落叶松-杨栅锈菌MlpMCM4蛋白和MlpHOG1蛋白互作关系初探
2024-01-20陈凯玥李子晔周显臻
杨 冰,陈凯玥,李子晔,周显臻,于 丹
(西北农林科技大学 林学院,陕西 杨陵 712100)
微小染色体维持蛋白(minichromosome maintenance protein,MCM)是AAA ATPase大基因家族中一类独特的亚群[1]。MCM蛋白在真核生物中高度保守,典型的MCM基因家族包括6个成员,即MCM2、MCM3、MCM4、MCM5、MCM6和MCM7,它们通常形成异源六聚体复合物[2]。MCM蛋白具有ATPase结构域,也称作MCM box,其中含有所有MCM蛋白具有的3个保守基序。第1个Walker A,对应MCM特异的序列GDPXX(S/A)KS;第2个Walker B,序列为IDEFDKM;第3个R-finger,序列较短,SRFD[1,3]。所有真核MCM蛋白具有锌结合基序,具体序列因蛋白类型有一定差异[1]。在酿酒酵母中,MCM基因家族在DNA复制起始过程发挥重要作用,并且以0.4 M NaCl处理细胞10 min作为外源渗透压胁迫,使用免疫共沉淀技术检测HOG1蛋白是否与各种复制复合体组分相互作用,最终发现HOG1能够与MCM4、CDC45、PSF2、DPB2及CDC7互作[4-5]。
在真菌中,MCM4基因的研究主要集中在酿酒酵母(Saccharomycescerevisiae)和裂殖酵母(Schizosaccharomycespombe)两大系统,也被分别称为CDC54和CDC21[1]。付静等[3]在全基因组水平上鉴定了植物病原真菌葡萄座腔菌(Botryosphaeriadothidea)MCM基因家族成员,并分析了其对硫化物胁迫的响应特征,为阐明韭菜及其主要成分对苹果轮纹病的防控机理提供了理论依据。专性寄生真菌落叶松-杨栅锈菌(Melampsoralarici-populina)能够侵染杨树引起杨树叶锈病,该病害广泛分布于世界各杨树栽培区,在我国对杨树用材林和防护林的安全生产造成严重影响[6]。落叶松-杨栅锈菌MCM基因家族的研究目前尚未见报道。前期研究推断MlpHOG1基因参与落叶松-杨栅锈菌侵染菌丝生长扩展和响应环境胁迫等生命活动[7-8]。本研究通过鉴定落叶松-杨栅锈菌标准菌株MCM基因家族成员和系统发育分析,从而确定MCM4候选基因,进而同源克隆获得中国菌株MlpMCM4基因的CDS片段,并对目的蛋白的基本特征进行预测分析,最后借助2种技术分析目的蛋白与MlpHOG1蛋白的相互作用情况,为后续明确目的蛋白与MlpHOG1蛋白的互作关系提供帮助。
1 材料与方法
1.1 试验材料和试剂
