邵武将石自然保护区花榈木种群结构特征研究
2024-01-16杨开兴
杨开兴
(邵武将石省级自然保护区管理处,福建 邵武 354000)
花榈木(Ormosia henryiPrain)为豆科(Leguminosae)红豆属(Ormosia)常绿乔木,生于山坡、溪谷两旁杂木林内[1],目前气候下核心分布区主要为江西、湖南、福建、浙江等省份及周边区域[2]。花榈木具有用来制作高档家具、入药和园林绿化的价值,由于人为过度砍伐和自身种子休眠期长、不易发芽、生长速度慢等原因[3]导致其数量稀少,被列为国家二级保护植物[4]。目前,国外关于花榈木的研究报道较少,国内的研究主要集中在花榈木种子及幼苗方面[3],而在种群生态学和保护策略方面的研究较为少见,花榈木仍缺乏较完善的保护策略。本文以邵武将石自然保护区内野生花榈木资源为研究对象,通过调查保护区内花榈木的种质资源,从资源数量、种群结构特征等方面进行系统、全面的研究,分析花榈木的致危原因,为进一步保护和开发利用其野生资源提供理论依据。
1 研究方法
1.1 研究区域概况
将石自然保护区位于我国东部的福建省南平市邵武市肖家坊镇境内,东经117°14′11′′ ~117°16′40′′,北纬27°02′03′′ ~ 27°05′21′′,属武夷山脉向南延伸部分,总面积约1 200 hm2。境内为温暖湿润的中亚热带季风性气候,年均气温17.7℃,最冷月(1月)平均气温6.8°C,最热月(7月)平均气温27.5°C,全年日照时数1 763.2 h。年平均降雨量1 846.6 mm。植被为中亚热带常绿阔叶林,原始森林已几经破坏,现多以壳斗科植物如甜槠(Castanopsis eyrei)、苦槠(Castanopsis sclerophylla)等为建群种的次生常绿阔叶林和以马尾松林为代表的暖性针叶林[5]。
1.2 野外调查方法
于2022年7月进行野外调查,在将石自然保护区内,沿已有道路、登山路和野路等共16条样线进行调查,共统计花榈木个体160株。对调查到的花榈木进行每木检尺,测量植株的胸(地)径、树高和冠幅,同时记录花榈木的生长状况、受威胁程度、人为干扰程度和周围的共生植物,整理形成将石保护区内的花榈木信息卡。
1.3 数据分析
参考戈峰[6]、李博[7]等对种群动态的分析方法,对调查数据进行分析整理。利用径级替代龄级的方法,将花榈木分为10个龄级,龄级宽度为2 cm,编制出静态生命表[8]、存活曲线和生存分析曲线。
生存分析曲线包含了种群生存率函数S(n)、累计死亡率函数F(n)、死亡密度函数f(n)和危险率函数λ(n)。公式分别如下:
其中:n为龄级;Pn为存活率;An为龄级宽度(此处为径级宽度,单位为cm),根据花榈木种群的生物学特性,龄级宽度划分标准以2 cm为一龄级。
参考以往研究[9]对于幼苗的界定,I龄级为幼龄,II、III龄级为中龄,IV龄级及以上为成年。根据种群动态数量量化方法[10],定量比较、评价种群数量动态。选用一次移动平均法[11-12],对未来 2、3、4个龄级时间后花榈木种群龄级结构进行模拟和预测。种群内两相邻龄级间(n到n+ 1龄级)个体数量变化动态指数(Vn)、整个种群的年龄结构的数量变化动态指数(Vpn)及考虑未来外部干扰时种群年龄结构动态指数(Vpn′),计算公式如下:
其中,n表示龄级;an为第n龄级的个体数;k为种群年龄级数量。
其中,t表示所要预测的时间年限表示在未来t年后,n年龄级的种群大小;Xk表示k龄级当前的种群大小,利用现有的k龄级预测n年龄级t年后的种群龄级结构。
2 结果与分析
2.1 种群静态生命表及存活曲线
