黔中地区马尾松-甜槠针阔混交林群落结构特征与林木空间分布格局及其关联性
2024-01-20文春玉唐雪娅
金 鹏,徐 明,文春玉,聂 坤,杨 雪,唐雪娅,魏 珊,张 健
(贵州大学 生命科学学院,贵州 贵阳 550025)
空间格局作为森林群落结构的一个重要属性[1],能反映森林群落中植物的扩散模式、物种间的相互作用、微环境以及其他随机事件[2]对生态系统结构和功能的影响[3]。空间格局可描述生态过程与空间模式的关系,揭示森林群落结构和动态变化的重要性[4]。森林不同生态过程如种子萌发、物种竞争和死亡可能会在个体空间结构上留下特定的痕迹,通过空间格局分析可以揭示这些规律[5-6]。对森林群落空间格局的研究[7],涉及热带[8-9]、温带[10]、地中海[11]等地区森林群落并被广泛用于森林群落的管理和优化[12-13]。
马尾松(Pinusmassoniana)是我国南方地区分布广泛的乡土先锋树种,具有耐贫瘠、适应性强、生长快和易成林等特点[14],常作为荒山造林、喀斯特恢复的主要树种,在我国生态环境建设中有着重要地位。因马尾松林纯林面积大[15],经长期多代连栽,其低质低效林面积大,林分质量普遍较差、生产力较低、抵御干扰能力弱和土壤退化等问题突出[16],通过实施近自然经营或人工引种的方式诱导其转化为结构复杂、生产力高、树种多样的异龄混交林,是低效马尾松纯林提质增效的重要途径[17]。目前,关于马尾松林的研究较多,主要集中在马尾松林群落结构[18]、土壤理化[19]和碳储量[20]等方面,对马尾松天然次生针阔混交林的研究不足[21],其中对马尾松林空间格局及其种内相互作用的研究相对较少[22]。因此,本研究以马尾松纯林和马尾松-甜槠(Castanopsiseyrei)混交林为对象,分析两者的空间分布格局和种间关联性,探讨2种林分结构差异、生境异质性对两者空间分布格局的影响,以及不同径级间树种的空间分布格局和种间关联性,以期为马尾松纯林阔叶化改造过程中合理树种空间配置培育异龄混交林提供科学参考。
