基于水稻大粒染色体片段代换系Z29的鉴定及QTL定位
2024-01-15刘金艳张朔语宗涵颖陈文博韦秘吴如会母建妍张记超凌英华张长伟何光华赵芳明张婷
刘金艳,张朔语,宗涵颖,陈文博,韦秘,吴如会, 母建妍,张记超,凌英华,张长伟,何光华,赵芳明,张婷
西南大学 农学与生物科技学院/南方山地农业教育部工程中心,重庆 400715
水稻作为重要的粮食作物,其产量三要素包括穗粒数、有效穗数和千粒质量,其中千粒质量主要受水稻粒型的影响,水稻粒型又由粒长、粒宽和粒厚决定.这些因素中的任意一个发生改变,都会对籽粒的大小产生影响,从而影响水稻的产量和品质.克隆粒型发育相关基因并解析其分子机制,有利于将其应用于水稻的高产育种工作中.根据GRAMENE网站(http: /www.gramene.org/archive)提供的数据,在水稻12条染色体上已有超过600个与粒型相关的QTL被检测到,还有部分QTL被精细定位和克隆.根据这些QTL所参与的代谢或信号途径,可以分为以下几类: G蛋白信号通路、植物激素信号通路、泛素-蛋白酶体通路、转录调控通路、丝裂原活化蛋白激酶信号通路和表观遗传调控通路[1].
在G蛋白信号通路中,RGA1/D1编码G蛋白的α亚基,突变体d1中G蛋白失活导致细胞分裂受到抑制,从而使籽粒变小[2];RGB1编码Gβ亚基,对细胞增殖起正向调控作用,基因突变导致植株变矮,籽粒变小[3];GS3编码1个水稻Gγ亚基,参与水稻Gβγ二聚体形成,是一个控制水稻粒长和粒质量的主效QTL,对粒长起负调控作用[4].在植物激素信号通路中,生长素原初响应基因BG1参与生长素的运输与转运,功能获得性突变体生长素含量增加、颖壳细胞的数量增多,种子变大,粒长和粒宽均显著增加[5];TGW6编码1个IAA-葡萄糖水解酶,是一个粒长负调控因子[6];SLG编码1个BAHD家族酰基转移酶蛋白,通过促进水稻中内源BR的合成,从而调控颖壳细胞的大小,进而控制粒长的发育[7].泛素-蛋白酶体通路中,GW2编码1个E3泛素连接酶,GW2会影响泛素分子与目标蛋白的连接,使得原本应该被降解的目的蛋白不能被蛋白酶体特异识别,进而激活颖壳外表皮细胞的分裂,增加颖壳的宽度,是一个粒宽负调节因子[8];WG1编码1个谷氧还蛋白,通过促进细胞增殖来控制粒宽,WG1功能缺失突变体的颖壳横向细胞数显著降低[9];WTG1基因编码1个otubain-like蛋白酶,是一个去泛素化酶,WTG1功能缺失突变体的颖壳细胞增大,表现出粒宽变宽、粒厚变厚以及每穗粒数增加的表型[10].转录调控通路中,GLW7编码植物特异性转录因子OsSPL13,这是一个能控制籽粒长度和宽度的主效QTL,通过正向调控颖壳细胞大小,从而提高籽粒长度和产量[11];GL8编码转录因子OsSPL16,通过促进细胞分裂及灌浆速率来增加籽粒宽度[12].丝裂原活化蛋白激酶信号通路中,SMG1编码1个丝裂原活化蛋白激酶OsMKK4,基因突变后会使水稻籽粒变小[13];DSG1编码1个丝裂原活化蛋白激酶OsMAPK6,dsg1突变体粒型变小、植株矮化[14].表观遗传调控通路中,OsmiR396调节GS2编码的生长调控因子OsGRF4,GS2突变会影响OsmiR396的调控,从而提高水稻的粒长和质量[15];SDG752编码甲基化转移酶H3K36,它的表达下降会导致种子变小[16];GW6a编码具有组蛋白乙酰转移酶活性的类GNAT蛋白OsgIHAT1,其通过控制细胞分裂来调控粒长[17].