从花榈木种群的静态生命表(表1)可以看出,生命期望大体呈现,“低-高-低”的变化。种群的生命期望值(en)最高出现在第IV龄级,为14.00,在调查中,所统计的第IV龄级为空白,而其en又为最高,第IV龄级前的龄级的en分别为1.32、1.74、3.33,同时ln急剧下降,这说明第IV龄级前的龄级死亡率较高,受环境影响反应较剧烈,推测在无胁迫下花榈木种群中第IV龄级的个体数量应不在少数。因此,若要恢复花榈木种群数量应从第IV龄级着手。

表1 花榈木种群静态生命表Tab.1 Static life table of O. henryi population
存活曲线的走势反映了生存率和死亡率随年龄变化的趋势[6]。图1存活曲线显示,保护区内的花榈木种群与标准的Deevey-I、Deevey-II和Deevey-III型存活曲线有较大的差别[6]。由于种群在IV和VII龄级个体的缺失,致使曲线表现出不规则 “M”型,这与Deevey-II型的b1变型[7]近似,即在种群发育的多个时期呈现出波峰波谷式的交替变化,个体存活率之间存在十分明显的差异。部分物种的生态位重叠,比如光照致使种间竞争加剧,有20株遭受过度荫蔽的威胁,达到调查总数的12.5%。并且均属于第I龄级,可能是第I龄高死亡率[13]的原因之一。由于第IV、VIII龄级的缺失,样方内花榈木种群的死亡率(qn)和消失率(Kn)曲线都不断在正负值之间变化,通过存活曲线的波折起伏,可以看出花榈木种群的不稳定以及种群的脆弱性,但花榈木种群在成年之后种群较为稳定。
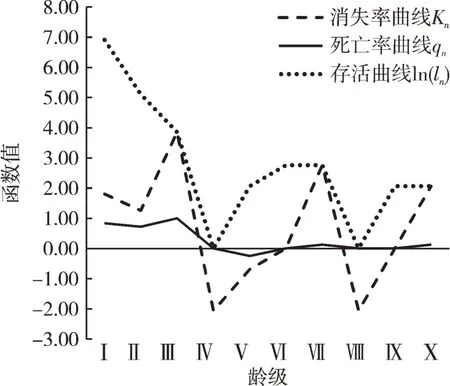
图1 花榈木种群的存活曲线ln(ln)、死亡率曲线qn、消失率曲线KnFig.1 Survival curve ln(ln)、 mortality rate curve qn and killing power curve Kn of the O. henryi population
2.2 种群生存分析
花榈木种群的4个生存函数曲线见图2。从图中可以看出,生存率曲线(Sn)从第I龄级开始就急剧下降,到III龄级基本降为0,并且接下来一直保持在0的水平。而累计死亡率曲线(Fn)则与Sn完全相反,形成对照,相互补充。这一现象的主要原因可能是样方中的多数幼苗在此期间死亡,造成中龄及成年树补充不足。死亡密度函数(fn)和危险率函数(λn)的走向分别与生存率曲线(Sn)和累计死亡密度函数(Fn)的走向基本一致。结合死亡密度函数(fn)和危险率函数(λn),都在II龄级处斜率产生较大变化,死亡速率下降,危险速率也下降,表明此时密度制约效应达到最大效果。4个生存函数曲线表明,花榈木种群前期幼苗数量多,中、后期个体数量过少,种群结构属于弱增长型。这与前面对花榈木种群存活曲线等的分析结果是相同的。
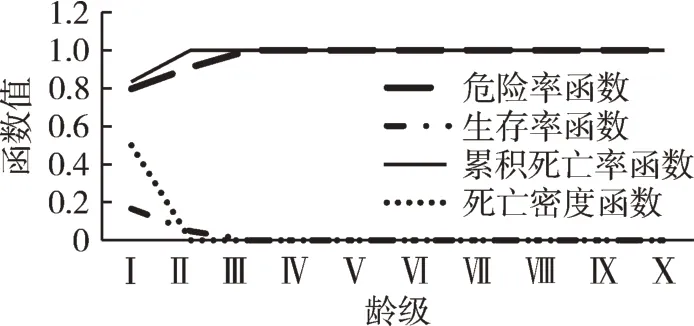
图2 花榈木种群生存函数曲线Fig.2 Survival function curves of O. henryi population
2.3 种群数量动态分析
对花榈木种群相邻龄级间个体进行数量动态量化指数分析得表2。由于部分的龄级个体数为0,数据缺失,VI龄级的Vn为0。由于Vn(Vpn)正、负和零分别表示种群的增长、衰退和稳定[10],因此各龄级间存在明显的波动现象,说明花榈木种群由幼树向中树的过渡过程表现出明显的衰退趋势。当考虑种群无未来外部环境随机干扰时,花榈木种群的Vpn均为正值,该区种群的多个龄级阶段的Vn< 0。推测出花榈木种群结构由衰退向增长过渡,表现出一种缓慢增长的状态。由于部分龄级缺失,个体数为0,在计算存在外部干扰时的数量变化动态指数Vpn′时没有除以最小龄级个体数min(a1,a2,a3,…ak)(等于0)。故Vpn与Vpn′之间相差为10倍(龄级数量k)。所以当考虑种群有未来外部环境随机干扰时,Vpn′均大于 0,且均远小于Vpn值,表明花榈木种群较为敏感脆弱,受环境影响较大,抵抗力稳定性较低。