1 材料与方法
1.1 研究区概况
研究区位于黔中腹地开阳县(106°45′-107°17′E,26°48′-27°22′N),海拔1 000~1 400 m,属典型的山原地貌,北亚热带季风湿润气候,年平均气温10.6~15.30 ℃,年无霜期为315 d左右。森林覆盖率达52.92%,主要针叶树种为马尾松和杉木(Cunninghamialanceolata),主要阔叶树多为壳斗科(Fagaceae)、山茶科(Theaceae)、冬青科(Aquifoliaceae)、樟科(Lauraceae),森林土壤主要以石灰土与黄壤为主。
1.2 样地设置与群落调查
在开阳县禾丰乡分别选取天然次生林马尾松-甜槠针阔混交林(p-cF)和近自然经营马尾松纯林(pF),每种植被类型分别设置1个50 m×50 m(2 500 m2)的植物群落调查样方,采用RTK测量仪(北斗海达TS5 Pro)将样方划分为100个5 m×5 m的单元网格,对样方内所有胸径(DBH)≥1 cm的树木进行每木检尺和GPS定位等信息,对草本和灌木进行群落调查,记录其高度、基径、盖度等指标;同时记录样地海拔、坡向、坡位和土壤类型等立地条件(表1)。
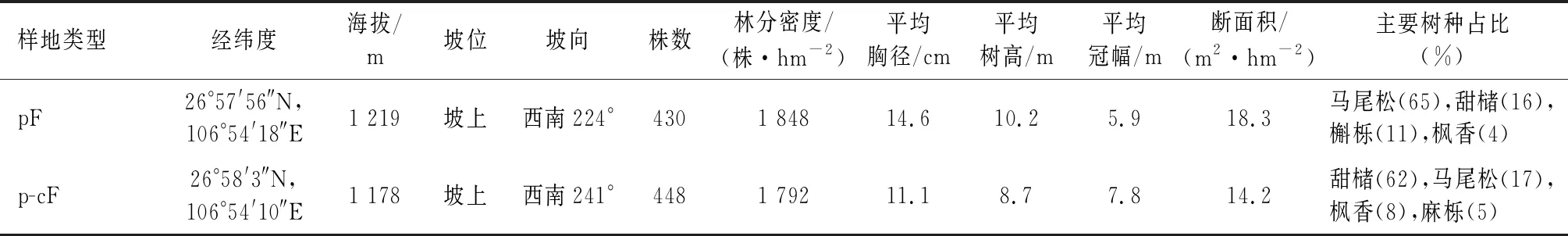
表1 样地基本情况
1.3 林分结构
径级是评价林分质量的主要依据,与林分的材积、盖度、胸高断面积等密切相关。本研究采用径级结构代替年龄结构的方法,根据样地内木本植物的调查情况,按照胸径结构将所测量DBH≥1 cm的林木划分为3级:DBH≤5 cm为幼树、5 cm
重要值是衡量及计算物种多样性的重要参数,是描述物种在群落中的地位和作用的综合数量指标[23],本研究筛选重要值前4位的树种进行研究。计算各物种的重要值,计算公式为:乔木重要值=(相对密度+相对频度+相对优势度)/3×100%,灌草重要值=(相对密度+相对频度+相对盖度)/3×100%。相对密度=(某种植物的个体数/全部植物的个体数);相对频度=(样方中某一树种的频度/所有树种的总频度);相对优势度=(样方中某一树种的个体胸高断面积和/所有树种个体胸高断面积总和);相对盖度=(样方中该种个体的分盖度/样方中全部个体所有分盖度之和)。
