Mepiquat chloride priming confers the ability of cotton seed to tolerate salt by promoting ABA-operated GABA signaling control of the ascorbate–glutathione cycle
2024-01-13QIQianWANGNingRUANSijiaMUHAMMADNoorZHANGHenghengSHIJianbinDONGQiangXUQinghuaSONGMeizhenYANGentuZHANGXilingandWANGXiangru
QI Qian, WANG Ning, RUAN Sijia, MUHAMMAD Noor, ZHANG Hengheng, SHI Jianbin, DONG Qiang,XU Qinghua, SONG Meizhen,2*, YAN Gentu*, ZHANG Xiling* and WANG Xiangru,2*
Abstract Background Ensuring that seeds germinate and emerge normally is a prerequisite for cotton production, esp.in areas with salinized soil.Priming with mepiquat chloride (MC) can promote seed germination and root growth under salt stress, but its mechanism has not been fully elucidated.In this study, physiological and biochemical experiments revealed that MC-priming promotes the tolerance of cotton seeds to salt stress by increasing the ability of antioxidant enzymes related to the ascorbate–glutathione (AsA-GSH) cycle to scavenge reactive oxygen species (ROS).
Keywords Seed priming, Salinity stress, ABA signaling, γ-aminobutyric acid, ROS scavenging, Mepiquat chloride
Background
As a global environmental problem, soil salinization affects approximately 1 billion hectares of land worldwide and causes tremendous losses in agricultural production(Jesus et al.2015; Wang et al.2022).Soil affected by salinization have been reported in more than 100 countries throughout the world with most cases of salinization induced by irrigation (Rengasamy 2006).Soil salinization and alkalization occur by natural or man-made processes and are among the adverse environmental stresses that affect crops.Cotton (especiallyGossypium hirsutumL.) is the largest source of renewable textile fibers, making it an important cash crop (Wang et al.2022; Wu et al.2018).It is considered an ideal pioneer crop to improve salinized soil because it can generally grow in soil with a salt concentration of not more than 0.3%.However, compared with its later development stages, the germination and emergence stages of cotton seeds are more sensitive to salt stress, which hinders its production in saline soils(Wang et al.2019, 2021a and b).Therefore, it is essential to improve the salt tolerance of cotton seeds grown in saline environment.
Generally, different abiotic and biotic environmental fluctuations, including high salt stress, affect the seed germination (Wang et al.2019, 2021a and b).Since germination is facilitated by the energy stored in the seed(Zhou et al.2019), the inhibition of seed germination induced by salt stress may primarily occur due to energy changes in the metabolic activities induced by osmotic and ionic pressure (Wang et al.2021b; Li et al.2022).Salt stress inhibits β-oxidation and reduces the production of nicotinamide adenine dinucleotide (NADH) from the tricarboxylic acid (TCA) cycle, which is important for respiratory adenosine triphosphate (ATP) synthesis,thereby reducing energy production (Zhou et al.2019).Additionally, the electron transport chain may be excessively reduced under salt stress, thus, increasing the generation rate of reactive oxygen species (ROS) and altering the redox state of the cells (Blokhina 2003).Despite their toxicity, at low levels ROS as signaling molecules that control plant form and function has received growing attention in recent years (Demidchik et al.2018; Mittler et al.2022).However, excessive ROS accumulation could cause adverse effects in cells, such as lipid and protein oxidation, reduced enzymatic activities, DNA damage,and ultimately lead to oxidative damage (Blokhina 2003;Mittler et al.2022).Normally, the ROS scavenging pathway, also known as ascorbate-glutathione (AsA-GSH)pathway, is contingent upon the activity of four important antioxidant enzymes, viz., ascorbate peroxidase (APX),dehydroascorbate reductase (DHAR), glutathione peroxidase (GPX) and glutathione reductase (GR); and two antioxidant metabolites, ascorbate (AsA) and glutathione(GSH) (Kohli et al.2019).APX reduces the reactive H2O2to water while liberating malondialdehyde (MDA) which soon gets dismutated to yield AsA and dehydroascorbic acid (DHA).Later, in the presence of GSH, the DHA is reduced to generate AsA with simultaneous oxidation of GSH to oxidized glutathione (GSSG).Finally, the activity of GR regenerates GSH by transferring a proton from nicotinamide adenine dinucleotide phosphate (NADPH)(Kohli et al.2019).Thus, supplementing exogenous antioxidants or boosting the endogenous antioxidant defenses could combat the undesirable effects of the oxidative damage induced by ROS under saline conditions(Wang et al.2017, 2022).
Phytohormone signaling could also increase the tolerance of plants to salt stress (Zhao et al.2020).For example, abscisic acid (ABA), a key phytohormone, has been implicated in the reaction of plants to salt stress (Wang et al.2019; Zhou et al.2019).Previous studies have confirmed that exogenous ABA can mitigate the negative effects of hypoxia-NaCl stress during the germination stage (Wang et al.2021b, 2022;Yang et al.2016).When seeds are exposed to salt stress,ABA level increase dramatically due to the activated expression of ABA synthetic pathway genes, including zeaxanthin epoxidase, 9-cis-epoxycarotenoid dioxygenase (NCED),and abscisic aldehyde oxidase coding genes (Smirnoffand Wheele 2000).The accumulated ABA is then sensed by the pyrabactin resistance1/PYR1-like/regulatory component of the ABA receptors (PYR/PYL/RCAR), which represses the activities of protein phosphatase 2C isoforms (PP2Cs).This activity releases SNF1-related Kinase 2s (SnRK2s) to phosphorylate the downstream substrates, which enhances the tolerance of plants to salt stress (Danquah et al.2014).
In addition to ABA, γ- aminobutyric acid (GABA), a non-protein amino acid, plays a crucial role in the tolerance of plants exposed to various stresses (Bown and Shelp 2016).Many studies have reported the ability of GABA to function as a metabolite or signaling molecule in various physiological processes under stress conditions.For example, exogenous GABA can relieve the salt injury of seedlings by increasing the activities of superoxide dismutase (SOD), catalase (CAT), APX, and other antioxidants (Jin et al.2019; Wu et al.2021).Current research has clarified that GABA content is significantly increased by salt stress and replenishes the TCA cycle,thus, contributing to salt tolerance (Che-Othman et al.2020; Ji et al.2020; Wang et al.2022).Renault et al.(2013)found that inhibiting the synthesis of GABA reduced the salt tolerance of plants.Recent studies also confirmed that the synthesis of GABA is significantly activated during the germination stage in response to salt stress (Wang et al.2022; Che-Othman et al.2020; Wu et al.2021).
Glutamate decarboxylase (GAD) is the key enzyme of the GABA synthetic pathway (Ramesh et al.2017).Previous studies reported that salt stress rapidly activates the production of GABA through the transcriptional and post-translational regulation of GAD activity (Wang et al.2022; Che-Othman et al.2020).Moreover, GAD is a Ca2+-dependent calmodulin (CaM)-binding protein, and its activity can be modulated by Ca2+signaling (Renault et al.2013).Calcium signaling is produced in response to various abiotic stress stimuli in plants, such as salt stress, drought stress, and adverse temperature (Laohavisit et al.2009; Jiao et al.2022).Many studies have demonstrated that increases in internal Ca2+facilitate the binding of Ca2+to the calmodulin binding site on GAD in response to salt stress, thereby leading to variation in the levels of GABA (Ramesh et al.2017; Wu et al.2021).These studies further indicate that the synthesis of GABA is closely associated with Ca2+homeostasis during salt stress.