1.1.1 试验材料 落叶松-杨栅锈菌中国菌株wh03为单孢菌系[9],使用太白杨(Populuspurdomii)在温室进行人工扩繁,根据曹支敏等[10]的方法进行菌株的活化及保存。酵母双杂交载体pDHB1、pPR3-N、pDHB1-largeT和pDSL-ΔP53以及荧光素酶试验载体pCAMBIA1300-CLuc、pCAMBIA1300-NLuc分别由西北农林科技大学植物保护学院刘慧泉教授和康振生教授实验室惠赠。大肠杆菌感受态细胞DH5α购自北京擎科生物科技有限公司。酵母感受态细胞NMY51和农杆菌感受态细胞GV3101购自上海唯地生物技术有限公司。
1.1.2 主要试剂 总RNA提取试剂盒购自QIAGEN公司;反转录试剂盒、Phusion Plus DNA聚合酶和Taq酶购自Thermo Scientific公司;高保真聚合酶Phanta Max Super-Fidelity DNA Polymerase公司和Clon Express Ⅱ One Step Cloning Kit购自Vazyme;pMD19-T载体购自TAKARA公司;SC/-Leu/-Trp Broth、SC/-Ade/-His/-Leu/-Trp Broth和X-gal购自北京酷来搏科技有限公司;D-荧光素钾盐购自上海笛医生物科技有限公司。引物序列合成和大肠杆菌转化子测序在北京奥科鼎盛生物科技有限公司(杨陵分部)完成。
1.2 试验方法
1.2.1 候选基因分析 从Pfam数据库(http://pfam.xfam.org/)下载MCM基因家族模型数据(PF00493),通过hmmer软件在供试真菌(表1)全体蛋白序列中搜索鉴定,提取鉴定到的序列并在NCBI CDD数据库(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)中比对,从而去除不存在MCM(PF00493)保守结构域的序列。将通过筛选的蛋白序列用软件MEGA7.0对齐,用gblocks提取保守序列。最后,用iqtree构建最大似然法(Maximum likelihood)树,设置“-m”参数调用ModelFinder获得氨基酸序列的最佳替代模型,设置“-b 1 000”采用1 000次重复计算的常规自展检验法(Bootstrap)对分支可靠性进行评估。生成的树文件使用MEGA7.0软件查看。

表1 参与MCM基因家族鉴定的物种Table 1 Species used in the phylogenetic analysis of the MCM family
1.2.2 目的基因克隆 用Rneasy Plant Mini Kit试剂盒提取落叶松-杨栅锈菌夏孢子总RNA,消解基因组DNA后用Thermo Scientific Revert Aid First Strand cDNA Synthesis Kit试剂盒合成第一链cDNA。根据落叶松-杨栅锈菌标准菌株98AG31(v1.0)[11]中Protein ID 48743的基因序列,利用软件Primer Premier 5设计特异性扩增引物48743-uF1和48743-uR2(表2)。