1.4 点格局分析
采用Ripley'sK函数演变而来的成对相关g(r)函数进行群落空间分布格局分析。g(r)函数是一个概率密度函数,是利用2点间的距离,计算任一点为圆心、半径为r、指定宽度的圆环内的点数量来进行空间点格局分析,以圆环代替圆,消除了大尺度上的累积效应,能更真实、直观地反映种群的空间格局[24]。对于单变量g11(r)函数,当实测值在包迹线内时,表示随机分布,当实测值在包迹线之上表示聚集分布,在包迹线之下表示均匀分布。对于双变量g12(r)函数,当实测值在包迹线内时,表示2个物种没有关联,实测值在包迹线之上代表2个物种为正关联,实测值在包迹线之下代表2个物种之间为负关联。
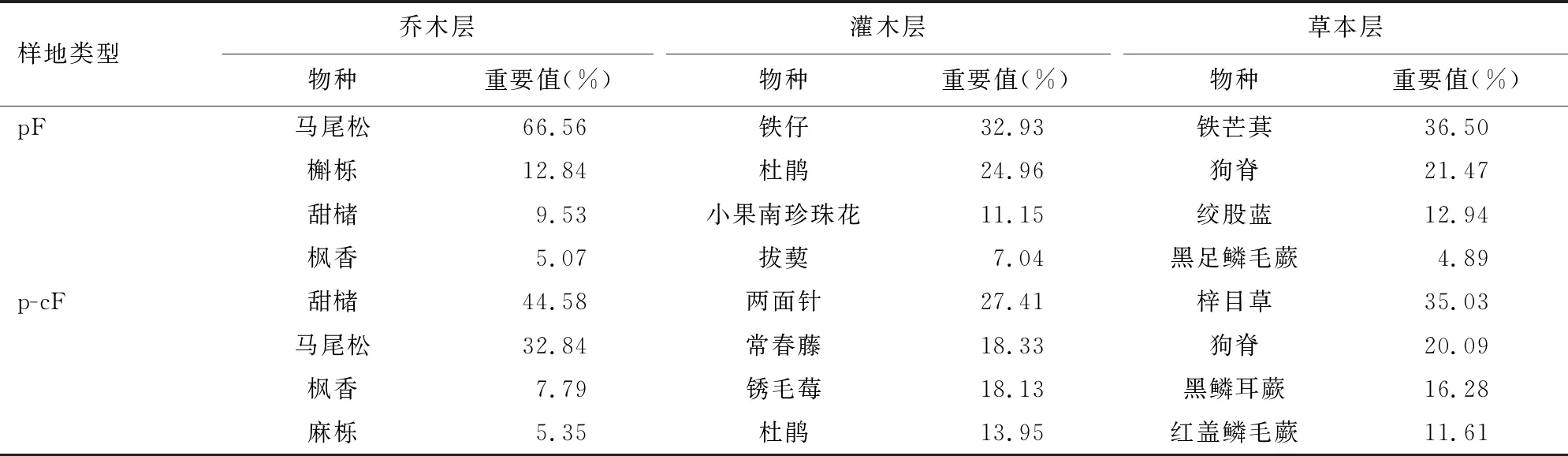
表2 不同样地类型不同冠层物种重要值
选择合适的零假设模型(null model),是进行空间点格局统计分析的关键[27]。本研究采用模型有:①完全空间随机模型(complete spatial randomness,CSR),也称同质泊松模型(homogeneous poisson process),其表示物种的空间点是完全互相独立的,不受任何生物或非生物过程影响,在研究区域内各点出现的概率相同,常作为零假设来检验当存在生境异质性时物种的空间分布[25];②异质性泊松模型(heterogeneous poisson process,HP),该模型是排除空间异质性效应的零假设模型,可检验排除大尺度环境异质性的影响,反映种群真实的空间分布特征[26]。本研究中,对于g11(r)函数选取了完全随机模型(CSR)来检验当存在生境异质性时物种的空间分布格局;采用异质性泊松模型(HP)来检验排除生境异质性时物种的分布情况;对于g12(r)函数用完全随机模型来分析2个物种之间的关联性。