Mepiquat chloride (1,1-dimethylpiperidinium chloride, MC), an inhibitor of gibberellin biosynthesis, is one of the effective regulators that enhances plant tolerance to various stresses, including salt stress (Wang et al.2019).Our previous results indicated that MCpriming might maintain the redox homeostasis of cotton in response to salt stress during seed germination and seedling growth stages (Wang et al.2019, 2021a and b, 2022).Additionally, the contents of ABA and GABA were also increased by MC-priming under saline condition (Wang et al.2021b, 2022).Previous research suggested that ABA acts as a signal molecule and is engaged in the homeostasis and cross-talk between H2O2and GABA to mediate salt stress tolerance in plants (Yang et al.2016; Wang et al.2021a).Thus, we hypothesized that ABA and GABA might form a nexus and cross-talk strategy to regulate ROS homeostasis in plant cells.
Although there have been some studies on the potential interactions between ABA, GABA, and redox homeostasis in plant cells, it is still unclear how MCpriming triggers the antioxidant system of cotton seedlings to resist salt stress.To gain insights into the possible mechanisms involved in the tolerance of cotton to salt stress mediated by MC-priming, we evaluated the antioxidase and antioxidants defense system and the ABA and GABA homeostasis of the germinated cotton seedlings treated with MC-priming under salt stress.Our results demonstrate that ABA signaling mediated by MC-priming activates the synthesis of GABA.This stimulates the AsA-GSH cycle and prevents the accumulation of ROS in response to salt stress.The data obtained from the present study provides the mechanistic link between MC-induced elevation in the GABA level and adaptive responses of cotton seeds to salt stress, and deepen the understanding of the MC-mediated salt tolerance in cotton during seed germination.
Results
Effect of MC-priming on the characteristics of seed germination under salt stress
The germination rate, root length, and seed vigor index(VI) of the W-priming seeds under salt stress decreased significantly compared with those under normal conditions (Fig.1A-D).After four days of germination, MCpriming mitigated the effect of salt stress on growth inhibition, thereby significantly increased the germination rate, root length, and VI by 25.5%, 30.5%, and 67.3%, respectively, particularly in comparison with the W-primed (soaked in deionized water) seeds subjected to salt stress.Salt stress also enhanced the content of MDA in the W-primed seeds, which was significantly reduced by MC-priming (Fig.1E).Therefore, we used fluridone(an ABA synthesis inhibitor) or 3-mercaptopropionic acid (a GABA-synthesis inhibitor, 3-MP) to characterize the effects of ABA and GABA on seed germination parameters in response to salt stress.Under normal conditions, treatments with fluridone or 3-MP reduced the parameters of seed germination sharply but enhanced the content of MDA compared with the control that lacked an inhibitor (Fig.1).The inhibitor treatments further reduced the germination rate, root length, and VI under salt stress with no significant differences between the W-priming and MC-priming treatments.Fluridone + salt stress dramatically enhanced the MDA content, and the MC-primed treatments resulted in more pronounced effects.The MDA content of the W-primed treatment increased by 48.3%, while those of the MC-primed treatment increased by 1.1-fold compared with the salt stress treatment alone.The 3-MP + salt stress treatment also increased the content of MDA by 22.0% and 30.4% in the W- and MC-primed treatments, respectively, compared with the salt stress treatment alone.Thus, these results indicate that ABA and GABA could be partially necessary for the tolerance to salt stress induced by MC in the cotton seeds.
Effect of MC-priming on the accumulation of ROS under salt stress
Oxidative stress was determined by the generation of O2.-and H2O2in the germinated seeds in response to salt stress.The contents of O2.-and H2O2were localized in situ based on the varying intensity of blue (Fig.2A)and reddish (Fig.2B) coloration, respectively, suggesting that MC-priming minimized the accumulation of O2.-and H2O2under salt stress.Moreover, the determination results on the contents of O2.-and H2O2were consistent with those of histochemical staining.MCpriming significantly reduced the contents of O2.-and H2O2by 11.4% and 14.6%, respectively, under salt stress compared with W-priming (Fig.2C, D), which reduced the degree of cellular oxidative damage.The addition of fluridone and 3-MP to the treatments intensified the staining (Fig.2A, B), which indicated that the contents of O2.-and H2O2were enhanced.Thus, the treatments that contained inhibitors significantly increased the contents of O2.-and H2O2under the control and salt conditions compared with the treatment that lacked inhibitors(Fig.2C, D).In addition, more substantial effects were observed in MC-priming, and the fluridone inhibitors created a higher or non-significant difference in these traits compared with W-priming in the plants that had been subjected to salt stress.Meanwhile, 3-MP + salt treatment further increased the accumulation of O2.-and H2O2in cotton seeds treated with water and MC,increasing the O2.-content by 9.8% and 13.1%, as well as the H2O2content by 22.1% and 11.7%, respectively.Therefore, these results suggest that ABA and GABA might be important contributors to the redox state of germinating seeds that is maintained by MC.

