the Czech Republic total area forest land (CZSO, 2022).Within the region, there are four types of landscape characteristic.The central part of the region is influenced by the main city of Brno, with the surrounding landscape being influenced by the large urban agglomeration.The northern part of region is defined by cave complexes in the Moravian Karst.The southern part of the region predominantly consists of flat areas of fields, meadows and vineyards, with remnants of flooded forests along the Dyje River.The dominant area at the southwestern tip of the region, Podyjí National Park, is an example of an exceptionally well-preserved river valley in a richly wooded landscape.Many of the water areas by the Dyje River between the cities of Znojmo and Bṙeclav are ideal nesting places for water birds.Beyond the Morava River in the eastern region, the landscape gradually rises into the hills of the White Carpathians.This biosphere reserve is one of the most valuable areas in Europe (CZSO, 2021).The reason why we chose the South Moravian as a region where we studied the effect of the landscape structure is that we wanted to study biodiversity trends in an intensively agricultural landscape, and the South Moravian Region meets this criterion.

Fig.1.Map of the Czech Republic with the locations of study area (South Moravian Region).The map was created in ArGIS Pro software (ESRI, 2021)and the ArcˇCR 500 database used (Arcdata Praha, 2016).
2.2.Species distribution data
To study the patterns of bird diversity we used semi-structured citizen science data which are suitable for modeling species distribution on the landscape scale (Johnston et al., 2020; Callaghan et al., 2021).Semi-structured data means that data included ancillary information on the observation process.The bird data used in this study were collected from the AVIF database of the Czech Society of Ornithology (CSO,2009–2019).Semi-structured data contained at least: precise location,time of day, and name of the observer.To the database can contribute anyone from the public, but the registration to the database is necessary.Most amateur ornithologists with knowledge of observed bird species contribute to the database.Together, we processed 168,739 records on bird presence in South Moravia from 2009 to 2019.The dataset contains information on the presence of individual detected bird species at particular places and times.It also contains additional information about the species, for example, their age, the number of individuals and other geographical information.As mentioned above, we used semi-structured data which was sorted before actual use, when incomplete records were removed.This means, that it was crucial for us to use only complete records with geographic coordinates and with certainty correctly identified bird species.Incomplete records and records with uncertain identification of the observed species were removed.As the dataset also contains information about species that are present in our territory rarely and for a short time, or non-native species, the data have been modified.These species could create a misrepresentation in the results, so their data have been removed.We focused mainly on species that nest in the Czech Republic or remain in the territory for long periods of time.To avoid the effect of bird migration on our results, we included in the analyses the period between May and September, during which most of the birds had already returned from their overwintering locality.It also means that the analysis did not include the winter season, during which bird communities are dominated by mainly overwintering species.
We used the Atlas of Breeding Birds in the Czech Republic to divide species between trophic and migratory guilds (Šťastný et al., 2021).We placed the birds whose food consisted partly of vegetable food and partly of protein food of either animal or insect origin in the omnivore trophic guild.For other specific trophic guilds, such as carnivore, herbivore, seed eater and insectivore, we considered the components that mostly predominated in the birds’ diets (component that prevailed over 70% of diet).
Since the data obtained from the faunistic database are public, semistructured citizen science data, they can be burdened with certain errors and distortions, which can be generated in different ways.This includes,in particular, the characteristics of the individual observed bird species(e.g., size–large bird species have a greater chance of being noticed).Furthermore, different field efforts and pseudo-absences (e.g., some habitats are more interesting to observers and may be preferred for observation–wetlands, large bodies of water).Furthermore, the size of the area of individual landscape covers (the number of species increases with the size of the habitat area) and the season (arrival and departure of migrating individuals).Therefore, before the data were processed by using statistical models, it was necessary to adjust them.In order to filter out the bias arising from the different detection properties of the species(i.e., the effect of body size), the average body size of the recorded bird species for each cadaster area appeared as a covariate in the models.Information on the size of individual bird species was obtained from Šťastný et al.(2021).Another negative effect of distortion resulting from the positive dependence between the number of species and the cadaster area (i.e., larger land covers will have the potential to host more species)was solved by using the logarithm of the area size as covariate.Variable identification skills among volunteers can cause bias during data collection due to different detectability of species.Therefore, volunteers with variable detection skill level may miss hard-to-detect species.We therefore applied the ‘Chao 2 estimator’, which is used to estimate the total potential number of species at study sites (Chao, 1987).Among non-parametric approaches, the Chao estimator was found to be the least biased and is able to improve estimates of semi-structured data(Shen et al., 2023).