根据最大距离不超过研究样地较短边长1/2的原则[27],将空间分析尺度设为0~25 m,并将其分别定义为小距离(0~10 m)、中距离(10~20 m)、大距离(20~25 m),栅格大小为1 m×1 m,圆环宽度为1 m,同时利用蒙特卡洛模拟检验199次,选取模拟值的第5最高值和第5最低值构建95%的置信区间,得到上下2条置信区间为95%的包迹线。
1.5 数据处理
采用Excel 2016整理数据,利用Programita 2018对主要树种进行空间点格局分析,采用Origin 2018软件进行绘图。
2 结果与分析
2.1 群落物种组成与结构特征
马尾松纯林与马尾松-甜槠针阔混交林群落组成和结构存在明显差异(图1)。调查共记录91种植物:马尾松-甜槠针阔混交林样地内共记录植物42种,其中乔木14种、灌木22种、草本6种,隶属于26科;马尾松纯林样地共记录植物68种,其中乔木17种、灌木21种、草本30种,隶属于35个科。
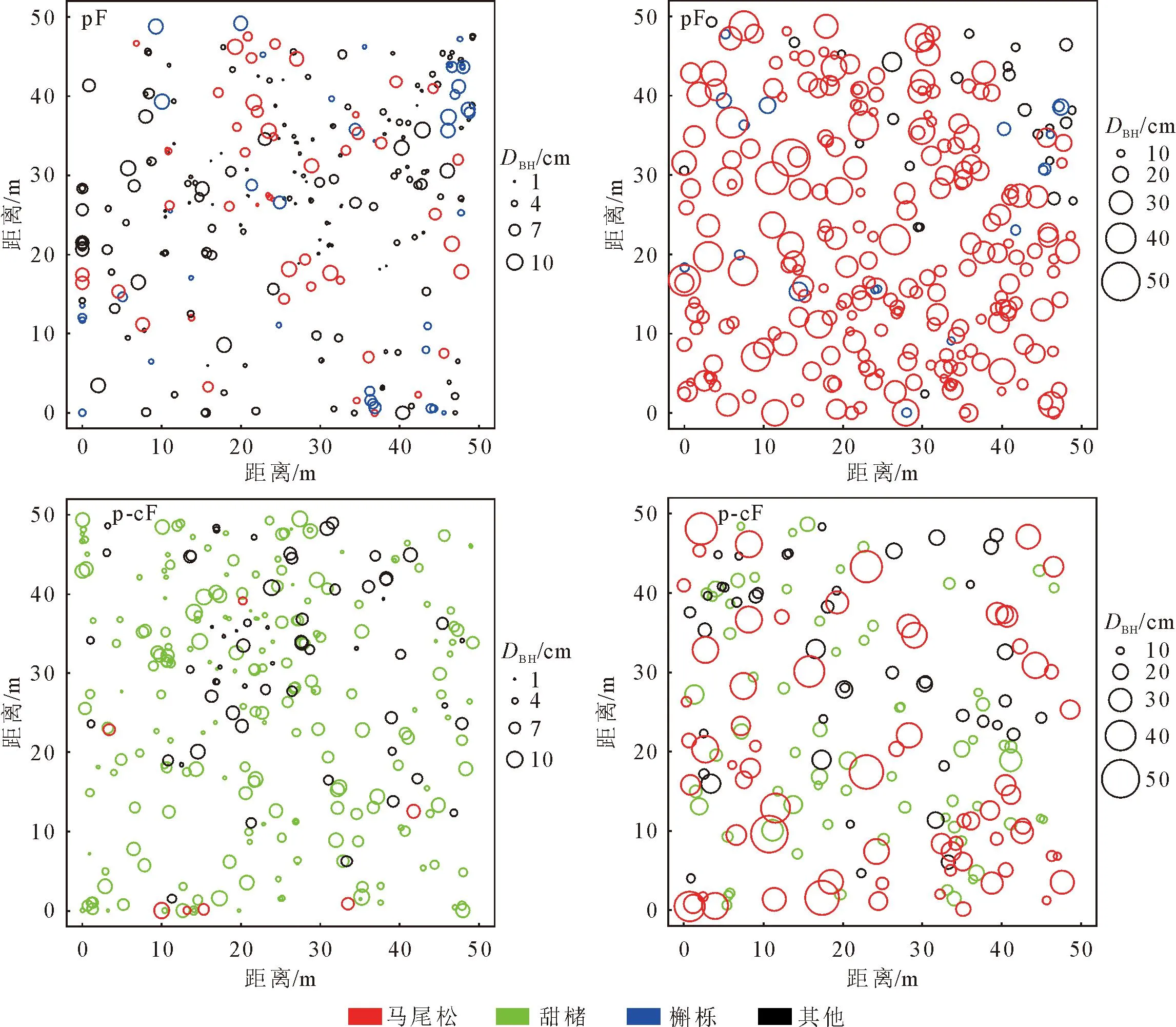
图1 不同样地类型主要树种径级分布
马尾松-甜槠针阔混交林与马尾松纯林植物群落重要值排序存在明显差异(表2)。乔木层上,马尾松-甜槠针阔混交林重要值前4位树种分别为:甜槠(44.58%)>马尾松(32.84%)>枫香(Liquidambarformosana,7.79%)>麻栎(Quercusacutissima,5.35%);马尾松纯林重要值前4位的树种分别为:马尾松(66.56%)>槲栎(Q.aliena,12.84%)>甜槠(9.53%)>枫香(5.07%);在灌木层上,马尾松-甜槠针阔混交林和马尾松纯林中重要值排序前4物种分别为:两面针(Zanthoxylumnitidum,27.41%)>常春藤(Hederasinensis,18.33%)>锈毛莓(Rubusreflexus,18.13%)>杜