Fig.1 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on the seed germination characteristics of the W- and MC-primed cotton seeds under salt stress.A Representative phenotypes of the W- and MC-primed cotton seeds after 24 h of salt stress.B Root length, C seed germination rate, D seed vigor index of the W- and MC-primed cotton seeds after 4 d of salt stress.E MDA content of the W- and MC-primed cotton seeds after 24 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS:water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.FWCK, FWS, FMCS:addition of fluridone based on WCK, WS and MCS.PWCK, PWS, PMCS: addition of 3-mercaptopropionic acid based on WCK, WS and MCS.The bars represent the mean ± standard error (SE) of three replicate samples
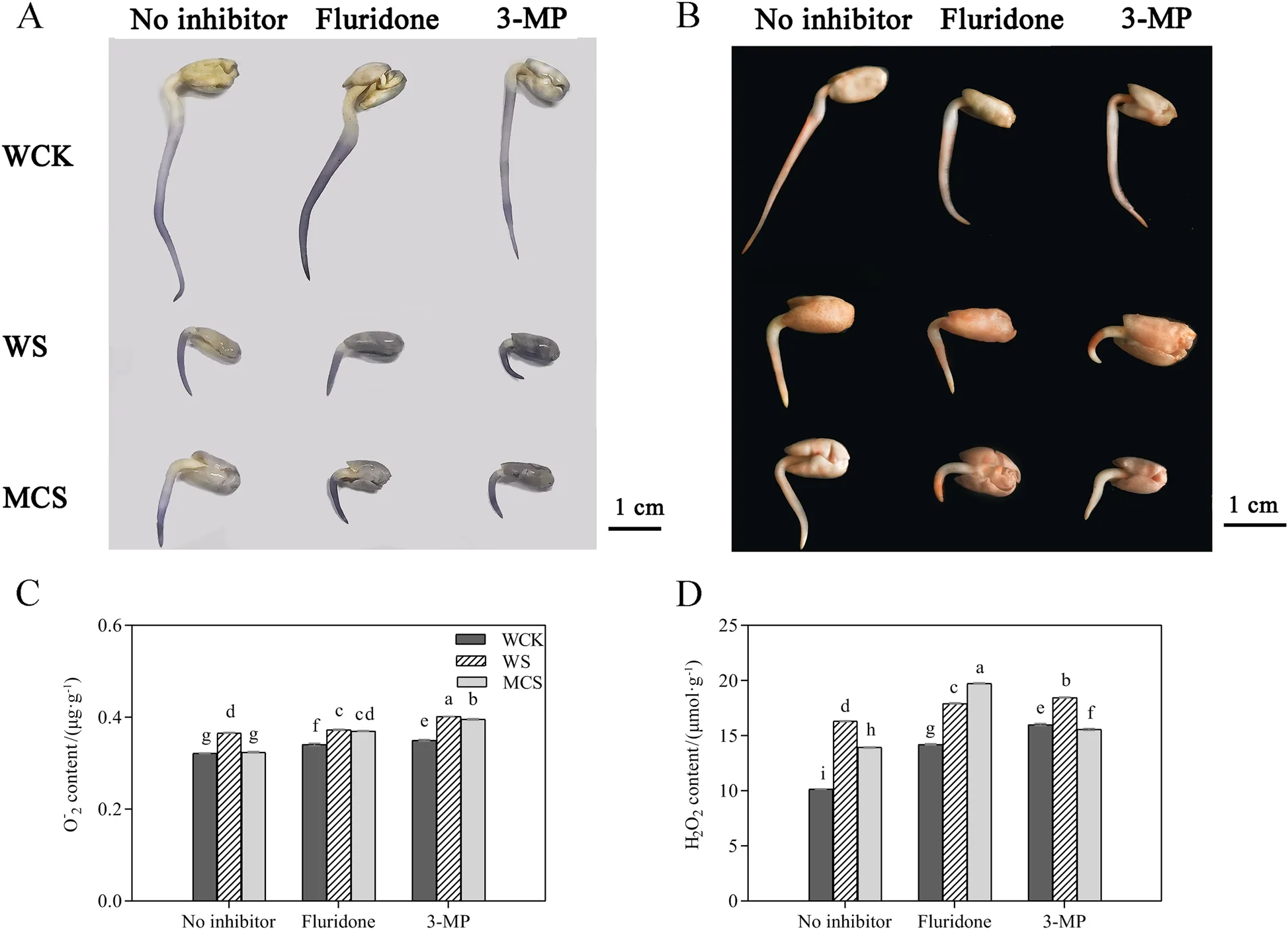
Fig.2 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on reactive oxygen species(ROS) homeostasis of the W- and MC-primed cotton seeds under salt stress.A Nitrotetrazolium blue chloride (NBT) and B 3,3’-Diaminobenzidine(DAB) histochemical staining of the W- and MC-primed cotton seeds after 24 h of salt stress.C Superoxide anion (O2.-) and D hydrogen peroxide(H2O2) contents in the W- and MC-primed cotton seeds after 24 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS:water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.The bars represent the means ± SE of three replicate samples
Effects of MC-priming on the activities of SOD, POD,and CAT under salt stress
Since antioxidant enzymes have crucial roles in mitigating cellular oxidation, we measured the activities of several antioxidant enzymes, including SOD, POD,and CAT.As shown in Fig.3, the activities of SOD,POD, and CAT and expression levels ofGhSODCC,GhPOD5, andGhCAT1were significantly reduced under salt stress compared with normal conditions.However, MC-priming significantly increased the activities of SOD, POD, and CAT by 26.2%, 46.8%, and 5.1%,respectively, which were evaluated by a comparison with the W-priming under salt stress (Fig.3A, C, E).No significant differences were observed in the expression levels ofGhPOD5andGhCAT1between the Wand MC-priming treatments (Fig.3D, F).The addition of fluridone and 3-MP to the treatments significantly reduced the activities of the antioxidant enzymes under normal and salt conditions compared with the treatment that lacked inhibitors (Fig.3A, C, E).However,the fluridone + and 3-MP + salt stress treatments also resulted in much higher levels of antioxidant enzyme activities (except for the POD activity under the fluridone + salt stress treatment) in MC-priming than that of W-priming.Moreover, the expression levels of the genes for these antioxidant enzymes were consistent with their enzymatic activities under inhibitor + salt stress treatment conditions.Thus, these results suggest that the ABA- and GABA-dependent pathways could influence the gene expression levels and activities of antioxidant enzymes.However, the higher activities of antioxidant enzymes induced by MC-treatment might not play a very important role in the tolerance to salt stress mediated by MC μ.
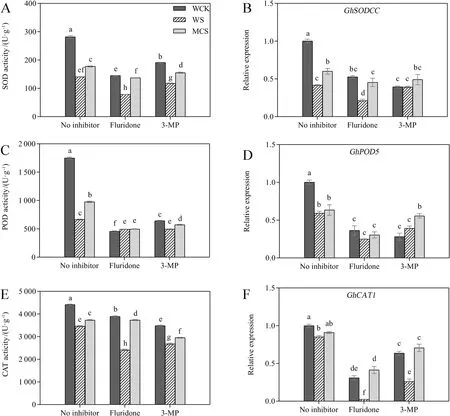
Fig.3 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on the antioxidant enzyme activities and related genes expression levels of the W- and MC-primed cotton seeds under salt stress.A Superoxide dismutase (SOD), C peroxidase(POD), and E catalase (CAT) activities of the W- and MC-primed cotton seeds after 24 h of salt stress.The expression levels of B GhSODCC, D GhPOD5, and F GhCAT1 in the W- and MC-primed seeds after 12 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS:water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.The bars represent means ± SE of three replicate samples
Effects of MC-priming on the AsA-GSH cycle under salt stress
AsA-GSH pathway can also produce antioxidants to eliminate ROS.Under salt stress, the activities of the enzymes related to the AsA-GSH pathway were down-regulated in the germinated cotton seeds (Fig.4A, C, E, G).However, the activities of enzymes that quench H2O2, such as APX, DHAR, GR, and GPX, increased significantly in the MC-primed seeds compared with the W-primed ones.The expression levels ofGhAPX2,GhDHAR3,GhGR, andGhGPX2were also significantly up-regulated in the MCprimed seeds under salt stress (Fig.4B, D, F, H).Treatments with fluridone and 3-MP significantly reduced the activities of enzymes in the AsA-GSH pathway compared with the treatments that lacked inhibitors (Fig.4A, C, E, G).Surprisingly, there were more clear impacts in MC-priming subjected to salt stress since there were significantly fewer traits described above in MC-priming under the fluridone + and 3-MP + salt stress treatment compared with W-priming.Moreover, those higher transcript levels of the AsA-GSH cycle enzymes coding genes (GhAPX2,GhDHAR3,GhGR,GhGPX2) in MC-priming were almost eliminated by the presence of fluridone and 3-MP (Fig.4B, D, F, H).