水稻粒型的遗传机理十分复杂,目前克隆的基因还不足以揭示粒型的遗传多样性和分子机理.水稻染色体片段代换系可以提供丰富的自然变异,且仅存在少量代换片段的差异,有清晰的遗传背景,因此是复杂性状研究的理想材料.本研究以日本晴为受体亲本、恢复系R225为供体亲本,鉴定到1个携带10个代换片段的水稻染色体片段代换系Z29(CSSL-Z29).通过对水稻染色体片段代换系Z29进行农艺性状的测量与细胞学分析,探究籽粒变化的原因,再利用构建的次级F2群体进行粒型相关QTL定位,进一步结合MAS法在F3代中选育了14个次级代换系片段.研究结果对于目标QTL的分子机制研究和分子设计育种有重要的意义.
1 材料与方法
1.1 材料
水稻CSSL-Z29是以测序粳稻品种日本晴为受体亲本、西南大学自育的优良籼稻恢复系R225为供体亲本,经过高代回交以及自交,结合全基因组SSR分子标记辅助选择选育而成的10片段优良代换系.QTL定位群体材料由日本晴和Z29杂交构建的由124个单株组成的次级F2分离群体.
1.2 水稻大粒CSSL-Z29培育及代换片段鉴定
利用在水稻基因组中均匀分布的429个SSR标记,对日本晴和R225进行多态性分析,筛选出253个多态性标记[18]; 然后将这些多态性标记从BC2F1代开始进行分子辅助选择,从中选择20株进行自交; 最后每个株系的每代取10株继续进行分子标记辅助选择[19],在日本晴/BC2F4自交的F7代选育出含10个代换片段的水稻长粒染色体片段代换系Z29.水稻染色体代换片段的鉴定参考Zhou等[20]的方法.当某个标记的带型与受体亲本相同时,表明该片段的DNA来自于日本晴; 当某个标记的带型与供体亲本相同时,表明该片段的DNA来自于R225.连续的供体亲本带型所在区段表示为代换片段,代换片段长度的计算参照Paterson等[21]的方法.
1.3 材料种植方法
2019年将日本晴与Z29杂交产生F1代,收获F1代种子并在夏末季于海南种植,收获F2代种子.2020年3月,将日本晴、Z29以及F2群体的种子在西南大学水稻研究所歇马试验基地播种.4月中旬,首先规划试验田(株距16.67cm,26.67cm),再将30株日本晴、30株Z29和124株F2单株移栽至试验田并进行常规处理.2021年4月,在西南大学水稻研究所歇马试验基地以同样的规格和管理方式种植F2代,从中选出目标株系及其受体亲本进行次级代换片段的选育.
1.4 表型分析和农艺性状统计
在水稻成熟后,试验基地取10株日本晴、10株Z29和124株F2群体,统计每株的株高、有效穗数、穗长、一次枝梗数、二次枝梗数、粒长、粒宽、籽粒长宽比、每穗实粒数、每穗总粒数、千粒质量、结实率和单株产量共13个性状.粒长和粒宽使用20 cm直尺测量连续排列整齐的10粒种子的总长和总宽,重复3次,然后取平均值; 结实率为每穗实粒数占每穗总粒数的百分比; 长宽比为粒长和粒宽的比.最后使用Microsoft Excel 2019统计10株日本晴和10株Z29及F2群体13个性状的最大值、最小值、平均值、标准差、偏度和峰度,并进行t检验.
1.5 QTL定位
采用CTAB法提取DNA[22],然后进行PCR扩增,聚丙烯酰胺凝胶电泳和银染法显色,日本晴带型、Z29带型、杂合带型、缺失带型分别用“-1”,“1”,“0”,“.”来表示基因型的赋予值.根据F2代124个单株对应的指定值,在SAS统计软上使用HPMIXED程序的限制最大似然法来进行QTL定位[23].
1.6 次级代换片段系选育
根据QTL定位结果,在日本晴和Z29的F2群体中选择有目标QTL的单株进行种植,发展成F3群体(每个株系种植20株),利用MAS法进一步筛选出含少量或不含杂合标记的次级单片段或多片段代换系.
1.7 扫描电镜
取抽穗期同一部位的日本晴和Z29水稻颖壳,Hitachi SU3500扫描电子显微镜和-40 ℃冷却台进行新鲜样品的观察拍照.