2.3.Environmental data
For urban data, we used the ground analytical dataset of the Czech Statistical Office (CZSO, 2009–2019).This dataset contains information about urban development and its surroundings, including information on the human population, the coefficient of ecological stability and the land use around the city, such as the built-up area.This database was used to get information about the total area of the individual cadastres(LAU 2).The Copernicus database, which includes environmental information, was used for the environmental data.The Copernicus Programme is the European Union’s Earth observation programme and makes it possible to monitor our planet and its environment.It provides information services based on Earth observation satellites and in situ data.The raster included forest-type data at a 100-m resolution (datasets from 2012, 2015 and 2018) (European Environment Agency, 2018a,2018b, 2020a), grassland data a 100-m resolution (datasets from 2015 to 2018) (European Environment Agency, 2018c, 2020b), water and wet area data at a 100-m resolution (datasets from 2015 to 2018) (European Environment Agency, 2020c, 2020d), small woody features data at a 5-m resolution (dataset from 2015) (European Environment Agency,2019c) and CORINE land cover data at a 100-m resolution (datasets from 2012 to 2018) (European Environment Agency, 2019a, 2019b).The next raster dataset came from the EarthEnv server and contained information on homogeneity and heterogeneity.Both datasets had a 1-km resolution (Tuanmu and Jetz, 2015).A large amount of CORINE land cover classes caused a large imbalance in their representation in the studied landscape.Therefore, some of the CORINE land cover classes were simplified by merging with other similar classes to improve predictive power of statistical models (Appendix Table S1).
2.4.Data processing
We mainly used Microsoft Excel (Microsoft Corporation, 2022) and a geographic information system (GIS) for data processing (ESRI, 2021).In Excel, we paired the CZSO ground analytical dataset and the bird species data from the CSO.For pairing, we used the unique nomenclature of municipalities codes (CISOB).Every city in Czech Republic has a unique code of this type, which helped us to pair the data correctly.When possible, we paired the data with the smallest time differences between the datasets.For the evaluation of raster datasets, we used the ArcGIS Pro system.As mentioned above, the CSO bird dataset contains GPS coordinates, which we used in the GIS to identify data from the raster.To evaluate how landscape factors (Copernicus data) affect birds on the local and regional scales, we used three buffers for analysis with a 100-, 500- and 1000-m radius around each data point, respectively.We used these specific scales because the similar buffers around studied points were selected in other studies (for example: Morelli et al., 2013;Litteral and Shochat, 2017; Humphrey et al., 2023).For example, the study of Litteral and Shochat (2017) describe, that most relevant scale for studying the trend in bird diversity at the landscape scale is around 500–2500 m.We used three different scales to determine which is the most relevant for explaining the distribution of bird diversity, calculated as the sum of species for a given cadastre.It means, that all species records for the same cadastre were pooled together and then the number of species was calculated for it.The buffer of 1000 m radius was evaluated by the model as the best representation of the landscape structure with regard to the spatial diversity distribution.In the subsequent statistical analyses, we used those proportions of individual land covers,which were calculated on the basis of the most appropriate scale.To calculate the values at the above radius scales, we used the ‘tabulate area’ function.The results of this function were a percentage representation and an area of examined landscape factors on three scales.To calculate the homogeneity and heterogeneity, we used a function to extract values from specific points rather than areas.This type of function evaluates the values at specific points using, for example, a buffer or a polygon around each point.
2.5.Statistical analysis
The relationship between the explanatory variables and species richness was studied using a generalized linear model (GLM) with a negative binomial error distribution and a log link function.To compensate for the confounding effect of spatial autocorrelation, the decimal logarithm of the district area and the GPS coordinates of the species observation records were used as covariates.To balance the risk of overfitting and at the same time failure to capture important aspect of bird diversity distribution we used ‘dredge’ function from the MuMIn package (Barto´n, 2022).This function returns a list of models with every possible combination of predictor variables and selects the best model by comparing the Akaike information criterion for a small sample size(AICc).To incorporate estimates from multiple candidate models which have similar support (ΔAICc ≤2) in the data, we have summarized the information by model averaging procedure by applying AICc weights.Parameter estimation was performed based on the ‘full average’ method(i.e., averages were obtained from the regression coefficients of all the models considered, and the regression coefficients for variables that were not included in the resulting models were assigned a value of zero).In addition, we were also aware of the negative effect of multicollinearity in the process of averaging of regression coefficient (Cade,2015).Therefore, final model was also checked for the multicollinearity by calculating variance inflation factor for each explanatory variables by using “vif” function (Fox and Weisberg, 2019).Models did not include interactions between factors and nonlinear predictors.In the Appendix Table S1, you can see a list of all the explanatory variables entering the model.The explanatory variables in the final model were tested using the chi-squared test, and the Wald test was used for testing the regression coefficients.The data were analysed in the R statistical programme(R Core Team, 2022).