Salt stress also increased the accumulation of DHA and GSSG but reduced the contents of AsA and GSH, thus,lowering the GSH and AsA redox potential of the seeds grown under salt stress (Fig.5).Notably, MC-priming balanced the redox potential of the seeds by increasing the contents of AsA and GSH and reducing the contents of DHA and GSSG.This resulted in a substantially higher ratio of AsA/DHA and GSH/GSSG in MC-primed seeds,particularly in comparison with the W-primed ones under salt stress (Fig.5E, F).The higher ratios of AsA/DHA and GSH/GSSG resulted in a favorable cellular environment for cotton seeds to counteract the oxidative damage induced by salt.Interestingly, the inhibition of ABA and GABA synthesis reduced the redox potential, particularly in the MC-primed seeds.For example,fluridone and the 3-MP + salt stress treatment reduced the content of AsA and AsA/DHA ratio but increased the contents of DHA and GSSG in the MC-primed seeds more than in the W-primed ones.These results indicated that the MC-stimulated AsA-GSH pathway, which is either ABA- or GABA-dependent, played a major role in the MC-mediated salt stress tolerance during the establishment stage of cotton seedlings.
Relationships and variation among the growth and biochemical attributes of cotton seeds under different treatments
The biplots of a principal component analysis (PCA)were used to evaluate the effects of different treatments on the growth and biochemical attributes of cotton seeds.Two major principal components, PC1 (Dim1) and PC2(Dim2), had shown a cumulative variance of approximately 83.5% in the dataset (Fig.6).Compared with the WS treatment, MCS clustered closer to WCK, indicating that MCS could alleviate salt toxicity during the stage of seedling establishment.Additionally, the MCS treatment and AsA-GSH cycle-related characteristics occurred in the same quadrant, which indicated that the AsA-GSH cycle significantly improved the tolerance of cotton seeds to salt stress that is mediated by MC-priming.The FWS,PWS, FMCS, and PMCS treatments clustered in different biplots and positively correlated with the ROS indicators, suggesting that the reduction of salt stress tolerance in seeds treated with inhibitors might be primarily due to the increased accumulation of ROS.Therefore, these results demonstrated that ABA and GABA might form a nexus and cross-talk strategy to regulate ROS homeostasis in cotton seeds.
Effects of MC-priming on the metabolism of GABA and ABA under salt stress
To further evaluate the possible relationship between ABA and GABA, we utilized fluridone and 3-MP to analyze the metabolism of ABA and GABA.The physiological analyses showed that the onset of seed germination increased with the accumulation of ABA under salt stress(Fig.7).Furthermore, the MC-priming treatment significantly increased the ABA content by 6.2% compared with the W-priming under salt stress.Similarly, MCpriming also significantly up-regulated the expression levels ofGhNCED2(a key gene in ABA synthesis) andGhCYP707A4(a key gene in ABA degradation) by 1.4-fold and 62.9%, respectively, compared with W-priming under salt stress (Fig.7A, B).These results indicated that MC primarily induced the accumulation of ABA under salt stress by enhancing the synthesis of ABA rather than reducing its degradation.Conversely, 3-MP + salt stress significantly decreased the expression level ofGhCYP707A4but had no significant effect on the expression level ofGhNCED2compared with the treatments that lacked an inhibitor.Similar to the transcription level ofGhNCED2, the 3-MP + salt stress did not have significant effects on the accumulation of ABA (Fig.7C).The MC-priming treatment also significantly enhanced the relative expression level ofGhGAD4compared with the W-priming under salt stress (Fig.7D).Similarly, MCpriming also significantly up-regulated the GAD activity and GABA content under salt stress by 11.7% and 17.5%,respectively, compared with W-priming (Fig.7E, F).The addition of fluridone significantly reduced the expression level ofGhGAD4, GAD activity, and content of GABA(Fig.7D-F).Moreover, a more pronounced reduction of the expression level ofGhGAD4, GAD activity, and GABA content was observed in MC-priming under fluridone + salt treatment.These observations indicated that the up-regulation of GABA mediated by MC-priming might have been induced by the early accumulation of ABA during the stage of seedling establishment under salt stress.
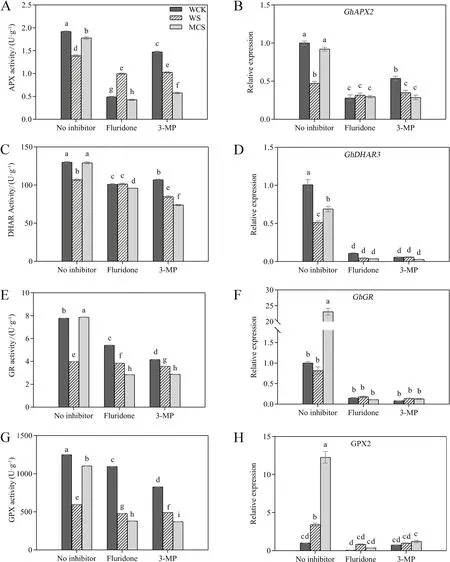
Fig.4 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on the activities and gene expression levels of ascorbate–glutathione (AsA-GSH) pathway enzymes in the W- and MC-primed cotton seeds under salt stress.A Ascorbate peroxidase (APX), C dehydroascorbate reductase (DHAR), E glutathione reductase (GR), and G glutathione peroxidase (GPX)activities of the W- and MC-primed cotton seeds after 24 h of salt stress.The expression levels of B GhAPX2, D GhDHAR3, F GhGR, and H GhGPX2 in the W- and MC-primed cotton seeds after 12 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS: water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.The bars represent the means ± SE of three replicate samples

Fig.5 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on the antioxidant components of the ascorbate–glutathione (AsA-GSH) pathways of the W- and MC-primed cotton seeds under salt stress.A Ascorbic acid (AsA),B dehydroascorbic acid (DHA), C reduced glutathione (GSH), and D oxidized glutathione (GSSG) contents and the ratios of E AsA/DHA and F GSH/GSSG in the W- and MC-primed cotton seeds after 24 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS:water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.The bars represent the means ± SE of three replicate samples
Effects of MC-priming on ABA signaling under salt stress
To explore the effects of MC-priming on the key proteins related to ABA signaling under salt stress, we determined the abundance and phosphorylation levels of the proteins related to ABA signaling in the W- and MC-primed cotton seeds during salt stress in a parallel study (Wang et al.2022).The multi-omics data indicated that salt stress inhibited ABA signaling during the stage of seedling establishment (Fig.8).For example, the abundance of protein phosphatase 2C (PP2C;A0A1U8HXQ2; A0A1U8NV47; and A0A1U8KHI7) significantly increased, while the phosphorylation level of serine/threonine-protein kinase (SAPK2, A0A1U8K326,A0A1U8I9U4, A0A1U8NZB1, A0A1U8K326, and A0A1U8LJ41) significantly decreased in WS_vs_WCK.Although the difference in MCS_vs_WS was not significant, MC-priming slightly alleviated the inhibitory effects of salt stress on ABA signaling.For example, MCpriming substantially up-regulated the phosphorylation level of ABA receptor PYL4 (A0A1U8HYE2) under salt stress compared with WCK (Fig.8B).The abundance of PP2C (A0A1U8NV47 and A0A1U8KHI7) and the phosphorylation level of SAPK2 (A0A1U8K326) showed no significant difference in MCS_vs_WCK (Fig.8).