1.8 石蜡切片
取抽穗期同一部位的日本晴和Z29水稻颖壳,在FAA溶液(50%乙醇,0.9 mol/L冰醋酸和3.7%甲醛)中抽真空,50 ℃条件下固定过夜; 然后将它们在一系列乙醇溶液中脱水,用二甲苯取代,并包埋在石蜡中; 对样品进行切片,转移到聚赖氨酸涂层的玻璃载玻片上,用二甲苯脱脂,并通过一系列的乙醇溶液脱水; 用1%的番红和1%的固绿对切片进行染色,通过一系列的乙醇溶液进行脱水,用二甲苯浸润,并安装在盖玻片下面; 用二甲苯对载玻片上的样品进行脱蜡,依次用0.1%番红和0.5%固绿对样品进行染色,用二甲苯浸润封片,并用Nikon Eclipse E600显微镜进行光镜观察.
2 结果与分析
2.1 Z29代换片段的鉴定
以粳稻测序品种日本晴作为受体,自育优良籼稻恢复系R225作为供体,选育出1个10片段代换系(图1),代换片段分布于第3,6,7,10和11号染色体上,总代换长度为28.95 Mb,最长代换片段为8.01 Mb,最短代换片段长度为1.18 Mb,平均代换长度为2.90 Mb.

每个染色体的右侧为代换片段长度和分子标记,左侧为代换片段鉴定出的QTL和标记的物理距离,黑色部分代表片段.GL为粒长,GW为粒宽,RLW为长宽比,NSB为二次枝梗数,PH为株高,PL为穗长.图1 Z29的代换片段
2.2 表型鉴定
Z29与受体亲本日本晴(Ni)相比株型差异不明显(图2a),但粒长极显著增加了38.83%(图2d和2e),粒宽极显著增加了11.52%(图2c和2f),千粒质量极显著增加了24.01%(图2g),每穗总粒数极显著增加了22.54%(图2h),每穗实粒数极显著增加了21.66%(图2i),穗长极显著增加了16.58%(图2j),一次枝梗数差异无统计学意义(图2k),但二次枝梗数极显著增加了32.21%(图2l).此外,株高极显著增加了11.88%(图2m),有效穗数极显著降低了32.21%(图2o),结实率和单株产量差异无统计学意义(图2n和2p).大粒、大穗是Z29最主要的特征,说明Z29是一个在水稻育种中具有重要价值的品种材料.
2.3 细胞学分析
为探究Z29籽粒变大的原因,利用扫描电镜观察Z29与受体亲本日本晴抽穗期颖壳中间部位内外表面的细胞形态(图3),利用ImageJ软件测量内表皮细胞的长度、宽度以及颖壳外表皮单位面积细胞数.统计结果发现,与日本晴相比,Z29在颖壳外表皮纵向细胞数极显著增加了31.48%(图3a和3i),横向细胞数极显著增加了46.07%(图3a和3j); Z29颖壳外表皮单位面积细胞数极显著增加了62.82%(图3b和3h); Z29颖壳内表皮细胞长度极显著增加了22.45%(图3c和3f),颖壳内表皮细胞宽度显著增加了0.05%(图3c和3g),因此,Z29籽粒变大主要是由于颖壳细胞的增殖和伸长共同引起的.
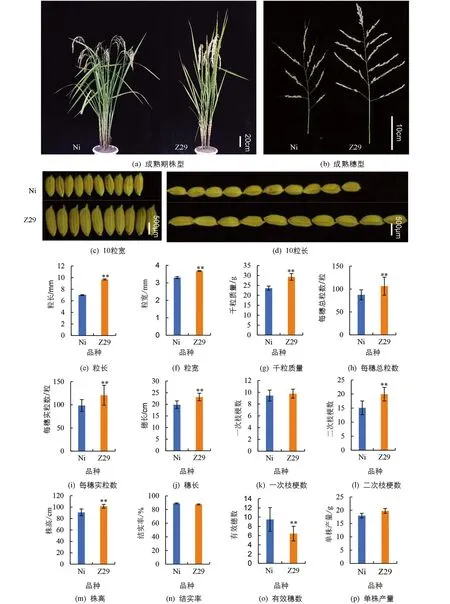
*表示p<0.05,**表示p<0.01,差异有统计学意义.图2 Z29和日本晴(Ni)的表型分析
石蜡切片分析结果发现,与日本晴相比,Z29颖壳的薄壁组织细胞长度显著增加了8.36%(图3d,3e和3k),但宽度极显著降低了10.08%(图3d和3l); 同时发现Z29颖壳的薄壁组织的细胞数极显著增加了8.92%(图3d,3e和3m),进一步从细胞学的层面上证实了Z29细胞增殖和伸长能共同促进Z29籽粒增大.