鹃(Rhododendronsimsii,13.95%)和铁仔(Myrsineafricana,32.93%)>杜鹃(24.96%)>小果珍珠花(Lyoniaovalifoliavar.elliptica,11.15%)>拔葜(Smilaxchina,7.04%);草本植物中,马尾松-甜槠针阔混交林和马尾松纯林中重要值排序前4物种分别为:梓木草(Lithospermumzollingeri,35.03%)>狗脊(Woodwardiajaponica,20.09%)>黑鳞耳蕨(Polystichummakinoi,16.28%)>红盖鳞毛蕨(Dryopteriserythrosora,11.61%)和铁芒萁(Dicranopterislinearis,36.50%)>狗脊(21.47%)>绞股蓝(Gynostemmapentaphyllum,12.94%)>黑足鳞毛蕨(Dryopterisfuscipes,4.89%)。
马尾松-甜槠针阔混交林与马尾松纯林的植物群落径级结构存在一定差异(图2)。其中,马尾松-甜槠针阔混交林与马尾松纯林总体径级结构均呈现倒“J”形分布趋势,这表明2个群落结构稳定,更新良好。2种群落类型中主要优势树种的径级结构和种群数量则呈现明显差异,马尾松纯林中马尾松种群结构相对稳定,槲栎则呈现种群增长趋势。马尾松-甜槠针阔混交林中甜槠种群增长趋势,而马尾松的种群呈现“下降型”种群结构,这表明甜槠将随群落演替过程中占据更为优势地位。
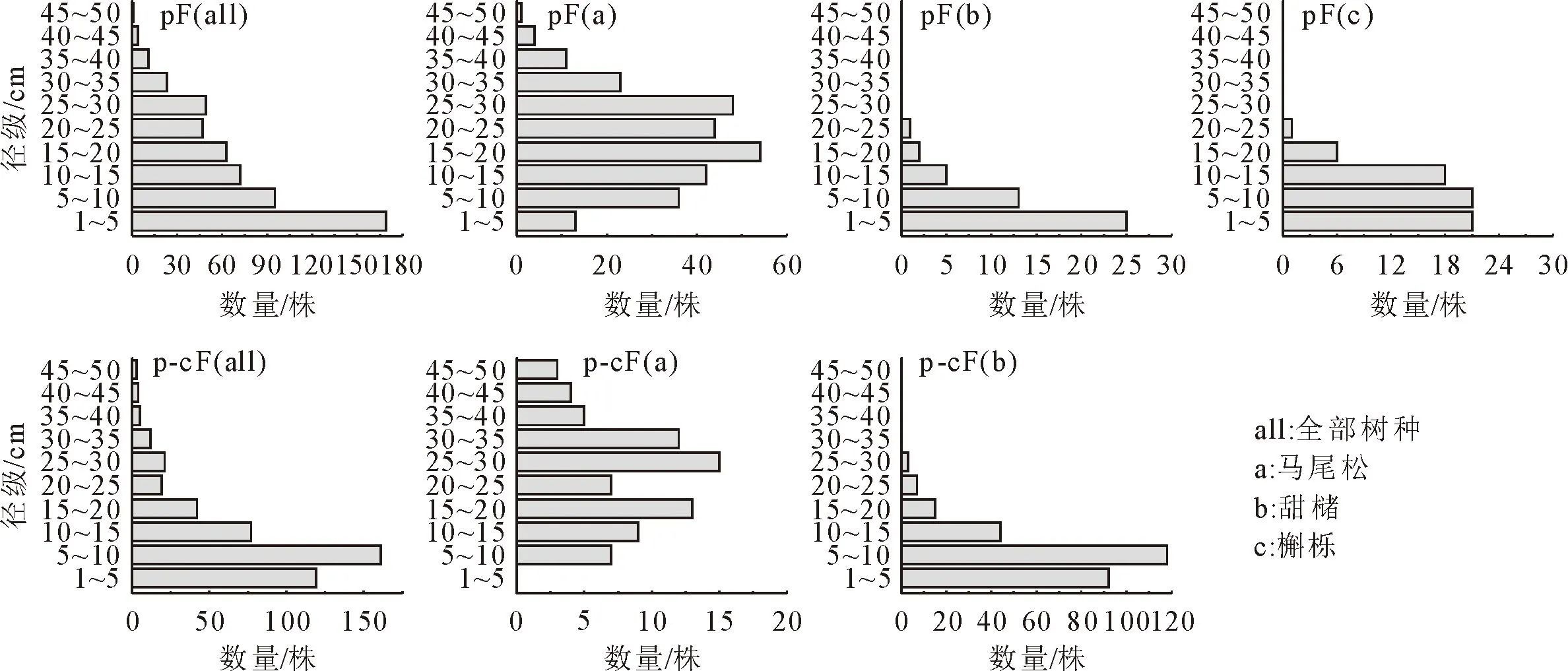
图2 不同样地类型群落径级分布
2.2 主要树种的空间分布格局
图3揭示了生境异质性下主要树种的分布情况。马尾松纯林中马尾松在0~2、7.5~8.5、12.5~14 m属于聚集分布,甜槠在0~2、3~5、11~12 m为聚集分布,在15.5~17 m为均匀分布,槲栎在0~4、5~10 m属于聚集分布,在19~20 m属于均匀分布;马尾松-甜槠混交林中马尾松整体属于随机分布,甜槠整体为随机分布,在0~2 m属于聚集分布。总的来看,马尾松纯林和马尾松-甜槠混交林相应树种在整体上分布格局相似,均以随机分布为主。