Effects of MC-priming on Ca2+ signaling under salt stress
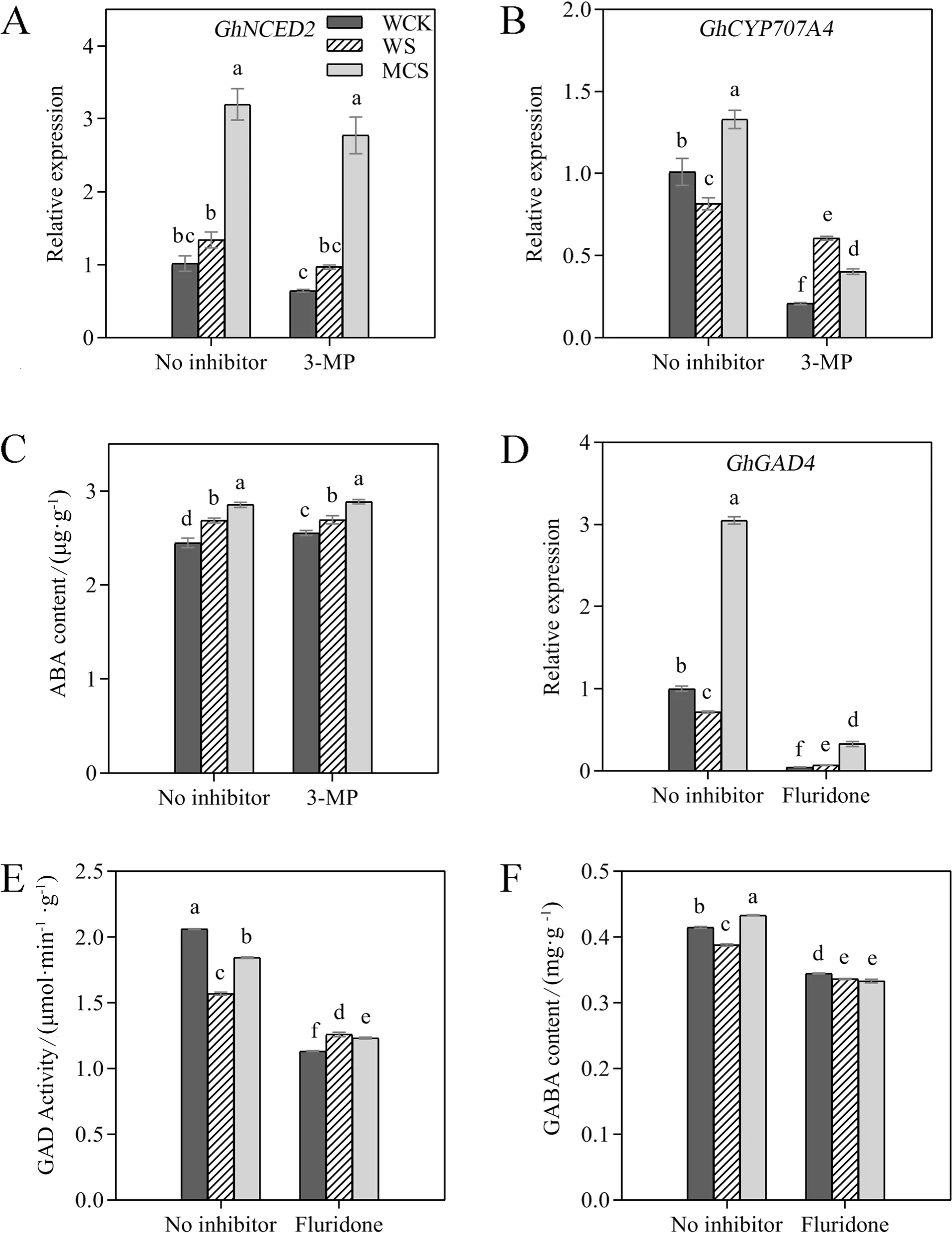
Fig.7 The effects of fluridone (ABA synthesis inhibitor) and 3-mercaptopropionic acid (3-MP; GABA synthesis inhibitor) on abscisic acid (ABA)and γ-Aminobutyric acid (GABA) synthesis in the W- and MC-primed cotton seeds under salt stress.The expression levels of A GhNCED2,B GhCYP707A4, and D GhGAD4 in the W- and MC-primed cotton seeds after 12 h of salt stress.C ABA and F GABA contents, and E GAD activity of the W- and MC-primed cotton seeds after 24 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS: water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.The bars represent the means ± SE of three replicates
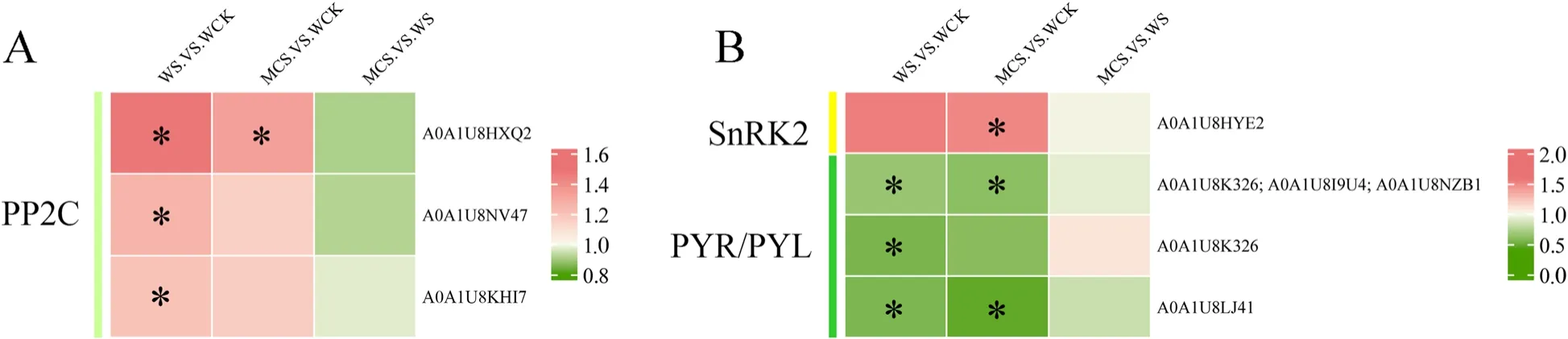
Fig.8 Heatmaps of the enriched abscisic acid (ABA) signal transduction pathways in the W- and MC-primed cotton seeds under salt stress.A The differentially expressed proteins (DEPs) and B differentially regulated phosphoproteins (DRPPs) related to ABA signal transduction pathways of the cotton seeds under WS_vs_WCK, MCS_vs_WCK, MCS_vs_WS treatments.The same name refers to the different phosphorylation forms of the protein.WCK: water-primed seeds germinated in the sand without NaCl; WS: water-primed seeds germinated in the sand containing NaCl;MCS: MC-primed seeds germinated in the sand containing NaCl.* represents significant differences at P < 0.05 (t-test)
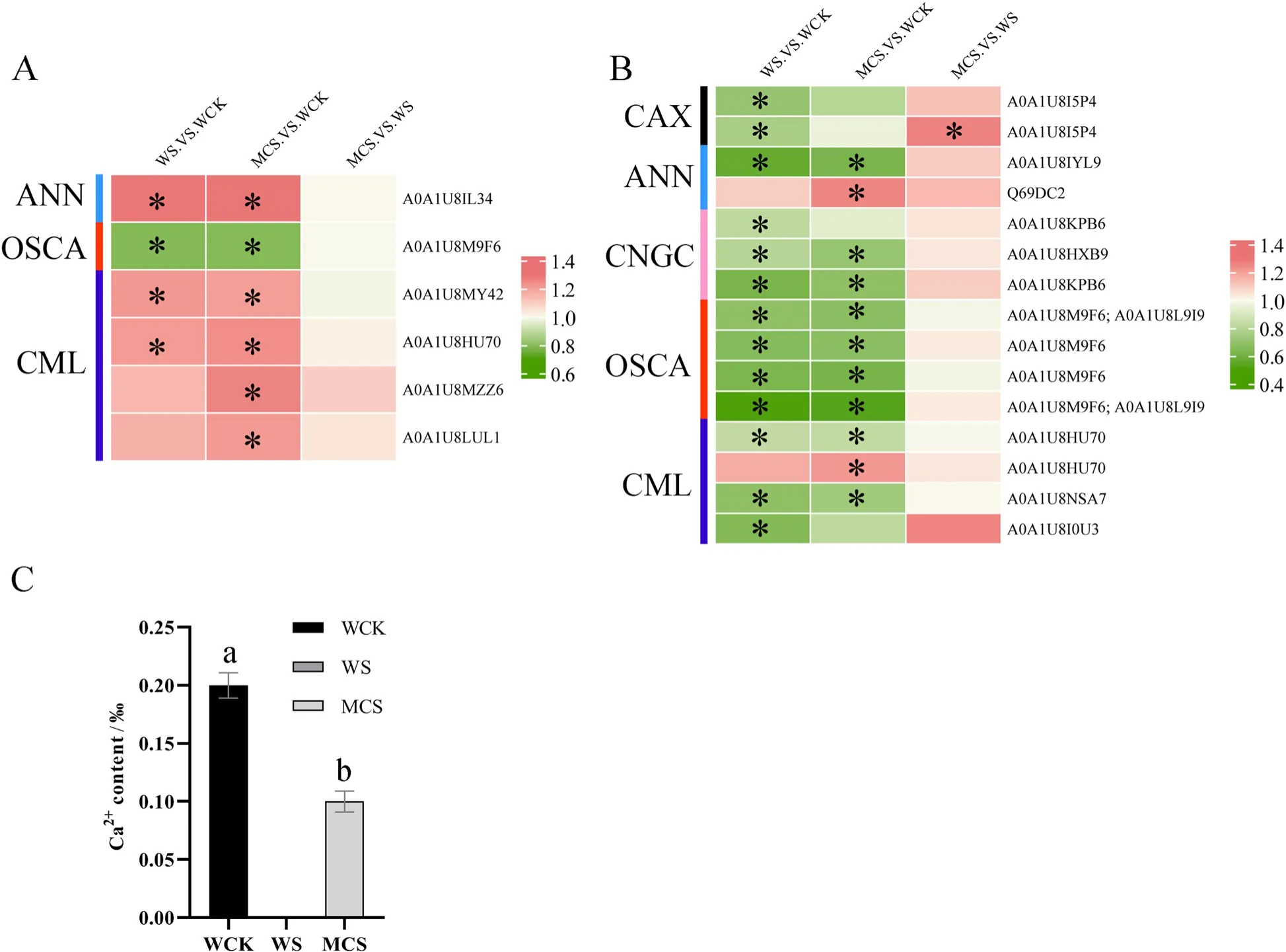
Fig.9 Heatmaps of the enriched calcium ion (Ca2+) homeostasis in the W- and MC-primed cotton seeds under salt stress.A The differentially expressed proteins (DEPs) and B differentially regulated phosphoproteins (DRPPs) related to Ca2+ channels/transporters and calmodulin-like protein(CML) in the WS_vs_WCK, MCS_vs_WCK, MCS_vs_WS treatments.The same name refers to the different phosphorylation forms of the protein.C Ca2+ content in the hypocotyl regions of the W- and MC-primed cotton seeds after 24 h of salt stress.WCK: water-primed seeds germinated in the sand without NaCl; WS: water-primed seeds germinated in the sand containing NaCl; MCS: MC-primed seeds germinated in the sand containing NaCl.* represents significant differences at P < 0.05 (t-test)