Unfortunately, we did not have a complete phylogram available for the analysed bird taxa.Therefore, a phylogenetic tree based on a hierarchical taxonomy was proposed (Šťastný et al., 2021; Dungel et al.,2021).The phylogenetic categories used in the analysis were order,family, genus and species.We constructed a rooted ultrametric phylogenetic tree, and branch lengths were calculated using the Grafen method (Grafen, 1989).Phylogenetic diversity was calculated from the sum of the total branch length that span the members of the studied species (Faith’s index) (Faith, 1992).Faith’s phylogenetic diversity measures the amount of the phylogenetic tree covered by the studied community.Simply it is phylogenetic generalization of species richness.Therefore, it is not surprising that species richness and Faith’s phylogenetic diversity is highly correlated.Next, we calculated the standardised effect size based on Faith’s phylogenetic diversity index using the ‘ses.pd’ function to remove the influence of richness.This function compares observed phylogenetic diversity to the values expected under various randomisations.The randomisation was performed using the‘independentswap’ null model (Gotelli, 2000) while maintaining species occurrence frequency and sample species richness.We created a GLM model with a Gaussian error distribution to fit the relationship between the explanatory variables and phylogenetic diversity.To select the best combination of explanatory variables explaining phylogenetic diversity variability, we followed the same procedure as in the species richness analysis.The phylogenetic analyses were performed using the picante package in the R program (Kembel et al., 2010).
To examine the spatial trends in the distribution of bird diversity, we interpolated diversity for each scale by applying the kriging method using the ‘krige’ function in the R program (Pebesma, 2004).The kriging technique estimates diversity values at unsampled locations based on the information obtained from the structure of the spatial model depicted by a spatial covariance function (i.e., a semivariogram).At first, we described spatial autocorrelation by using the variogram function from the gstat package in R (Pebesma, 2004).Using variogram,we were able to identify a mathematical function representing the semi-variance describing the spatial autocorrelation calculated between each possible pair of sampling localities.The mean semi-variance was then plotted against an increasing distance interval to obtain the semivariogram.From the fitted model, we obtained information on the total variance (i.e., sill variance) and the spatial variation at distances shorter than the minimum sample spacing (i.e., nugget variance).This spatial function defined the size and shape of the area for the diversity interpolation.We used exponential spatial covariance structure as the most appropriate mathematical description of the spatial correlation.This method specifies that the spatial correlation decreases as an exponential function of the distances between neighbourhood points.
To visualise the association between each land cover type and species composition, a partial canonical correspondence analysis (pCCA) was applied.The abundance of each bird species was transformed by a decimal logarithm.We used the decimal logarithm of the district area and the GPS coordinates of the species observation record as a covariable to eliminate the effect of spatial autocorrelation.The significance of the ordination axes was tested by the unrestricted permutation of the residuals of the reduced model.The testing and selection of the most parsimonious combination of land cover types explaining most of the variability in the species data was performed using a forward selection procedure.Ordination analyses were performed using CANOCO 5 (ter Braak and Šmilauer, 2012).
To model the relationships between bird traits (i.e., trophic and migratory status and average body size of birds, for more information see subsection 2.2.Species distribution data) and the proportion of land cover type, we used a combined RLQ and fourth-corner analysis with the‘ade4’ package in R (Thioulouse et al., 2018).To solve the issue of inflated type I error, we used a modified version of the fourth-corner analysis, adjusting the significance by taking the maximump-value of the row- and column-based permutation test (ter Braak et al., 2012; ter Braak, 2017).The RLQ method allowed us to analyse a covariance matrix between bird traits and land cover types mediated by species abundance (Dolédec et al., 1996).The fourth-corner method was used to test multiple associations between bird traits and local and landscape variables.We created a factorial map of the RLQ analysis and an RLQ biplot to visualise the significant associations identified by the fourth-corner analysis.
3.Results
A total of 223 bird species from a total of 20 orders were evaluated.Based on the analysis of the representation of species traits the body size of the birds ranged from 9 to 153 cm.The majority of birds had a body size ranging from 9 to 23 cm.Aside from the abundance in this range,the numbers were quite evenly distributed, and the average body size was 33.21 cm.Regarding migration, we evaluated a total of 139 migratory species, 40 partial migratory species and 44 permanent species.We counted five trophic guilds: herbivores, insectivores, carnivores, omnivores and seed eaters.The omnivorous and insectivorous birds were the most numerous, with 75 omnivorous species and 71 insectivorous species.The number of carnivorous birds was 51, that of herbivores was 15, and that of seed eaters was 11.
3.1.Association between land cover composition and bird diversity
Based on our results forest land cover classes (i.e., mixed, coniferous and broadleaf forests) and additional woody features (i.e., patchy tree structures which usually link the linear landscape features to forests)had the highest negative effects on species richness (Fig.2).The proportion of urban area in the landscape was also found to have a negative influence on species richness.We also revealed that presence of permanent water bodies, small woody features and shrub cover has the most positive impact on bird species richness (Fig.2; Table 1).These findings correspond to the spatial prediction of model averaging results using all models which have similar support (ΔAICc ≤2) in the data(Appendix Table S2), which predicts three diversity hotspots situated into: a) the northeastern edge of Brno (where are situated rocky slopes with steppe grasslands and bushy vegetation); b) habitats between Mikulov and Breclav city (where fine mosaic of wetlands, meadows,woods and dry sands can be found); and c) the area between Mikulov,Hustopece and Breclav (This area is characterized by the largest water bodies in South Moravia) (Fig.3A).The same pattern of richness distribution is visualized also by using raw measured data (Appendix Fig.S1.1).The result of GLM model showed that small woody features had positive impact on species richness until their proportion did not exceed ~5% of the landscape composition over a 300 ha area, then decreasing slightly, reflecting a quadratic relationship (Appendix Fig.S2).