PCR 反应总体系50 μL,其中cDNA模板1.5 μL,10 μM上下游引物各2.5 μL,2×Phusion Plus PCR Master Mix 25 μL,ddH2O 18.5 μL。扩增程序:98 ℃预变性30 s;98 ℃变性7 s,60 ℃退火10 s,72 ℃延伸1 min 25 s,35个循环;72 ℃延伸 5 min,16 ℃保存。扩增完成后,在PCR产物中加入0.5 μLTaqDNA聚合酶和0.5 μL dNTP,72 ℃延伸30 min,16 ℃保存。用1.0%琼脂糖凝胶电泳检测PCR扩增产物,然后用琼脂糖凝胶DNA回收试剂盒纯化目的片段,再连接至pMD19-T载体骨架,转化大肠杆菌DH5α感受态细胞,PCR检测筛选后将阳性转化子送公司测序。
1.2.3 目的基因编码蛋白生物信息学分析 利用在线软件ProtParam预测和分析目的基因编码蛋白的基本理化性质。使用软件Clustalx1.83对供试蛋白序列进行多序列比对,然后使用GeneDoc软件进行查看。使用SOPMA在线工具预测目的基因编码蛋白的二级结构。借助3种在线软件EuK-mPLoc 2.0、WOLFPSORT和PROTCOMP预测分析目的蛋白的亚细胞定位区域。
1.2.4 酵母双杂交 使用高保真聚合酶扩增目的片段,扩增引物见表2。NcoⅠ单酶切pDHB1载体,BamH Ⅰ和EcoR Ⅰ双酶切pPR3-N载体。用重组克隆试剂盒,将MlpMCM4(wh03)基因片段和MlpHOG1(wh03)基因片段分别连接至pDHB1载体质粒和pPR3-N载体质粒,测序确认后获得bait载体pDHB1-MlpMCM4和prey载体pPR3-N-MlpHOG1。将获得的重组载体质粒共转化至酵母感受态细胞NMY51,观察30 ℃条件下整合外源片段的酵母转化子在缺陷培养基SD-Trp-Leu和SD-Trp-Leu-His-Ade上培养3 d的生长情况,并检测以X-gal为反应底物时能否发生蓝色反应。其中pDHB1-largeT与pDSL-ΔP53共转化转化子为阳性对照,pDHB1-largeT与pPR3-N共转化转化子为阴性对照,pDHB1-MlpMCM4与pPR3-N共转化转化子为自激活组。
1.2.5 萤火虫荧光素酶互补试验 使用高保真聚合酶扩增目的片段,扩增引物见表2。KpnⅠ和SalⅠ酶切空载体pCAMBIA1300-CLuc和pCAMBIA1300-NLuc。用重组克隆试剂盒将目的基因片段分别连接至载体骨架,测序确认后获得重组载体pCAMBIA1300-CLuc-MlpMCM4和pCAMBIA 1300- NLuc-MlpHOG1。选取生长3~4周、长势良好的本氏烟(Nicotianabenthamiana)作为烟草瞬时表达系统的材料。将获得的重组载体质粒分别转化至农杆菌感受态细胞GV3101,参考Chen等[12]的方法进行菌液处理,在烟草叶片上分别注射不同的菌液组合。2 d后在每个接种点均匀涂抹D-荧光素钾盐溶液,室温黑暗静置5 min。在多光谱动态荧光显微成像系统(PlantView 100)中观察其注射烟草叶片区域是否有荧光出现。