GABA synthesis is primarily stimulated by salt stress due to an increase in the concentration of cytosolic Ca2+and the stimulation of CML and activity of GAD (Ji et al.2018; Yang et al.2016).Therefore, we identified the DEPs and DRPPs enriched in the Ca2+channels/transporters and CML to further explore the effects of MC-priming on cytosolic Ca2+increase/oscillations (Fig.9).The results showed that salt stress affected the Ca2+channels/transporters of the germinating seeds during the stage of seedling establishment.These Ca2+channels/transporters were identified in WS_vs_WCK and included the phosphorylation-related vacuolar cation/proton exchanger(CAX, A0A1U8I5P4 and A0A1U8I5P4), annexin(ANN, A0A1U8IYL9), cyclic nucleotide-gated ion channel (CNGC, A0A1U8KPB6, A0A1U8HXB9, and A0A1U8KPB6), and calcium-permeable stress-gated cation channel 1 (OSCA1, A0A1U8M9F6, A0A1U8L9I9,A0A1U8M9F6, A0A1U8M9F6, A0A1U8M9F6, and A0A1U8L9I9) (Fig.9B).Furthermore, the abundance of ANN (A0A1U8IL34) protein significantly increased,while that of OSCA1 (A0A1U8M9F6) significantly decreased in WS_vs_WCK (Fig.9A).The abundance of DEP (A0A1U8MY42 and A0A1U8HU70) and DRPP proteins related to CML increased, while there was a decrease in the phosphorylation of three DRPP proteins (A0A1U8HU70, A0A1U8NSA7, and A0A1U8I0U3)in the WS compared with the WCK treatment (Fig.9A,B).However, MC-priming mitigated the adverse effects of salt stress on Ca2+increase/oscillations.For example, the phosphorylation level of ANN (Q69DC2) and CML (A0A1U8HU70) and the abundance of CML(A0A1U8MZZ6 and A0A1U8LUL1) were highly up-regulated in MCS_vs_WCK in contrast to those in WS_vs_WCK.The phosphorylation of CAX (A0A1U8I5P4) was significantly up-regulated in MCS_vs_WS.However, the phosphorylation level of CAX (A0A1U8I5P4), CNGC(A0A1U8KPB6), and CML (A0A1U8I0U3) remained unchanged in the MCS compared with WCK.These results were further supported by the increased oscillations of Ca2+in the MC-primed cotton (Fig.9C, S1).