The phylogenetic diversity was negatively correlated with the percentage of forest cover, grassland and urban areas (Table 2).Cultivated land with a high proportion of natural vegetation significantly supported phylogenetic diversity.In addition, phylogenetic diversity was found to be positively influenced by the presence of pastures, in contrast to meadow (grassland) ecosystems (Fig.4; Table 2).In more detail, the spatial distribution of predicted phylogenetic diversity shows that most of the landscape cover has tendency to generate phylogenetically clustered bird assemblages (SESFaith’sPDvalues approaching -2) except for the area in the northwest between Blansko and Tisnov and the area laying between Brno and Znojmo (Fig.3B).Both localities are characterized by a cultural landscape in which larger forest areas are often represented, and agrarian areas contain a large number of linear features(e.g., avenues, forest strips or copses).The same pattern of diversity distribution is visualized also by using raw measured data (Appendix Fig.S1.2).Comparing the explained variability between spatial and landscape characteristics, we found that spatial distribution of phylogenetic diversity was less affected by spatial autocorrelation and area size than species richness distribution (Table 3).
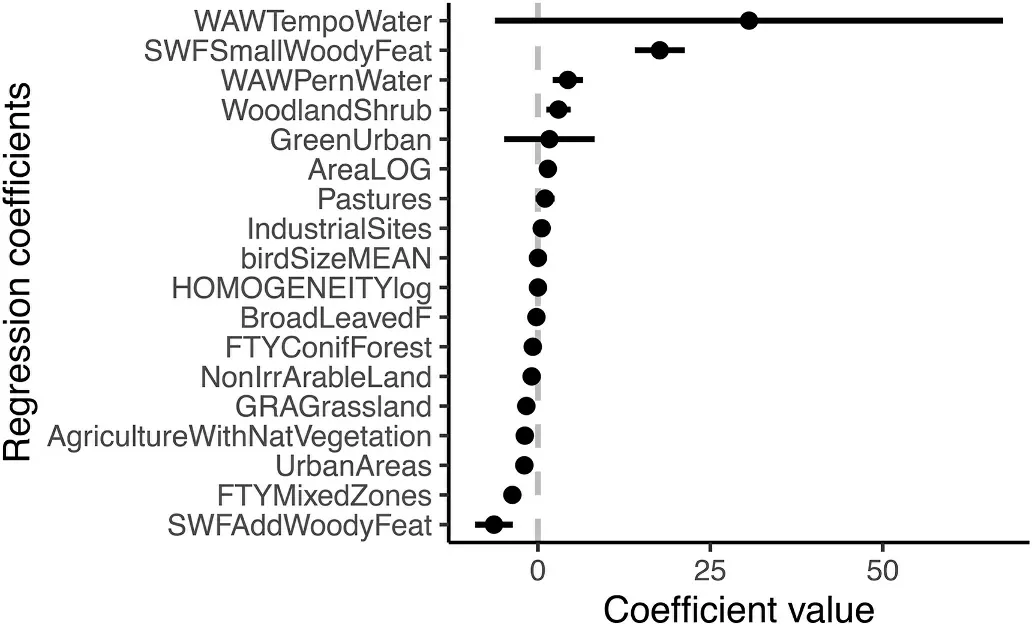
Fig.2.Model averaging results showing which land cover type or landscape characteristic positively or negatively correlated with species richness distribution within the buffer radius 1000 m.The dots represent the coefficients from the regression analysis (i.e., the GLM), and the lines represent 95% confidence intervals.The abbreviations of used variables are individually described in Appendix Table S1.
3.2.Association between land cover bird species composition
By using pCCA analysis, we identified that proportion of arable land with a minimum forest cover, the presence of permanent water bodies and proportion of forest habitats had the greatest impact on the bird species composition (Appendix Table S3).This analysis also identified that the highest gradient within the species abundance distribution was caused by a gradient formed by forest ecosystems on one side and agricultural ecosystems on the other (Appendix Fig.S3).The second greatest gradient divided the birds according to whether they prefer water bodies and wetlands or are more likely to be found in non-forested areas (Appendix Fig.S3).Moreover, based on the pDCCA biplot, the most indicative species of: a) water areas was Mallard Duck (Anas platyrhynchos) and Common House Martin (Delichon urbica); b) pastures was Yellowhammer (Emberiza citrinella) and Song Thrush (Turdus philomelos); c) urban areas was Barn Swallow (Hirundo rustica) and the Eurasian Tree Sparrow (Passer montanus); d) forest habitats was Great Woodpecker (Dendrocopos major), Common Finch (Fringilla coelebs) and Tits (Parussp.); and e) cultivated agricultural land was Eurasian Skylark(Alauda arvensis), Buzzard (Buteo buteo), Marsh Harrier (Circus aeruginosus) and Common Kestrel (Falco tinnunculus).Ordination diagram also shows a tendency of agricultural landscape together with small woody features and urban areas to increase species composition heterogeneity of bird communities in rural landscape (Appendix Fig.S3).