2 结果与分析
2.1 候选基因的确定
借助hmmer软件将下载获得的MCM基因家族模型数据在10个供试物种的蛋白序列中进行搜索鉴定,共获得65条序列。其中2个基因序列太短,不具有保守结构域,进而使用63条序列构建最大似然法系统进化树,结果表明,进化树具有6个分支,分别代表MCM基因家族的MCM2、MCM3、MCM4、MCM5、MCM6和MCM7(图1)。ID 48743(v1.0)为酿酒酵母MCM4基因在落叶松-杨栅锈菌标准菌株98AG31中的直系同源基因,命名MlpMCM4。
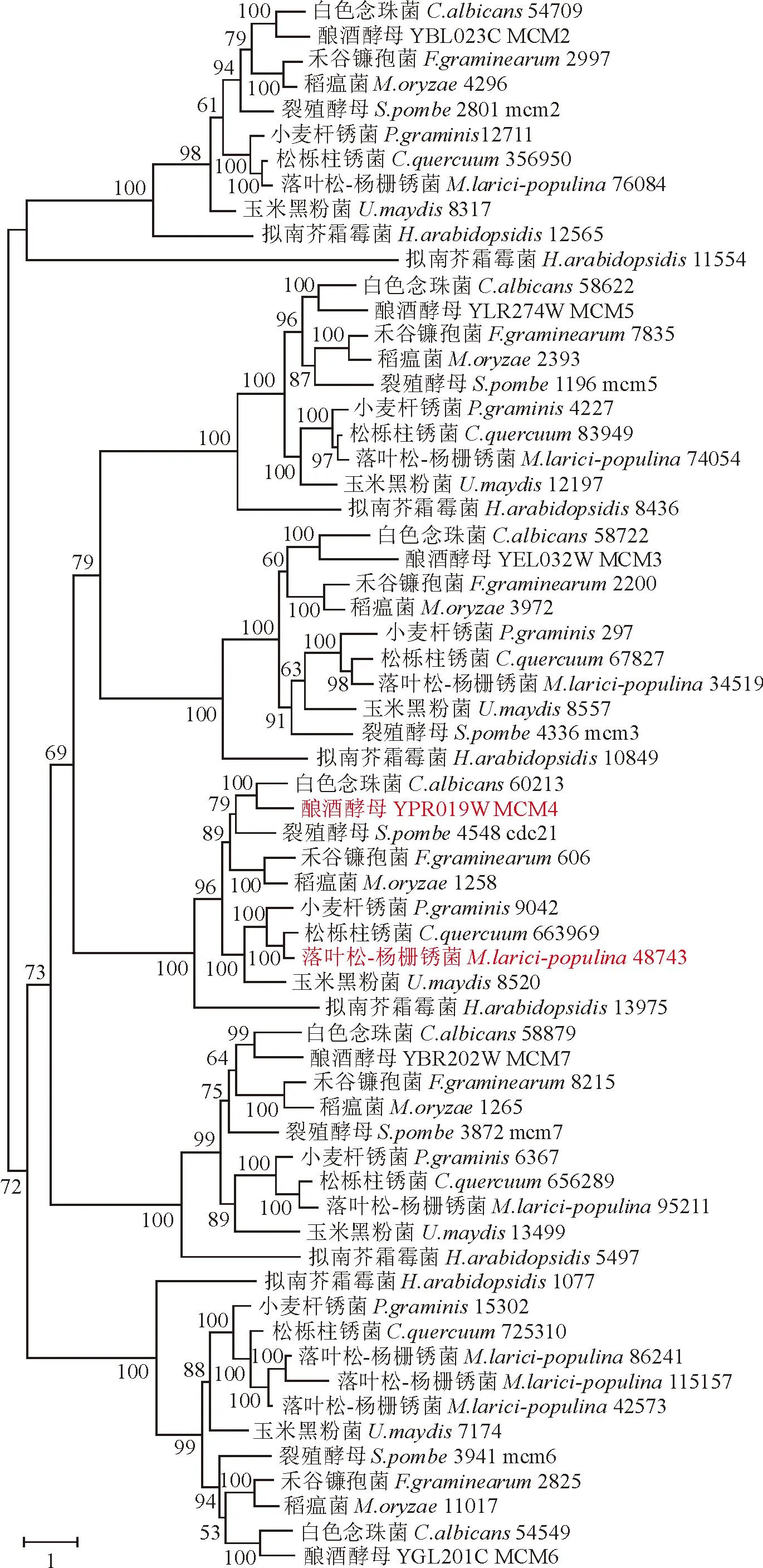
1)物种后的数字代表在相应物种数据库中的蛋白序号(Protein ID); 2)自展支持率标注于节点。
2.2 目的基因的克隆
以落叶松-杨栅锈菌中国毒性菌株wh03(图2A)夏孢子cDNA为扩增模板,根据ID 48743(v1.0)序列设计特异引物进行同源克隆,扩增结果呈现一个较为特异且浓度较高的大小约2.5 kb的条带(图2B)。通过TA克隆及测序分析结果表明,成功获得wh03菌株ID 48743基因的CDS片段,长度为2 460 bp,将该基因命名为MlpMCM4(wh03)(GenBank OR751401)。
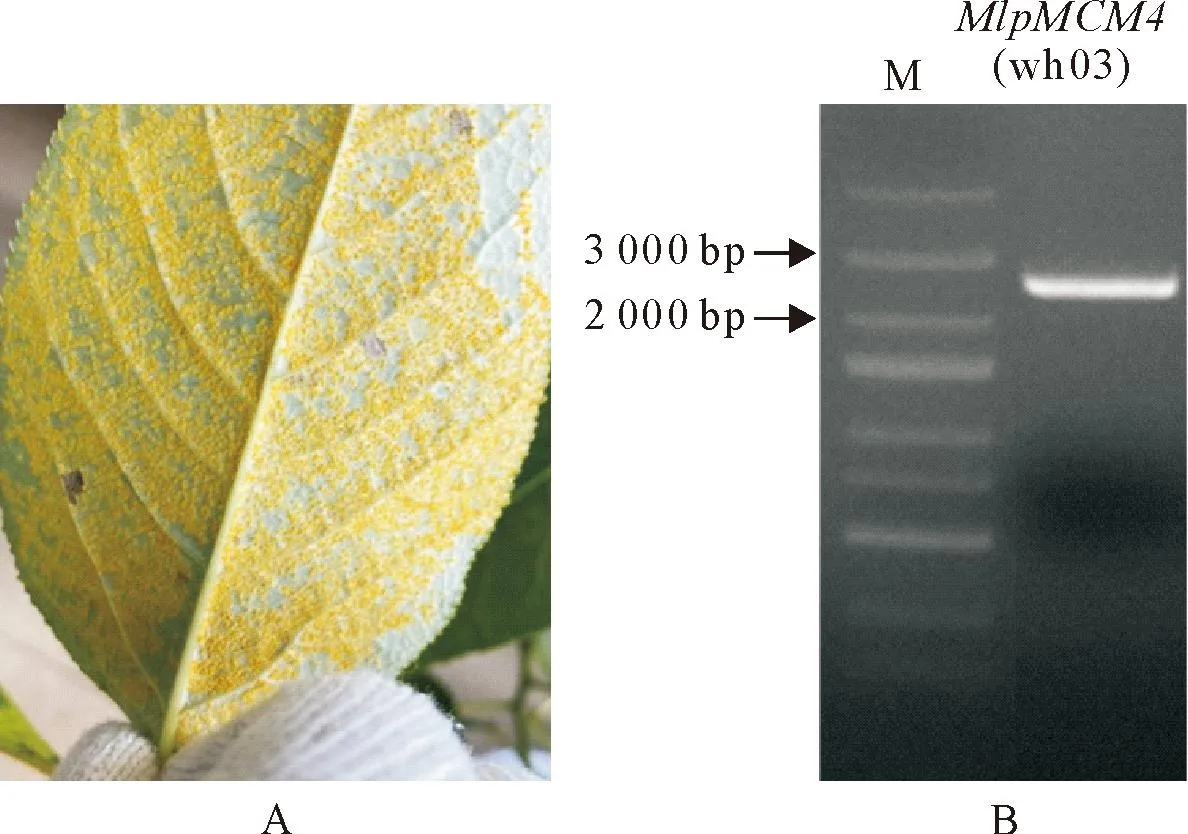
A.落叶松-杨栅锈菌中国毒性菌株wh03接种杨树叶片后的寄主表型;B.从wh03菌株夏孢子cDNA中扩增MlpMCM4基因片段
2.3 目的基因编码蛋白特征
使用在线软件ProtParam分析蛋白理化性质,结果显示MlpMCM4(wh03)基因编码819个氨基酸,相对分子量90.31 ku。负电荷氨基酸残基(Asp+Glu)总数为110个,正电荷氨基酸残基(Arg+Lys)总数为97个,带负电荷的数目略多于正电荷。MlpMCM4(wh03) 基因编码蛋白理论等电点(pI)为5.78,即偏酸性。预测的不稳定指数为44.56,超过40,推测属于不稳定蛋白。
将MlpMCM4(wh03)基因编码蛋白序列和酿酒酵母MCM4蛋白序列进行比对分析,发现目的蛋白具有MCM蛋白共有的3个重要基序,即Walker A(GDPXX(S/A)KS)、Walker B(IDEFDKM)和 R-finger(SRFD),还具有MCM4类型锌结合基序(CX2CX18CX2-4C)(图3)。

1)深绿色横线区域隶属酿酒酵母MCM4的MCM box结构域,其中浅蓝色横线表示Walker A、Walker B和 R-finger基序;2)*表示锌结合基序。