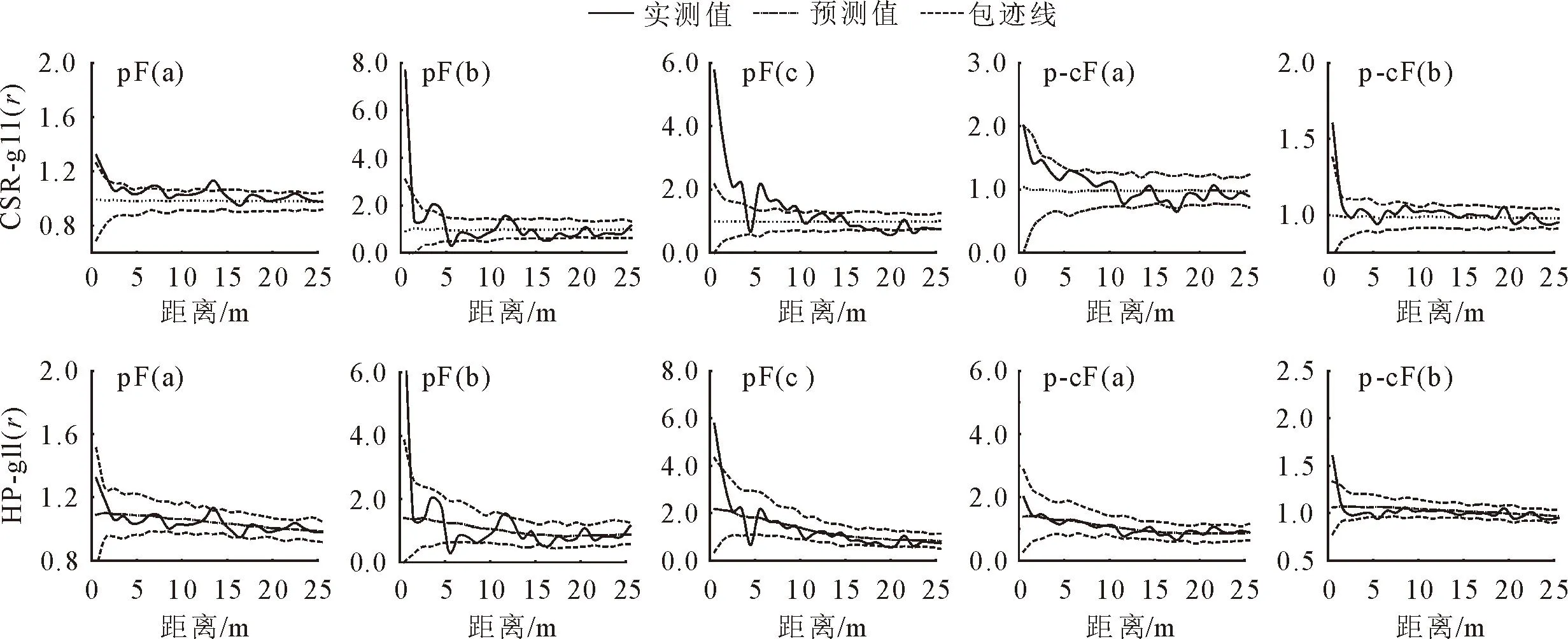
a.马尾松;b.甜槠;c.槲栎
排除生境异质性后,2种林分类型的分布格局与存在生境异质性时相比,出现了较多的随机分布,其中马尾松纯林中槲栎聚集分布的尺度只在0~2 m,其余树种,无论马尾松纯林还是马尾松-甜槠混交林,其聚集尺度均有所下降。通过CSR和HP模型对比发现:生境异质性对树种的空间分布格局有一定影响。
根据径级大小将主要树种胸径划分为3个阶段,利用g11(r)函数分析显示(图4),马尾松纯林和马尾松-甜槠混交林中,幼树、中树阶段和总体的分布格局相似,在小尺度均表现为聚集分布,随着尺度增加转为随机分布;而大树阶段均为随机分布。

s.幼树;m.中树;l.大树;a.全部树木
2.3 主要树种间的空间关联性
利用g12(r)函数,对2个样地主要树种空间关联性进行分析(图5),结果表明马尾松纯林中马尾松和甜槠整体上并未表现出关联性,马尾松和槲栎在2~23 m空间距离上表现为负相关,甜槠-槲栎关联性较为复杂,在2.5、5~6、8~9、13、15 m表现为正相关;而马尾松-甜槠混交林中马尾松-甜槠在5 m为正相关,在7.5~20、24~25 m尺度上为负相关。
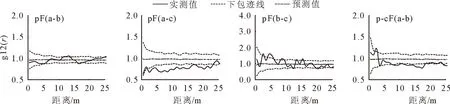
a.马尾松;b.甜槠;c.槲栎
2.4 主要树种径级间的空间关联性
根据g12(r)函数对主要树种不同径级空间关联性进行了分析(图6),结果显示,马尾松纯林中幼树-中树在0~1、12~13 m处为正相关,幼树-大树、中树-大树在所有尺度上均表现为不相关;马尾松-甜槠混交林中,中树-大树在2~3 m处表现为负相关,幼树-中树、幼树-大树在所有尺度上均为不相关。
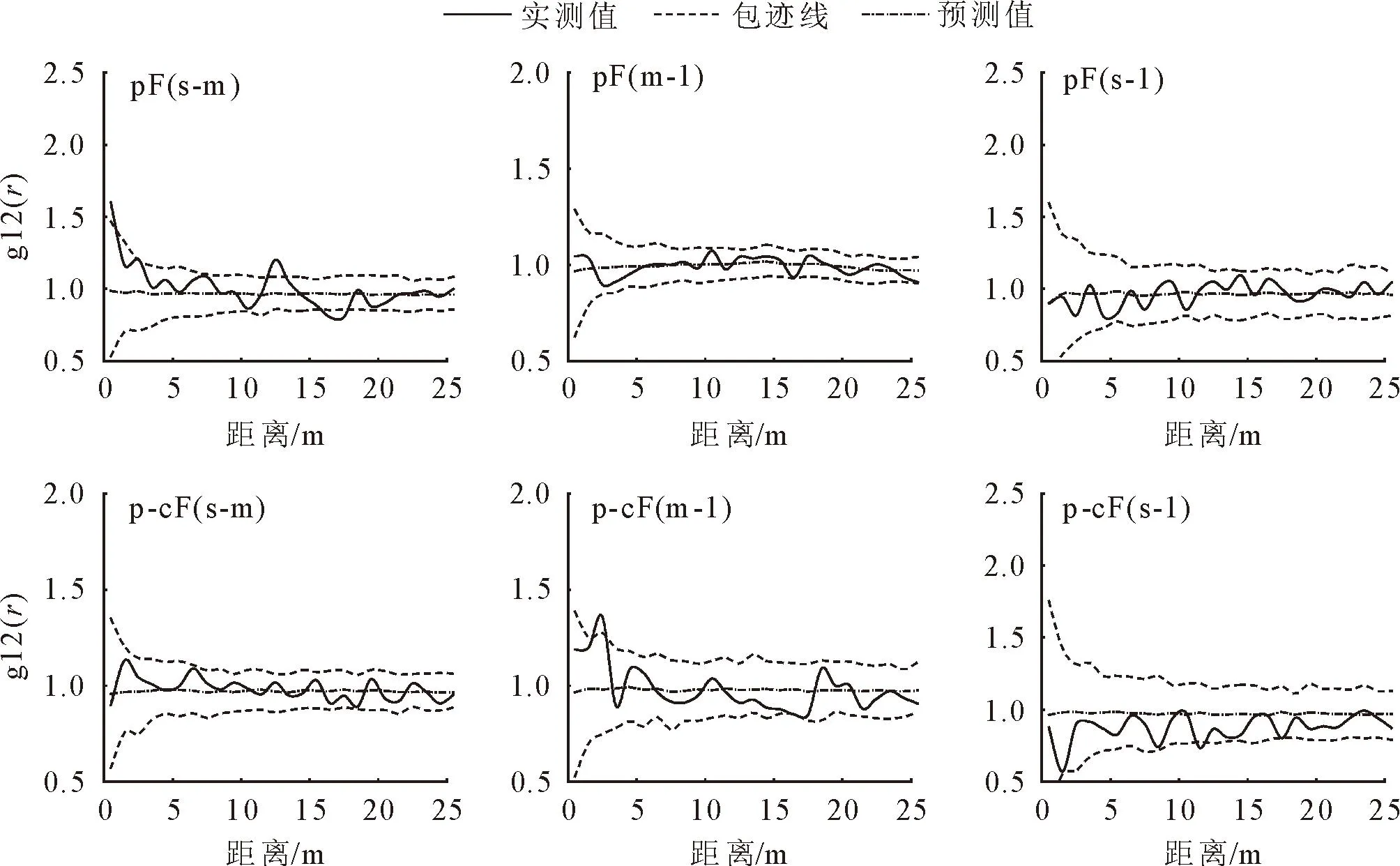