3.3.Distribution of ecological bird guilds among land cover classes
We also studied the association between land cover type and the distribution of ecological bird guilds using three ecological traits: trophic guild, migration and body size.Our results revealed that the body size and migratory ability of birds were significantly positively correlated with the presence of water in the landscape and that body size was additionally correlated with the presence of small woody features (Appendix Fig.S4).The presence of migratory and large birds was also negatively correlated with the occurrence of deciduous and mixed forests (Appendix Fig.S4).The trophic guild of birds was significantly affected by the presence of forests and wetlands, with forest habitats negatively correlating with the occurrence of carnivores and the presence of wetlands negatively correlating with the occurrence of omnivores (Appendix Fig.S4).In general, the distribution of the studied characteristics of birds was mainly influenced by two landscape gradients: forest–non-forest and water bodies–dry habitats (Appendix Fig.S4).
4.Discussion
Based on the data collected from the public ornithological database of the Czech Republic, we found a clear trend in the bird species richness and phylogenetic diversity across the landscape.It was determined that the proportion of permanent water and small woody features (e.g.,woodlots) had a significant positive effect on the species richness, while mixed forest, urban areas and additional woody features had a negative effect.Phylogenetic diversity was negatively associated with the proportion of woodlands and urban areas and positively associated with pastures and agriculture area with scattered natural vegetation.These results suggest that a complex agricultural landscape with a high proportion of wetland and scattered vegetation (e.g., woodlots, forest strips or copses) is important for maintaining a high bird richness and diversity of ecological guilds.On the other hand, we also found out potential danger of structurally rich rural landscape in the increase in the density of forest features or overgrowth by forest vegetation.
We revealed that woody elements, such as shrubs and linear or scattered vegetation, are important to support the species richness within the agricultural landscape.The bird population can benefit from these as places for nesting, foraging and shelter (Jakobsson and Lindborg, 2017; Sasaki et al., 2020).Even a small amount of these landscapeelements can have a great impact on structural habitat complexity(Manning et al., 2006; Fischer et al., 2010).The effect size of small woody features on species richness will depend on the nature of a farmland which is related to an extent of pastoral versus crop lands, the varieties of crops grown, the configuration of different crop and natural land covers, and the intensity of techniques used to grow crops(Tscharntke et al., 2002; Manning et al., 2006; Fahrig et al., 2011;Wilson et al., 2017).Therefore, we expect the largest impact of small forest patches in simple and structurally homogeneous landscapes with a low proportion of non-crop habitats (Tscharntke et al., 2012).In addition to the woodlots, traditionally managed orchards can also serve as a tool for maintaining diversity within crop production landscapes.Traditional high-stem orchards with sparse canopy unlike intensively-managed, low-stem fruit-tree plantations, contain mature trees with hollow trunks and branches which offer nesting sites and the associated grasslands, if extensively managed, provide invertebrate prey for birds (Horak et al., 2013; Rime et al., 2020).Bennett et al.(2022)pointed out that sparse, scattered trees in farmland have a critical complementary role in rural landscapes.Sasaki et al.(2020) also highlighted that scattered greenery and woodland edges provide important habitats for farmland birds especially if they are rich in shrubs and deciduous trees.On the other hand, this does not say anything about quality of the bird community, because in such landscape features usually occurs generalist species with high dispersal ranges (Dvoṙáková et al., 2022).Le Roux et al.(2018) highlighted that younger, smaller tree alone is not sufficient to support all bird species.Our finding may just be an artefact of the spill-over effect that usually manifests itself in a landscape with a simple structure.High productivity of crop fields can make crop fields a source of organism spilling to adjacent natural habitats (Tscharntke et al., 2012).