Discussion
Seed germination marks the beginning of the plant life cycle and is the plant growth stage that is most sensitive to environmental factors (Lai et al.2020).As a major abiotic stress, salt stress has been reported to reduce the percentage of seed germination, germination index, and antioxidant enzyme activities of crop seedlings (Li et al.2019).Consistent with the previous reports, our results showed that salt stress strongly influenced the germination of cotton seeds and the establishment of seedlings.We also found that MC-priming positively regulated the germination of cotton seeds and seedling emergence under salt stress by alleviating oxidative stress through enhanced detoxification of ROS.These data are consistent with those previously reported on cotton seedlings (Wang et al.2010, 2014, 2022).However, most of these previous studies were observational and did not have a physiological and molecular basis.In this study,we showed that MC-priming stimulated ABA signaling to activate the GAD-dependent GABA synthesis, and increased the activities of the AsA-GSH pathway, thus resulting in enhanced ROS scavenging in response to salt stress (Fig.10).
The harmful effects of salt stress was manifested by the excessive accumulation of ROS, which negatively impacts plant metabolism (Julkowska and Testerink 2015; Yang and Guo 2018).To maintain ROS homeostasis, plants activate their antioxidant enzymes (including SOD, CAT,POD, APX, GR and others) and non-enzymatic defense system components (AsA, GSH, and other antioxidants)(Qamer et al.2021).Thus, plants with better antioxidant defenses are always associated with higher tolerance to salt stress.In this study, we showed that the components of the antioxidant defense system was also activated in MC-primed seeds in response to salt stress.We found that MC-priming stimulated the antioxidant system of the cotton seeds under salt stress, which resulted in effi-cient ROS scavenging.This was consistent with the lower levels of MDA and ROS, such as H2O2and O2.-, in the MC-primed seeds under salt stress.Thus, these findings suggest that MC-priming improved the salt tolerance of cotton seeds by increasing the ability of the antioxidants to scavenge ROS.Our results are also consistent with those of previous studies which showed that MC can alleviate the effects of oxidative stress by improving the antioxidant capacity of various plant species (Wang et al.2022; Liu et al.2015).
Numerous studies suggest that complex signal transduction pathways are engaged in the antioxidant defense response against salt stress and other oxidative stresses(Qamer et al.2021).In this study, the addition of inhibitors of ABA and GABA biosynthesis to the treatments significantly decreased the gene expression levels and activities of the antioxidant enzymes related to the AsAGSH cycle.This suggested that the ABA- and GABAdependent signaling pathways are required to induce the antioxidant defense system under salt stress.It was observed that higher antioxidant enzyme activities and the coordinated adjustment of the redox status of antioxidants are related to salt stress tolerance in various plants (Julkowska and Testerink 2015).However,our results indicated that the AsA-GSH cycle while not antioxidant enzymes played a major role in the tolerance to salt stress mediated by MC, possibly due to three reasons: 1) Our results revealed that the higher activities of antioxidant enzymes could not explain the insignificant difference between the growth indices under stress treatments augmented with fluridone and 3-MP.2) The presence of fluridone and 3-MP reduced the activities of antioxidant enzymes related to the AsAGSH in MC-priming, which significantly reduced the redox potential of cotton seeds under salt stress; and 3) The MCS treatment and the AsA-GSH cycle-related characteristics occurred in the same quadrant of the PCA, suggesting a close relationship between the AsAGSH cycle and salt stress tolerance.These results led us to hypothesize that the tolerance to salt stress mediated by MC during the stage of seedling establishment can be primarily facilitated by the MC-induced AsA-GSH cycle, which is partly mediated by ABA- or GABAdependent signaling.
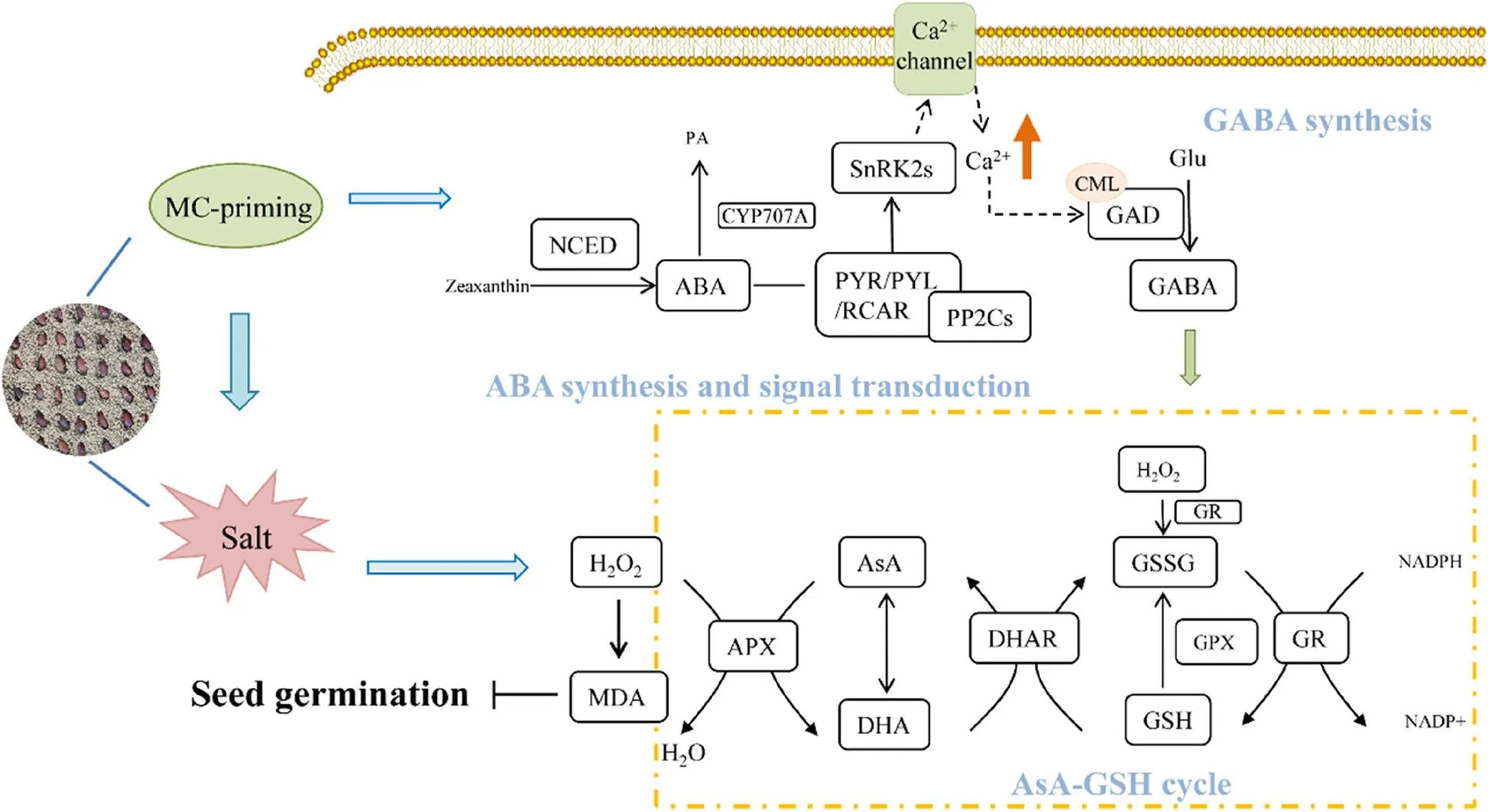
Fig.10 A model showing how MC-priming alleviated the inhibitory effects of salt stress on cotton seed germination
Previous studies suggested that changes in the levels of ABA alter those of GABA under several abiotic stresses,including salt stress (Bown and Shelp 2016; Yang et al.2016; Wang et al.2021a, 2022; Ji et al.2018).This suggests that cross-talk between ABA and GABA may mediate the tolerance to salt stress.Our results showed that MC-priming coordinated the enhancement of contents of ABA and GABA under salt stress by increasing the expression levels ofGhNCED2andGhGAD4and the activity of GAD.The inhibition of ABA synthesis reduced the positive effects of MC-priming on GABA content,while the inhibition of GABA synthesis did not significantly affect the MC-priming effects on ABA content.Thus, the increased expression level of ABA synthetic genes that is induced by MC might occur upstream of the GABA synthetic genes by up-regulating gene expression and GAD activity under salt stress.This hypothesis is consistent with previous research that exogenous ABA resulted in the accumulation of GABA in germinated fava beans (Vicia fabaL.) under salt stress by activating the activity of GAD (Yang et al.2016).