s.幼树;m.中树;l.大树
3 讨论
本研究中马尾松纯林、马尾松-甜槠混交林物种组成丰富,两者乔、灌、草优势种多数不同,两者群落结构有所差异。其中马尾松-甜槠混交林物种数低于马尾松纯林,这可能是因为乔木层群落环境导致林下光照不足,从而使不耐阴物种不能生存,或是耐阴物种间竞争导致其物种丰富度下降。马尾松纯林和马尾松-甜槠混交林群落乔木总径级结构呈现倒“J”形,小径级的树数量充足,说明2个群落结构稳定,更新良好。2块样地马尾松幼树数量低于中树和大树,说明马尾松种群在林分中介于稳定型与衰退型之间,这与已有报道相一致[28]。可能是因为马尾松作为演替前期的先锋物种,能迅速占据优势地位并大量扩散,从而改善了环境条件,为后来物种生存和发展创造了条件。随着演替进行,其逐渐受到竞争抑制,导致径级结构向正态分布靠拢。马尾松-甜槠混交林中甜槠及马尾松纯林中槲栎小径级个体数量多,可能会取代马尾松成为优势种。
黔中山地马尾松-甜槠混交林和马尾松纯林的群落空间分布格局受空间异质性影响作用明显。其中,当考虑生境异质性时,马尾松纯林和马尾松-甜槠混交林群落主要树种在小尺度内聚集分布,而整体呈现随机分布;而当不考虑生境异质性时,尽管2种群落类型样地仍以随机分布为主,但主要树种发生聚集的空间尺度明显下降,这表明样地内的树种分布受到了生境异质性的影响。2种群落类型不同零模型下马尾松种群空间分布均表现为随机分布,这可能是马尾松作为先锋优势物种随演替过程中的“自疏效应”而趋于空间随机分布,以及马尾松的更新受到群落光照限制而趋向随机分布。马尾松纯林中槲栎种群在小尺度内呈现聚集分布,这可能是其更新方式以母株为中心扩散的结果[29]。此外,随着径级的不断增加,马尾松纯林和马尾松-甜槠混交林在小尺度下聚集分布,在中尺度、大尺度下为随机分布,这种格局也出现在其他森林生态系统中[30],可能是因为幼树和中树在小尺度范围聚集能有效降低外界环境的胁迫[31],增加对资源的利用率,提高种间竞争能力,因此利于种群的增长;到了大树阶段,群落中的各物种生态位已固定,密度过高的树因竞争排斥而死亡,最终形成了随机分布的格局。