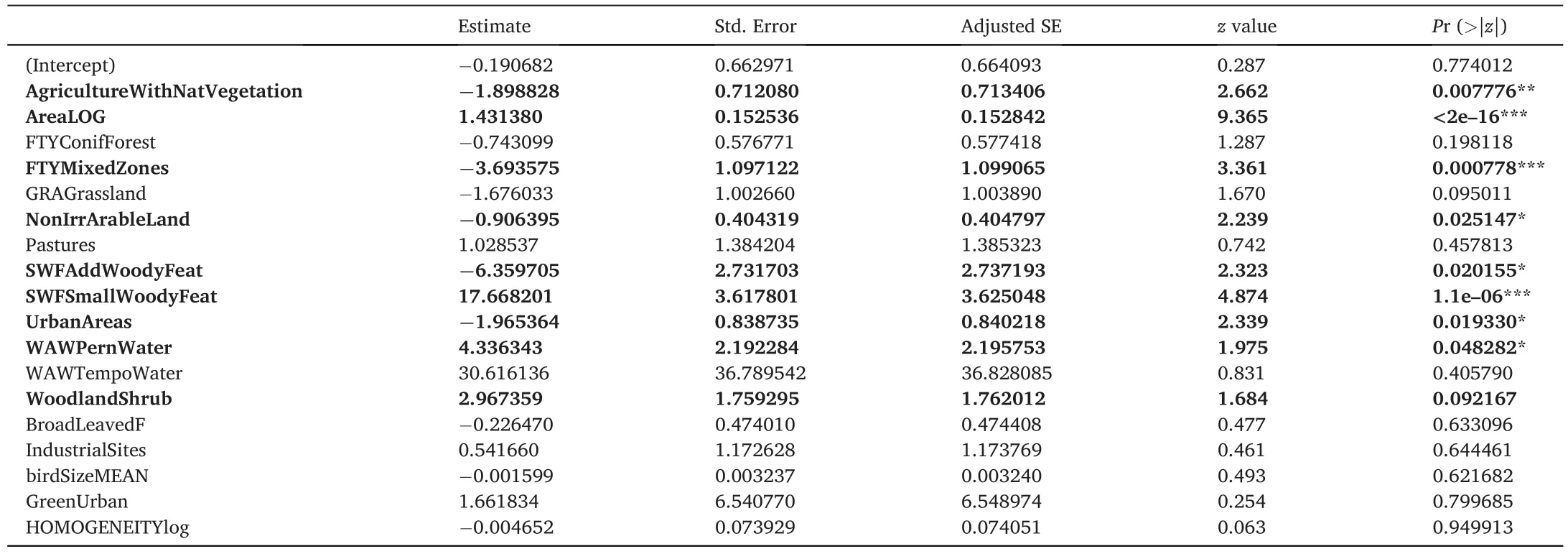
Table 1Model averaging results selecting the best parsimonious combination of predictors explaining species richness variability.

Fig.3.Visualisation of the model averaging results: (A) species richness and (B) phylogenetic diversity (i.e., standardized effect sizes of Faith’s phylogenetic diversity index).We used the ordinary kriging interpolation technique with an exponential spatial covariance structure to estimate the diversity distribution for the entire region of South Moravia.The predicted values are marked in a grey scale, the darker the shade, the more species-rich the defined geographical area is.In the case of phylogenetic diversity, the lightest shades mean the tendency of the assemblages to consist of the related taxa, on the other hand, the darkest shades indicate tendencies of the assemblages to consist of phylogenetically unrelated species.A map scale is approximately 1:1,100,000.

Table 2Model averaging results selecting the best parsimonious combination of predictors explaining the variability in phylogenetic diversity (i.e., standardised effect sizes of Faith’s phylogenetic diversity index).
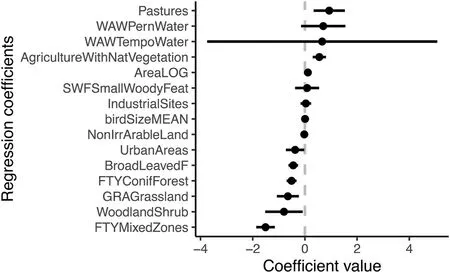
Fig.4.Model averaging results showing which land cover type or landscape characteristic positively or negatively correlated with phylogenetic diversity distribution (i.e., standardized effect sizes of Faith’s phylogenetic diversity index) within the buffer radius 1000 m.The dots represent the coefficients from the regression analysis (i.e., the GLM), and the lines represent 95% confidence intervals.The abbreviations of used variables are individually described in Appendix Table S1.

Table 3Comparison of explained variability (null deviance–residual deviance/null deviance) of species richness and phylogenetic diversity by GPS coordinates(space), district area (area) and best combination of land cover types (landscape)calculated using a generalized linear model (GLM) with a negative binomial error distribution along with a link function log in the case of species richness and a Gaussian error distribution in the case of phylogenetic diversity (i.e.,standardised effect sizes of Faith’s phylogenetic diversity index) at 1000-m scale.
As the second most important landscape element generating substantial diversity was wetlands and water bodies.Water in the agricultural landscape can serve as avian diversity ‘hotspots’.This is consistent with the wide recognition of freshwater ecosystems as the most diverse ecosystems for plants and animals (Reid et al., 2019; Dertien et al.,2020).The presence of water in the landscape has a positive influence on bird richness and diversity (Tu et al., 2020).In a study by Zamora-Marín et al.(2022), it was found that small human-made water bodies can have a positive influence on farmland birds if their structure is properly managed.In our study, we also found that having water habitat in the landscape has a positive influence on migratory birds (Appendix Fig.S3).The waterbodies and watercourses in the landscape often serve as stops for migratory birds and provide them a place for foraging(Pocewicz et al., 2013).