*表示p<0.05,**表示p<0.01,差异有统计学意义.图3 Z29和日本晴(Ni)的细胞学分析
2.4 粒型性状在日本晴/Z29的次级F2群体中的频率分布
随机取F1代125颗籽粒进行形态分析,Z29的长粒相对短粒呈显性遗传.根据在日本晴/Z29的次级F2群体中的频率分布可知,其粒长(图4a)、粒宽 (图4b)、长宽比(图4c)以及千粒质量(图4d)的分布均呈现出非标准正态分布,表明这些性状由多基因控制,可进行QTL分析.

图4 粒型性状在日本晴/Z29构建的次级F2群体中的分布频率
2.5 基于日本晴/Z29的次级F2群体的粒型QTL鉴定
在次级F2分离群体中共鉴定出8个粒型QTL,分别为2个粒长QTL(qGL-3,qGL-10),1个粒宽QTL(qGW-3),4个长宽比QTL(qRLW-3,qRLW-6-1,qRLW-6-2,qRLW-10)和1个千粒质量QTL(qGWT-7),这些QTL分布在第3,6,7,10号染色体的相应代换片段上(表1和图1).qGL-3和qGL-10与Z29的粒长性状连锁,分别分布于3号、10号染色体的代换片段上,其中qGL-3的表型贡献率为13.28%,使粒长增加0.20 mm,表现为主效QTL(贡献率大于10%).qGL-10表型贡献率为9.43%,使粒长增加了0.17 mm,表现为微效QTL.qGW-3与Z29的粒宽性状连锁,对粒宽变异的贡献率为10.17%,使粒宽减少0.06 mm,表现为主效QTL.影响谷粒长宽比的QTL为qRLW-3,qRLW-6-1,qRLW-6-2和qRLW-10,对表型的贡献率依次28.49%,5.64%,7.92%和8.89%,其中qRLW-3和qRLW-10增加长宽比,加性效应分别为0.12和0.06,而qRLW-6-1,qRLW-6-2分别减小长宽比0.05和0.06.qGWT-7与Z29的千粒质量性状连锁,对表型变异的贡献率为5.88%,使千粒质量增加0.61 g.此外,粒长和千粒质量QTL均表现出增加性状值的正向加性效应,4个长宽比QTL中2个增加、2个减少,因而Z29的长粒主要由粒长和长宽比增加所致.长粒由1个主效QTL(qGL-3)和1个微效QTL(qGL-10)共同控制.qGL-3,qGW-3,qRLW-3共同连锁于分子标记RM6266,可能属于一因多效或QTL紧密连锁.

表1 Z29所鉴定出的粒型QTL(2021年)
2.6 次级染色体片段代换系的选育
在QTL定位的基础上,结合MAS法,在F3代中选育了14个次级片段代换系(表2),包括单片段代换系4个(S1-S4)、双片段代换系5个(D1-D5)、三片段代换系2个(T1-T2)和四片段代换系3个(FS1-FS3).为进一步研究数量性状遗传机理提供了理想材料.

表2 F3选育出的次级染色体片段代换系

续表2
3 讨论
本研究鉴定了1个以日本晴为受体亲本、恢复系R225为供体亲本的大粒染色体片段代换系Z29,携带10个来自供体亲本R225的代换片段,平均代换长度2.90 Mb.其粒长、粒宽和千粒质量均极显著增加,同时具有株高、穗长也显著增加等表型.组织学分析表明Z29籽粒变大与细胞增殖和细胞伸长有关; 粒型相关农艺性状在F2群体中呈非标准正态分布,表明这些性状均由多基因控制.
