森林粗木质残体分解因素的研究进展
2024-01-20彭凡茜耿庆宏刘文芳张惠光
彭凡茜,耿庆宏,2,刘文芳,周 艳,张惠光,杨 文,徐 侠,*
(1.南京林业大学 生物与环境学院,南方现代林业协同创新中心,江苏 南京 210037;2.西北农林科技大学 林学院,陕西 杨陵 712100;3.武夷山国家公园科研监测中心,福建 武夷山 354300;4.福建武夷山森林生态系统国家定位观测研究站,福建 武夷山 354300;5.陕西师范大学 生命科学学院,陕西 西安 710119)
大量的碳(C)储存在森林中,其中8%储存在木质残留物中[1-2]。粗木质残体(coarse woody debris,CWD)对森林功能有重要的“来世”影响,在陆地生态系统的C循环中起着关键作用[3]。但迄今为止,研究人员对森林生态系统中的分解研究大部分集中在叶片和细根部分,对森林中CWD分解过程的研究仍然十分有限[4]。CWD分解过程导致固定在植物中的C和营养物质通过不同的形式重新进入生态系统循环,这种C和营养物质无论固定还是释放都将对土壤C和营养物质储存产生重大影响[5-7]。CWD的命运将影响森林C平衡[2,8]。因此,为了进一步深入理解和准确预测森林C循环的变化动态,研究CWD的分解情况十分必要。
CWD的分解为大部分土壤生物提供了关键资源,充当短期的库和长期的营养源[9]。相较于叶片凋落物来说,CWD具有体积大、结构复杂、分解和养分释放过程十分缓慢的特点,使得其在森林中逐渐积累。目前我国的CWD的研究大部分集中在生态功能、储量等方面[10-14],而对影响CWD分解因素的了解尚少。本研究基于国内外相关领域文献查阅与分析,总结了影响CWD分解的主要因素,以期为全球变化背景下改进C和养分循环模型提供理论依据。
1 CWD概念
CWD的分解过程影响着C、氮(N)等养分循环[15]。Harmon等[16]整合了温带生态系统中CWD的研究,将CWD定义为“直径≥2.5 cm的死木质物”。20世纪90年代,美国的相关部门(农业部林务局和长期生态研究站)制定了统一标准,将CWD定义为“直径(一般指较粗的一端)≥10 cm,长度≥1 m的死木质残体”。除了对CWD的尺寸特征进行划分外,对CWD中所包含的内容也进行了划分。侯平等[6]将倒木、枯立木、大型凋落枝条、根桩(含伐后树桩)、地表和地下大型死根都包括进了CWD中,闫恩荣等[17]将CWD的定义概括为“较粗部分的直径≥10 cm,长于1 m的倒木、枯立木、粗枝或长度<1 m的树桩,且还包括直径>1 cm的残根”。
2 影响CWD分解的因素
CWD的分解过程与凋落叶的分解相似,主要包括浸出、破碎、呼吸和生物转化[18]。也有研究者将木质碎片的分解分为浸出溶解有机碳(DOC)、燃烧、物理降解、微生物分解以及昆虫分解5种途径。木质碎片中的纤维素、半纤维素和木质素等将会转化成更小的单元并最终矿化,以无机的形式回到环境中[2]。环境因素(温度和水分)、分解微生物和植物凋落物性状通常被用于预测凋落物的分解[19]。Swift等[20]提出了关于影响分解的三角形模型,模型假设环境、生物分解者和凋落物性状是控制分解速率的重要因素,将环境主要分为气候和土壤因素,并强调了气候因素的首要性。近期也有观点认为,凋落物性状在广阔的空间尺度上是主要的控制因素。Bradford等[21]修改了经典的三角模型(图1),认为不应该错误地夸大气候在分解中的作用,但也不能片面地认为只有其中的某一个方面控制着分解速率。随着分解的不断进行,木质的改变影响其周边的微环境,且参与分解的生物类群也在不断发生改变。但究竟是何种因素主导了分解过程,目前影响分解因素的相对主导性仍存在争论。本研究将从环境、分解者和木材本身特性3个方面入手,综述这些因素对CWD分解的影响。
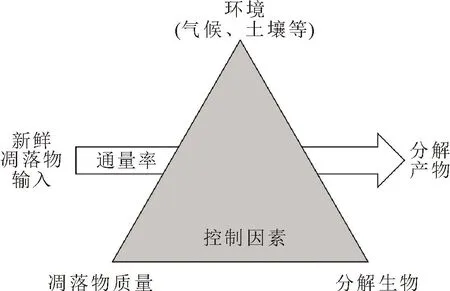
图1 根据Bradford等[21]的调控凋落物分解速率控制因素修改的分解三角形Fig.1 The decomposition triangle modified according to the control factors regulating the decomposition rate of litter by Bradford et al.[21]
2.1 环境变化