We revealed, that the proportion of arable land exhibited a negative association with bird species richness.Agricultural land in the Czech Republic is mostly formed by large homogeneous fields, with one crop type having a negative impact on many organisms in the landscape(Lojka et al., 2021).For example, Šálek et al.(2021) found that the Austrian agriculture management system of small-scale farming promotes significantly higher species richness of birds than that practiced in the Czech Republic border region, where large-scale agriculture management dominates.The negative effect of arable land may indicate a homogeneous landscape structure on the regional scale of South Moravia.Surprisingly, we also found the same negative association for agricultural areas with a significant proportion of natural vegetation.We explain it by the fact that the scattered vegetation within the agricultural landscape still insufficiently compensates for the negative effects of fields with agricultural crops.One of the possible reasons may be predominance of young structurally simple forest stands in the agricultural landscape of South Moravia.In addition, our results agree with the work of Bennett et al.(2022) that an increase in the density of forest features within landscape beyond a certain limit can have a negative effect on bird diversity (Appendix Fig.S2).In the context of this result, we interpret the negative association between agricultural areas with natural vegetation and the number of species as a possible consequence of the woodlots distribution, which may have a negative effect on bird diversity at higher densities.Inadequate species and spatial structure of woodlots can cause birds that breed in small forest patches to have small local populations making them more prone to extinction (Andrén,1996).Le Roux et al.(2018) revealed that habitat resources associated with large trees, such as hollows are important for bird diversity.Moreover, within landscapes that have only small forest remnants, the regional pool of forest-breeding bird declines and migrants become less likely to encounter fragments (Brown et al., 2019).On the other hand,this type of land cover was positively associated with diversity of higher taxons.Possible explanation is diverse land cover structure within agroecosystems on large scales compared for example to uniform forest stands.Our results may indicate that this type of landscape has the potential to host few bird species but with diverse ecological functions.It is in the concordance with the work which pointed out that species composition diverged in the initially quite similar forest fragments based on the different landscape context (Laurance et al., 2007).Another type of open habitat with a tendency to negatively affect the number of species were meadows.This may be connected with the expansion of grassland area caused by the abandonment of unproductive areas after communism fell in 1989 in the Czech Republic and the relatively low intensity of grassland management, e.g., low mowing frequency and low stock densities on pastures (ˇCerná et al., 2007; Reif and Hanzelka, 2016).Based on our results, we can see a tendency of pastures positively influence species richness compared to meadows.This may reflect differences in the management of pastures and meadows (Bucher et al., 2016), which is in concordance with a study by Kamp et al.(2018), who revealed that species of farmland birds mostly profit from the abandonment of croplands and grassland management,unlike grassland bird species.Other causes of the decline in the diversity of grassland birds is the intensification of production in agroecosystems(Hanioka et al., 2018), which leads to the loss and degradation of habitats through the overgrazing or intensive mowing of grassland areas(Hill et al., 2014; Reif and Vermouzek, 2019).
We did not find positive association between forest cover and bird diversity.These results contradict the works, which state that forest ecosystems host an important part of bird diversity (Reif et al., 2022).Our result may indicate either that forests are not the source of bird diversity in agricultural landscapes or that their poor condition (i.e.,structural and age homogeneity) reduces bird diversity.The cultivation of conifer monocultures of the same age was introduced in Central Europe from the middle of the 18th century and continued on until recent times (O’Hara, 2016).In addition, after WWII, coppice woods diverse in both structure and species were abandoned in many parts of Europe and replaced by homogeneous high forest (Müllerová et al.,2014).The legacy of changes in these forest management practices can synergistically impoverish bird communities despite the fact that total forest area in the Czech Republic is increasing.This is in the agreement with a study by Betts et al.(2022), who revealed that forest degradation through the reduction of old forest types and intensifying forestry may be the primary driver of forest biodiversity loss.
Based on the previous results, we suggest that the actual negative association of landscape homogenisation with diversity distribution is connected with the increasing of forest area based on the overgrowing of abandoned pastures and meadows.This statement is in concordance to the work of Romero-Calcerrada and Perry (2004), who found that land abandonment led to the recovery of semi-natural vegetation and the long-term recovery of woody area cover.The consequences of this succession negatively affect the species that tend to favour open landscape areas (Sirami et al., 2007).Although the abandonment of agricultural land may increase heterogeneity in the short term, at the same time, it leads to forest overgrowth in the long term, which will again lead to landscape homogenisation (Gellrich et al., 2007).An example of this is provided by the Red-backed Shrike (Lanius collurio), which prefers open habitats with scattered forest and shrub vegetation.The abundance of this species has decreased as a result of land abandonment, leading to gradual forest regrowth (Brambilla et al., 2010).
We are also aware that the association of specific land cover type with bird diversity will be influenced by the spatial scale, for example some bird guilds in grassland environments are more associated with local conditions, while water birds with good fly ability perceive the landscape at larger scales (Morelli et al., 2013).A study by Galitsky and Lawler (2015) found that landscape scale dependence may be driven by bird species characteristics.In their study, landscape scale factors were determined to be five times more important for Corvidae omnivores than for seed eaters and frugivores and 10 times more important for flycatchers.A study by Callaghan et al.(2018) showed that bird diversity distribution was related less to the habitat predictors at the local and more to those at the regional scale.Another important determining factor influencing how bird species react to the landscape is their degree of habitat specialisation (Zurita et al., 2017), the sensitivity to habitat grain is also species-specific (Betts et al., 2014).