表2 花榈木种群两相邻年龄级个体数量变化的动态量值及种群年龄结构的动态指数Tab.2 Dynamic values of the number of individuals in two adjacent age classes and dynamic indices of the population age structure of O. henryi population
2.4 种群时间序列预测
利用时间预测模型,将结果绘成龄级与个体数的关系图(图3)。花榈木种群在推迟 2 个龄级时间以后,II、III、IV、V龄级个体数量均有所增加,而后与现种群数量M0基本相同,成年个体数基本为0。推迟 3、4 个龄级时间后,趋势基本相同。其中移动后的第一个龄级都有着最大的增幅,依次为350.00%、850.00%、383.00%。由此可以推测,若幼龄植株可以顺利发展为中龄、成年植株,补充高龄级的个体,则如上述结果所示种群数量可以获得大幅度增加。结合上面的分析结果,表明花榈木种群中幼龄个体数量充足,种群结构为弱增长型。我们可以初步推测,按照目前发展趋势,再经过4个龄级时间以后,花榈木种群数量虽然可以得到提升,但与正常成年种群相比仍有一定差距。同时如果不采取合理就地保护措施,VII龄级之后的个体数仍然处于极低的水平,成年个体过少,种群数量仍无大幅度提升,种群结构可能由弱增长型转为衰退型。

图3 花榈木种群数量动态的时间序列预测Fig.3 Time sequence prediction of O. henryi population dynamics
3 结论与讨论
将石自然保护区内花榈木种群分散,幼龄级个体数量丰富,而中龄、成年树个体数量较少,种群结构呈弱增长型。种群虽具备自然更新的潜能,但生命表显示幼龄树存活率低,存活曲线与Deevey-II型的b1变型[7]近似,种群前期不稳定且较为脆弱。部分龄级缺失,成年树个体数稀少,种群增长力低,发展受阻,这与猪血木(Euryodendron excelsum)[11]与元宝冷杉(Abies yuanbaoensis)[9]相似。由于成年个体过少,种群数量无法恢复,种群结构可能由弱增长型转为衰退型。
种群年龄结构、生命表和数量动态分析可以了解一个种群的现存状态,是分析种群过去、现在与未来的发展动向的重要工具之一[14-15]。对以上结果,主要原因有以下几点:(1)花榈木种子不易萌发,幼苗存活率低,生长相对缓慢,易受病虫害危害。种子萌发后的生态环境不适宜生存,导致了花榈木种群在I龄级后的高死亡率,这是花榈木种群难以发展的根本原因。(2)花榈木所在群落植物生长茂盛,优势树种影响强烈,这使得萌发出的花榈木幼苗在其进一步成长为中龄树的过程中,由于对光照需求增加而环境郁闭度高的矛盾中遭受了较大的阻力,在群落中易成为种间竞争中的牺牲品。(3)花榈木有部分遭到劈砍,保护区内农户的保护意识不够强烈,导致种群内成年个体更加稀缺,甚至部分年龄级个体缺失。以上原因使得花榈木种群内成年个体无法得到正常补充,个体数量极少,种群发展、自然更新受阻。同时从种群动态预测结果看,种群的波动性也表明花榈木种群有恢复为正常种群的潜能,只要受到适宜的保护,其种群数量有可能逐步恢复[16]。但时间序列预测分析结果显示,如按目前趋势发展下去,可能导致花榈木的种群由弱增长转为衰退型。因此,若要恢复花榈木种群数量,首先,对保护区内的花榈木种群中的各龄级分别采取保护措施,尤其是成年个体的保护;其次,对民众进行科普教育,严格控制人为砍伐及保护区生态旅游开发等其它方式的破坏,为花榈木种群发育提供适宜的环境;最后,在花榈木生长特性、种子休眠、人工育种等方面开展系统的技术研究,解决种苗和自然回归问题。综上,恢复花榈木种群数量,保证花榈木种群自然更新是目前的首要任务,是遵循基于自然的解决方案的有效途径[17],这对花榈木自然种群的保护和发展有着极重要的意义。