环境变化对分解的影响通常可以分为全球尺度和局域尺度。传统上,气候变化被认为是全球尺度上分解速率的主要控制因素,木材腐烂过程很大程度会受到温度、湿度的影响[22]。温度和水分变化可以通过影响CWD的呼吸来影响木材分解[23-24]。Gough等[25]研究发现粗木质碎片呼吸(RCWD)随着温度和含水量的增加而增加。Forrester等[26]发现CWD产生的CO2通量与木材温度呈正相关。Liu等[27]研究发现,随着温度和水分增加CWD的CO2的释放速率不断提高。Pietsch等[28]研究发现,木材分解受温度的控制比受树种和分解者多样性的控制更为强烈。有利的气候条件(适宜的温度和充足的湿度)有助于分解者的丰富度和活性,从而解释有机物的分解速率[29]。此外,在整个气候梯度中,太阳辐射增强也会显著影响CWD分解。Wu等[30]的研究发现,暴露于增强的光降解作用下,木荷、杉木和香樟3种树的CWD分解明显加速。
局域尺度上的分解速率受到森林组成、年龄分布和结构等方面的影响[31-32]。空间异质性是森林CWD的主要特征,处于不同位置的CWD会产生复杂的空间分布,特别是直立木材和倒下的木材。Romero等[33]在红树林中进行了木材分解的研究,用木圆盘模拟了CWD在不同位置(埋于地下、地面、空气中)分解的状况,发现在土壤表面分解的圆盘其衰减率高于埋在地下的圆盘,但两者的衰减率都比空气中的圆盘高。同样,Erdenebileg等[34]的研究也证明了埋藏于土壤中的木质残体质量损失比处于地表和空气中的快。此外,不同生境的环境和养分状况不同[35],也会影响到木质残体的分解过程。Jones等[36]的研究表明,南洋樱在溪流中的分解速度比陆地快,可能是因为河流的冲击和淋溶导致更快的质量损失。
2.2 分解者
2.2.1 土壤动物 在分解早期,土壤动物起着重要作用。土壤动物可以将大块的有机物切割、粉碎成更小的单元。有研究表明,土壤动物所分解的木质残体占每年枯木质量损失的10%~20%[37]。尽管许多研究人员提出土壤动物在木材的分解过程中扮演着重要角色[38-40],但目前这方面的研究仍然未受到重视。
土壤动物通过自身产生内源消化酶以及通过与真菌、原生生物和其他生物之间的共同作用,促进木材的降解[37]。土壤动物分解木材的方式主要分为2种,一种是以木材为食,能够消化木材;另一种以破坏木材为主,其破坏木材的目的是养殖真菌或筑巢[37]。Hyodo等[41]研究发现,白蚁通过其共生的真菌分解木质素,拥有外共生的真菌使白蚁比依赖肠道微生物分解木材的昆虫具有更快的分解能力,尤其是对木质素的降解[42]。不同种类的土壤动物对CWD的分解贡献也不同。美国东南部的一项研究发现,白蚁在2 a内消耗的木材量是其他所有昆虫总和的6倍[43]。此外,分解木材的大部分土壤动物是捕食者或寄生者,这些动物可以通过改变生物之间的活动或者群落特征来影响分解速率。一些土壤动物会通过挖掘隧道的方式穿过树皮去获取树皮下方有营养的木质层,而进入韧皮部通常需要钻穿保护树的外层形成孔隙,其他的有机体可以通过这些孔隙进入,进而加速木材分解[37]。一项关于凋落叶的研究也证明,蚂蚁可能通过减少以微生物为食的无脊椎动物数量来加速凋落叶的分解[44]。总而言之,土壤动物的主要作用发生在分解的早期,其分解CWD主要通过两方面,一方面以木材作为食物或与其共生的微生物之间相互作用对木材进行分解,另一方面则是直接破坏木材,对大块的木材进行破碎化处理,增大木材的接触面积从而促进了分解。目前研究土壤动物不同种群之间的相互作用对CWD分解的影响仍然较少,在今后的研究中应该加强关注。
2.2.2 微生物 微生物的活动贯穿于木材分解的始终,能够促进生态系统中C和养分循环,并为其他生物群提供资源[45-46]。微生物的一个重要特征就是能够释放酶,酶能够将复杂的底物分解成足够小的化合物,是影响分解的重要条件之一[47]。另一方面,微生物在分解过程中由于资源竞争导致的群落结构变化也将会影响分解速率[48-50]。Hu等[51]采用时间序列法研究了杉木木桩的C损失率与微生物群落组成的关系,结果表明,在木材含水量较低的早期,真菌表现为优势群落,但在末期,细菌生物量则呈上升趋势。微生物群落组成的不同也将导致分解速率的不同[45,52]。有研究发现真菌之间的相互作用会降低C利用效率[53]。Hiscox等[54]发现,木块中生长的担子菌之间的相互作用推动了腐烂木质中真菌群落的变化。除此之外,真菌本身的特性也能够解释木材分解[55]。真菌主要分解有机物中难降解的木质素和细胞壁,其可分为内生真菌和外生真菌[56]。内生真菌存在于树木成活期间,当树木死亡后,内生真菌则发挥“优势效应”。即内生真菌本身已经在木材中定殖,这些真菌通常很快占据主导地位开始分解死亡的木材。Cline等[57]研究木材灭菌处理和未处理对分解的影响,发现木材的内生菌能够影响分解早期的微生物群落组成以及腐烂率,并增强枯落物的质量损失。与非灭菌处理相比,外来的菌种需要重新定殖无菌木材,从而导致参与分解的时间滞后。内生菌的存在不仅可能影响木材分解的微生物群落的组成,而且还间接促进了后来外生菌的定殖。随着木材腐烂程度加深,优势种逐渐由内生真菌转变为外生真菌[58]。此外,还有研究发现,内生菌和后来到达的物种之间可能会产生拮抗作用导致分解速率降低[59-60]。