运用SOPMA在线预测目的蛋白二级结构,显示MlpMCM4 (wh03)蛋白二级结构主要由4种形式组成(图4),分别为41.64%的α-螺旋(Alpha helix)、6.23%的β-转角(β-turn)、15.26%的延伸链(Extended strand)和36.87%的无规则卷曲(Random coil),其中α-螺旋和无规则卷曲为主要二级结构。
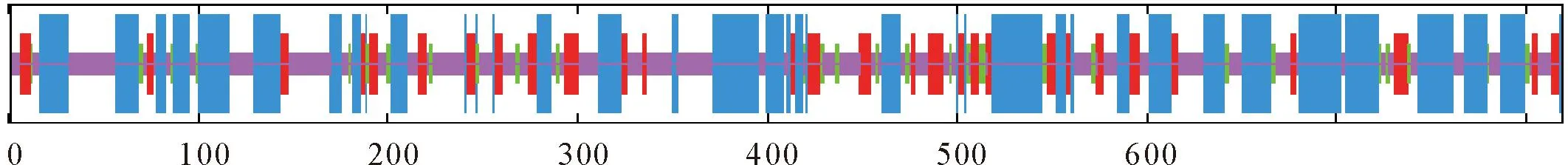
蓝色:α螺旋;绿色:β-转角;红色:延伸链;紫色:无规则卷曲
使用EuK-mPLoc、WOLFPSORT和PROTCOMP这3种在线软件对MlpMCM4 (wh03)蛋白进行亚细胞定位预测分析。EuK-mPLoc预测目的蛋白定位在细胞核(Nucleus)区域。WOLFPSORT和PROTCOMP均预测显示细胞核(Nuclear)区域数值明显高于其他区域(26/27和8.84)。综合3种软件预测结果,推断MlpMCM4 (wh03)蛋白定位在细胞核区域。
2.4 MlpMCM4蛋白和MlpHOG1蛋白的互作关系
借助酵母双杂交技术分析落叶松-杨栅锈菌MlpMCM4蛋白和MlpHOG1蛋白是否相互作用。在SD-Leu-Trp培养基上,阳性对照(pDHB1-largeT+pDSL-ΔP53)、阴性对照(pDHB1-largeT+pPR3-N)及自激活组(pDHB1-MlpMCM4+pPR3-N)均能正常生长。在SD-Leu-Trp-His-Ade培养基上,阳性对照正常生长,阴性对照不能生长,自激活组也不能生长;X-gal染色显示阳性对照变蓝,阴性对照没有变蓝,自激活组也没有变蓝(图5A)。因此,MlpMCM4(wh03)基因编码蛋白不具有自激活能力。阳性对照、阴性对照及试验组(pDHB1-MlpMCM4+pPR3-N-MlpHOG1)在SD-Leu-Trp培养基上均能正常生长。在SD-Leu-Trp-His-Ade培养基上,阳性对照正常生长,阴性对照不能生长,试验组也不能生长;X-gal染色显示阳性对照变蓝,阴性对照没有变蓝,试验组也没有变蓝(图5B)。因此,利用本研究中分离泛素酵母双杂交系统发现MlpMCM4(wh03)基因编码蛋白和MlpHOG1(wh03)基因编码蛋白二者不存在相互作用关系。

A.自激活检测结果;B.试验组检测结果
同时,借助萤火虫荧光素酶互补试验进行分析。在植物活体荧光成像系统中观察不同组合烟草叶片注射区域,阳性对照pCAMBIA1300-CLuc-X与pCAMBIA1300-NLuc-Y共表达的叶片区域呈现荧光(结果未报道),3组阴性对照pCAMBIA1300-CLuc+pCAMBIA1300-NLuc、pCAMBIA1300-CLuc-MlpMCM4+pCAMBIA1300-NLuc、pCAMBIA1300-CLuc+pCAMBIA1300-NLuc-MlpHOG1,以及试验组pCAMBIA1300-CLuc-MlpMCM4+pCAMBIA1300-NLuc-MlpHOG1共表达的叶片区域都没有检测到荧光信号(图6)。表明该系统中MlpMCM4(wh03)基因编码蛋白和MlpHOG1(wh03)基因编码蛋白也不存在相互作用。