空间关联分析结果是种群空间关系的呈现,正负关联特征可能是物种间相互作用的结果,也可能是物种生境趋同或趋异的表现。在种群水平,马尾松纯林中马尾松-槲栎大部分尺度呈现负相关,这表明马尾松与槲栎存在竞争关系或者生长环境存在差异,有生态位分离[32],这可能是因为马尾松与槲栎同属喜光物种,彼此对光照、水分等资源的竞争导致其关联性呈现负相关。此外马尾松纯林中马尾松-甜槠、槲栎-甜槠空间关联性为不相关[33],说明在现有群落中这些树种总体上表现为中性关系。在马尾松-甜槠混交林中马尾松-甜槠的关联性在中尺度范围表现为负相关,这可能是因为甜槠作为演替后期物种,开始与马尾松竞争并逐渐取代马尾松成为群落的优势种。不同径级种间关联性的差异可能受植物大小的影响,个体间植株大小的差异越大,个体间的正相关性就越弱[34]。在马尾松纯林和马尾松-甜槠混交林中,幼树-中树、中树-大树、幼树-大树在研究尺度上均趋于不相关,这可能是因为幼树、中树与大树在空间上分布为随机分布,且幼树和中树对光资源、土壤养分及水分等资源的竞争力微弱造成的[35]。随着径级增加,树种对环境资源的获取和控制能力逐渐增加,导致小径级个体大量死亡,个体间距离增加,正相关性减弱,幼树-大树种间关联性更趋向于负相关和不相关。
4 结论
通过对黔中马尾松-甜槠针阔混交林的研究发现,黔中山地典型的马尾松-甜槠针阔混交林和马尾松纯林群落空间格局分布存在明显差异。生境异质性是影响该区马尾松针阔混交林树种空间分布格局的一个重要因素,在今后研究中还应考虑环境因子(温度、地形、土壤养分和光照等)和空间格局的关系。本研究建议在该区马尾松纯林阔叶化改造过程中,可采取适当的采伐措施(如:择伐死亡树木、距离相近的大径级马尾松、修剪枝叶等方法),以改善马尾松与其他阔叶树种间(如马尾松-槲栎、马尾松-甜槠)的负相关关系,进而促进阔叶树种(如槲栎、甜槠)的生长。