Urbanisation is one of the largest threats to wildlife and biodiversity(McKinney, 2002; Isaksson, 2018).In our study, we found that it caused a strong negative effect on bird species richness, at the same time, its influence was lower than that of agricultural land.Biodiversity has a declining trend in urban environments (Chamberlain et al., 2018) and it seems that the homogenisation process of urban areas may result in a decrease of specialist bird species with gradual replacement by generalist species in the city areas (Clergeau et al., 2006; Xu et al., 2022).Interestingly, when comparing industrial areas (e.g., areas mainly occupied by industrial activities of manufacturing, road and rail networks) and urbanized areas (e.g., areas mainly occupied by dwellings and buildings used by administrative or public utilities), we found a difference between the effect of these two land cover types (Fig.2).In particular, the industrial sites had a weak positive influence on species richness, in contrast with the strong negative impact of urban areas(Fig.2).In some cases, industrial areas can also have a positive effect on birds, such as around quarries (Salgueiro et al., 2020) or railways and power lines.For example, the pylons at the bases of power lines can serve as cover from predators or nest sites (Morelli et al., 2014), and mining areas can mimic natural nesting habitats (Salgueiro et al., 2020).In a study by Pellissier et al.(2012), it was found that, in urbanized landscapes, omnivorous, arboreal and ground species responded to building height heterogeneity, while insectivorous species were sensitive to the spatial arrangement of green areas.The height of buildings can also influence the flight paths of birds (Yang et al., 2022).Urban greenery can be crucial for maintaining biodiversity in cities, but we did not find a significant association between bird diversity and this land cover type in our study.Contrary to our results, other studies have confirmed the positive effect of greenery on the bird diversity of urban areas (Carbó-Ramírez and Zuria, 2011; Threlfall et al., 2017; Leveau et al., 2019; Morelli et al., 2021).In particular, the size of green space area within towns can lead to an increase in the bird species diversity within urbanized ecosystems (Callaghan et al., 2018).For example,Carbó-Ramírez and Zuria (2011) found that a high level of disturbance decreased bird diversity along road strip vegetation corridors in comparison to parks and gardens.
5.Conclusion
In conclusion, this study found that bird species richness and phylogenetic diversity was dominantly associated with the proportion of linear or scattered vegetation within agricultural landscape, wetlands or water bodies.Interestingly, our results showed that agricultural land with natural vegetation is not significantly associated with bird species richness, meaning scattered natural vegetation in agroecosystems does not sufficiently mitigate the negative influence of a homogeneous landscape mosaic on bird communities.This finding may correspond to inadequate species and spatial structure of the majority of woodlots which may at high densities negatively influence bird diversity at the landscape scale.The percentage of woody vegetation was also found to be associated with low bird species richness.We thus suggest that the expansion of forests (with simple species composition and vertical structure) at the expense of abandoned agricultural areas and their overgrowth with trees could be one critical driver of the bird species decline in the agricultural landscape.We also revealed that land cover types increasing species richness do not completely coincide with land cover type increasing phylogenetic diversity.We think that diverse mosaic of land covers in agricultural landscape can generate diverse guild composition.To restore and/or maintain the bird species richness and phylogenetic diversity in the agricultural landscape, we suggest increasing the number of wetlands and small interactive elements (e.g.,hedgerows, headlands, woodlots and copses) while preventing the overgrowth of pastures by structurally simple forest.Moreover, when increasing the percentage of interaction elements (i.e., woodlots), their possible negative impact on the diversity of communities should be taken into account.
Funding
This study was supported by the internal grant agency of the Faculty of AgriSciences of Mendel University in Brno (AF-IGA2022-IP-034).
Ethics statement
Ethics approval was not required for this study.We did not manipulate any living animals.Data were obtained from a public database.
CRediT authorship contribution statement
Denisa Dvoˇr´akov´a:Conceptualization, Data curation, Investigation, Methodology, Project administration, Resources, Writing – original draft, Writing – review & editing.JanˇSipoˇs:Conceptualization, Data curation, Formal analysis, Methodology, Software, Writing – review &editing.Josef Suchomel:Conceptualization, Investigation, Methodology, Writing – review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We would like to thank the Czech Society for Ornithology for operating the ornithological database and all the volunteers who contributed data to the public database, without whom the study could not have been performed.We would also like to thank Kontrolujeme s.r.o.for the professional language editing.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.avrs.2023.100147.
杂志排行

Avian Research的其它文章
- Selecting the best: Interspecific and age-related diet differences among sympatric steppe passerines
- Morphology and morphometry of two hybridizing buntings at their hybrid zone in northern Iran reveal intermediate and transgressive morphotypes
- Quiet in the nest: The nest environment attenuates song in a grassland songbird
- Characteristics of cross transmission of gut fungal pathogens between wintering Hooded Cranes and sympatric Domestic Geese
- Fecal DNA metabarcoding reveals the dietary composition of wintering Red-crowned Cranes (Grus japonensis)
- Short-term night lighting disrupts lipid and glucose metabolism in Zebra Finches: Implication for urban stopover birds