水稻产量由千粒质量、穗粒数和有效穗数决定,千粒质量主要受粒型的影响,粒型由粒长、粒宽和厚度决定.我们以日本晴与Z29杂交后构建的F2群体作为QTL定位群体,共鉴定到8个粒型相关的QTL,分布在第3,6,7,10号染色体上,包括2个粒长QTL,1个粒宽QTL,4个长宽比QTL,1个千粒质量QTL,这些聚集在同一标记上的QTL可能属于一因多效或紧密连锁.在已报道的研究中,OsSGL是1个一因多效的关键基因,能调节细胞的轴向性生长,最终使水稻的籽粒变大并且产量增加,过量表达OsSGL基因能显著增加粒长和粒质量.细胞学观察发现,颖壳细胞数目的增加、细胞体积纵向增大是粒长增加的主要原因[24].SLG7编码与LONGIFOLIA 1和LONGIFOLIA 2同源的蛋白质,通过增加纵向细胞长度、减小细胞横向宽度,产生细长籽粒[25].GS5编码1个丝氨酸羧肽酶,其表达量的升高会促进水稻颖壳的细胞分裂,使粒宽增加[26].GL7编码拟南芥LONGIFOLIA蛋白的同源蛋白,而LONGIFOLIA能调节细胞的纵向伸长,GL7表达水平上调会增加水稻粒长并改善稻米外观品质[11].这些基因均通过调控细胞增殖与细胞大小来影响籽粒的大小.与之相比,Z29同样通过影响颖壳细胞的数目与大小来影响籽粒大小,Z29籽粒增大是由于颖壳细胞增殖和伸长共同引起.这些研究结果为粒型相关QTL的进一步分解和功能分析奠定了基础,也为水稻分子设计育种提供了理论依据.
水稻籽粒增大的原因可能是由于颖壳细胞增殖、细胞扩张或增殖与扩增共同作用所致.GL3.1编码1个丝氨酸/苏氨酸磷酸酶,该激酶对水稻粒长起负调控作用,并且GL3.1能直接去磷酸化底物T1; 3(细胞周期蛋白),而T1; 3表达下调会导致籽粒变短[27].GW6a编码1个拥有组蛋白乙酰转移酶的类GNAT蛋白OsglHAT1,GW6a表达上调,通过增加细胞数目使籽粒增大[17].GW5在种子的发育过程中参与泛素化蛋白酶体的降解过程,该基因功能缺失后,不能正常参与蛋白质泛素化降解途径,从而促进水稻颖壳的细胞分裂过程,导致颖壳的细胞宽度增加,最终影响水稻的籽粒宽度[28].大多数基因都是通过调控水稻颖壳的细胞数目来调节水稻籽粒大小的,而GLW7缺失突变以及过量表达对水稻颖壳的细胞数目并无明显影响,其主要是通过控制细胞的扩张从而影响籽粒大小[25].SMG11是DWARF2(D2)的新的等位基因,编码1个参与油菜素内酯BR生物合成途径的细胞色素P450家族成员;SMG11通过促进籽粒颖壳的细胞扩张来控制籽粒大小,在一定程度上影响了细胞扩张中涉及籽粒大小的相关基因的表达,从而调控细胞扩张;SMG11的适当表达增加了谷粒的大小、粒质量和产量[29].此外还有一些基因能够同时影响细胞的增殖和扩张,BG1编码1个膜定位蛋白,通过增加颖壳细胞增殖与扩张正向调控籽粒大小[14].OsmiR396-GS2/OsGRF4-OsGIFs调控途径通过调节细胞扩展和细胞增殖,进而对水稻的粒型和粒质量产生影响[15].本研究中,Z29籽粒的大小由颖壳细胞的大小和数量共同调控,这有利于深入研究水稻调控籽粒大小的分子机理,为挖掘提高水稻产量及品质的优良品种提供重要材料.
4 结论
本研究选育了1个以日本晴为受体、自育优良籼稻恢复系R225为供体亲本的水稻染色体片段代换系Z29.Z29含有来自R225的10个代换片段,平均代换长度2.90 Mb.农艺性状调查发现,相较于日本晴,Z29粒长极显著增加了38.83%,粒宽极显著增加11.52%,表现为大粒表型.扫描电镜观察和细胞学分析结果表明,Z29籽粒变大是由细胞数量极显著增多、增大引起.利用日本晴与Z29杂交构建的次级F2群体进行QTL定位,共检测到8个与粒型相关的QTL,包括2个粒长QTL(qGL-3,qGL-10),1个粒宽QTL(qGW-3),4个长宽比QTL(qRLW-3,qRLW-6-1,qRLW-6-2,qRLW-10)和1个千粒质量QTL(qGWT-7).进一步利用MAS法在F3群体中选育出14个次级染色体片段代换系,包括4个单片段代换系、5个双片段代换系、2个三片段代换系和3个四片段代换系.