2.3 木材特性
木材性状代表着植物功能变异的重要方面,木材往往比其他植物组织(如叶、花、果实等)有更多的营养限制。在给定的气候条件下,木材分解速率与一系列木材特性相关,木材本身的理化性质影响分解的过程[61-62]。物种影响分解的差异幅度比以前认为的大得多,并且大于气候因素引起的变化[63]。Kahl等[64]的研究发现,裸子植物的平均衰变率显著低于被子植物,这可能是因为裸子植物木质素中的愈创木基结构和被子植物不尽相同,木质素通常难以分解,较高的木质素含量会限制分解的速率。并且由于裸子植物的树皮和木质部组织中都有发达的树脂管系统,这些组织中主要包含萜类和酚类化合物等,这些物质能够起到保护作用[65]。Weedon等[66]对来自世界各地的分解研究进行了整合分析,同样也表明了被子植物的分解速度快于裸子植物。此外,木材的初始性状对分解速率也起着重要的调控作用。多数研究表明,木材初始N、木质素氮比、碳氮比和直径等都是解释分解速率变化重要因素[62,67]。
即使在同一根原木上,物理化学性质也会在空间上存在差异[68-69]。一株完整的树可以分为心材、边材、树皮三大部分。树皮能够保护营养丰富的组织免受动物的侵害[61,70],因此在分解早期,树皮未脱落时,能够作为环境过滤器影响相关的分解者对木材进行分解[68,71]。粗糙的树皮能够使得无脊椎动物更好附着,影响动物的聚集[72]。同样,Jones等[73]研究也发现树皮的存在减缓了木材的分解。在分解后期,木材内部的特性变得更加重要,部分原因是大部分树皮已经脱落或者树皮特性退化。Zuo等[74]比较了11种温带树种的树皮特征及其对无脊椎动物丰富度和群落组成的影响,结果表明树种间树皮性状的差异性越高,无脊椎动物的丰富度越高。这表明在早期分解过程中树皮性状的差异比树种多样性作为影响分解指标更好。除了树皮的影响外,心材和边材的理化性质差异也会导致分解速率的不同[75]。心材通常比边材坚硬,木质素含量更高,更耐腐蚀。心材和边材之间的氧气可利用性、含水量和pH都可能存在显著的差异。Moll等[76]研究发现,不同树种边材和心材的微生物群落结构和微生物群落聚类(operational taxonomic units,OTU)丰富度存在显著的差异,边材OTU的丰富度高于心材。另外,大多数木本植物的心材分解还受到各种脂肪酸和醇类的抑制,渗透性和含水率低。心材中的单宁和萜类提取物的含量通常高于边材,并且心材提取物中通常含有较高的毒性物质,这也是导致心材的分解通常比边材慢的原因[77]。木材自身的特性很大程度影响到分解的过程。不同的木质部位分解的速度也不尽相同,这主要取决于不同部位木材的木质素、萜类、酚类等化合物的含量,含量越高的木材分解越缓慢。
3 展望
森林中的CWD代表了陆地生态系统的重要C储存之一,在全球范围内发挥着关键的生态作用,确定枯木分解对于理解全球C循环和维持生物多样性至关重要。已有关于CWD分解研究极大地促进了对CWD分解过程的理解,并为其管理和保护提供了关键信息。影响CWD分解有诸多因素,如气候、各种生物类群、木材本身特性等,以及其之间复杂的相互作用。在全球尺度上,气候因子是影响分解的主要原因,而在局域尺度上则取决于分解者群落和木材本身特性。土壤动物通常在分解早期通过破碎CWD来促进分解,而微生物群落变化在CWD分解的全过程中产生积极(或消极)影响。此外,木材自身的特性很大程度影响分解的过程,木质素、萜类、酚类等化合物含量越高的木材分解越缓慢。
由于CWD具有结构复杂、分解缓慢等特点,影响分解的因素之间具有复杂的相互作用,研究方法存在诸多的不足,使得CWD分级研究仍然面临许多困难。为了进一步明确CWD在生态系统中的作用,未来的研究重点应关注:CWD分解过程中各因素之间的相互作用;CWD的分解是一个缓慢的C释放过程,除了需要对其进行长期的定位研究,区分位于不同的分解阶段的CWD的转化过程以及养分释放对生态系统循环的影响外,还应将地上地下的生态过程联系起来,探究C输入和C输出之间的平衡;随着宏基因组和分子生物学的飞速发展,这些新兴的技术手段打破了原始研究方法的局限性。通过宏基因组学的方法关注与分解过程息息相关的重要的腐生生物类群以及类群之间的相互作用,应成为今后研究的重点。