图6 萤火虫荧光素酶互补试验分析MlpMCM4蛋白和MlpHOG1蛋白的互作关系Fig.6 The interaction between MlpMCM4 andMlpHOG1 detected by the firefly luciferase complementary assay
3 结论与讨论
MCM4基因是MCM家族成员之一,一对一比对涉及的序列相似度较高,因此根据家族模型在全基因组水平鉴定落叶松-杨栅锈菌MCM基因家族成员,进而通过构建系统进化树来确定成员直系同源基因。本研究发现ID 48743是酿酒酵母MCM4基因的直系同源基因,因此命名为MlpMCM4。同时,也鉴定出其他成员,ID76084是酿酒酵母MCM2基因的直系同源基因,ID34519是酿酒酵母MCM3基因的直系同源基因,ID74054是酿酒酵母MCM5基因的直系同源基因,ID86241、115157和42573是酿酒酵母MCM6基因的直系同源基因,ID95211是酿酒酵母MCM7基因的直系同源基因。这为后续研究该锈菌MCM基因家族的功能奠定基础。本研究使用MCM模型在供试担子菌和子囊菌中均鉴定到MCM2-7,没有其他成员如MCM8和MCM9,这与MCM8和MCM9仅存在于高等生物体内的报道是一致的[13]。
比对分析显示MlpMCM4 (wh03)蛋白具有3个保守的亚结构域,即Walker A、Walker B和R-finger,它们与MCM蛋白利用ATP的能力密切相关[13]。同时,目的蛋白还具有MCM4类型特有的锌指结构(CX2CX18CX2-4C),研究显示在许多MCM蛋白中该结构与酵母菌株的存活密切相关[14-15]。因此,比对预测分析显示目的蛋白较为保守。大多数MCM4蛋白还具有一个保守的CDK激酶作用位点(S/T)PX(K/R)[1]。但没有在目的蛋白中预测出该位点,推断可能CDK激酶通过新的途径或者作用复合体其他基因来发挥作用。酿酒酵母6个MCM蛋白成员在G1期主要集中在细胞核区域,在S期逐渐从核内输出,到了G2/M期则完全从核内排出[16]。本研究亚细胞定位预测显示目的蛋白定位于细胞核,这与报道具有吻合度,有待于后续开展相关试验进行分析。
落叶松-杨栅锈菌是专性寄生真菌,目前还没有稳定遗传操作体系,导致致病分子机理研究明显落后于模式植物病原真菌。基于前期研究进展,期望能够理清落叶松-杨栅锈菌HOG途径在该病原菌侵染致病和响应胁迫方面的作用机制。酿酒酵母中在外源渗透压处理下HOG1和MCM4蛋白具有特异的互作关系[5]。本研究酵母双杂交系统和萤火虫荧光素酶互补试验中,落叶松-杨栅锈菌中国菌株HOG1蛋白和MCM4蛋白没有表现相互作用,推断可能因为没有外源渗透压刺激。后续考虑增加外源渗透压胁迫条件及借助多种互作检测方法,来继续分析2个蛋白的互作关系。