GAD is a CaM-binding protein whose activity can be regulated by Ca2+oscillations (Renault et al.2013; Jiao et al.2022).Previous studies indicated that SnRK2 (a key protein in ABA signal transduction) can promote the internal flow of Ca2+by activating the Ca2+channels/transporters (CAX, ANN, CNGC, and OSCA1) in response to salt stress (Liu et al.2021; Lee et al.2004;Laohavisit et al.2009; Su et al.2021).Therefore, we determined whether the differences in Ca2+oscillations affected the accumulation of GABA by regulating ABA signaling.The results indicated that the higher abundance or phosphorylation level of the proteins related to ABA signaling, CML, and Ca2+channels/transporters in MC-priming up-regulated the content of Ca2+and the activity of AsA-GSH cycle compared with W-priming.A possible explanation for this phenomenon is that MC-priming intrinsically alleviates the accumulation of glutamate (Glu) under salt stress(Wang et al.2022) through a common precursor of GABA and GSH (Hasanuzzaman et al.2019; Qiu et al.2020).Similarly, studies have indicated that the foliar or seed application of Glu in the greenhouse or field can improve the antioxidant capacity of crops by activating antioxidant enzymes (Qiu et al.2020).However,the physiological rationale behind the enhancement of activity of AsA-GSH cycle induced by GABA remains elusive (Seifikalhor et al.2019).There is also the possibility that ABA might directly regulate the activity of AsA-GSH cycle (Hasanuzzaman et al.2019).Thus,further studies are merited to understand the association of GABA and ABA with the AsA-GSH cycle at the genetic and molecular levels.
Conclusions
In summary, salt stress significantly repressed the antioxidant capacity of cotton seeds by increasing the accumulation of ROS, thus, inhibiting seedling establishment under salt stress conditions.However, MC-priming conferred salt stress tolerance of cotton seeds by activating AsA-GSH cycle, thereby enhancing their antioxidant capacity during seed germination.This trait was mediated by MC-induced ABA signaling that operated upstream of GAD-generated GABA synthesis by activating the oscillations of Ca2+.This increased the activities of AsA-GSH pathway enzymes and enhanced the redox potential of AsA and GSH.Therefore, our findings help to provide new insights into the physiological and molecular mechanisms used by MC-priming to confer the tolerance of cotton to salt stress and valuable theoretical guidance to breed cotton that is resistant to salt stress.
Materials and methods
Germination conditions
Cotton seeds (Gossypium hirsutumL.cultivar J1020)were used in this study.The seeds were primed and germinated as previously described (Wang et al.2022).Briefly, healthy and plump cotton seeds were first soaked in 200 mg·L-1MC (MC-priming) or deionized water (W-priming) in the dark at 30 ℃ for 12 h.After priming, the seeds were sown in plastic trays(24 cm × 24 cm × 4 cm) containing 0 (control: CK) and 0.3% (w/w) NaCl (salt stress: S) of sandy clay soil (13%moisture content) at 30 ℃ in a dark growth chamber(Wang et al.2022).Furthermore, 1.3 μg·g-1of fluridone, an inhibitor of ABA synthesis, or 35 μg·g-1of 3-mercaptopropionic acid (3-MP), an inhibitor of GABA synthesis, was applied to acquire more evidence that ABA and GABA are involved in regulating the tolerance of cotton seeds to salinity stress.Thus,there were three groups with three treatments each: i)the group that lacked inhibitors; water-primed seeds germinated in the sand without NaCl (WCK); waterprimed seeds germinated in the sand that contained NaCl (WS); and MC-primed seeds germinated in the sand that contained NaCl (MCS); ii) the fluridone group: fluridone was added based on the WCK, WS,and MCS treatments; and iii) the 3-MP group: 3-MP was added based on the WCK, WS, and MCS treatments.The seeds were germinated in a dark incubator,and the plastic trays that contained the seeds were covered with black plastic bags to prevent the evaporation of moisture.Each treatment was replicated three times (three plastic trays), and each tray contained 100 seeds.
Seed germination traits
The seed germination rate and the root length of the germinated seeds were determined at 1, 2, 3, and 4 d after sowing.The seed vigor index was calculated as described by Wang et al.(2021a and b).
Determination of the peroxidation of membrane lipids of seeds
The germinated seeds sampled from each treatment were ground to powder in liquid nitrogen at 24 h after sowing.A total of 0.1 g of powder was used to determine the content of MDA with an MDA kit (BC0025,Solarbio, Beijing, China) according to the manufacturer’s instructions.The absorbance of the extract was calculated at 600 nm and 532 nm to determine the level of MDA.
Determination of Ca2+ concentration in the hypocotyl regions
After 24 h of germination, the seed samples were collected, pretreated, and examined using a transmission electron microscope (TEM, JEOL 2100, Tokyo, Japan),as described by Wang et al.(2021a and b).Briefly, the samples were subjected to X-ray microanalysis, and the relative weights of mineral ions in the embryogenic hypocotyl cells were quantified automatically based on the net K-shell X-ray peak counts after the background X-ray counts had been subtracted.
Analysis of protein abundance and phosphorylation levels of the related differentially expressed proteins
Preliminary results indicated that the seed germination stimulated by MC-priming was associated with increased ABA, GABA, and ROS metabolism under salt stress (Wang et al.2021a, 2022).Therefore, we analyzed the recently published proteomic and phosphoproteomic datasets (Wang et al.2022) to identify the abundance and phosphorylation levels of the proteins linked to ABA signal transduction, Ca2+channels/transporters, and calmodulin-like protein (CML)in salt-treated germinated seeds.From the proteomics profiling, several differentially expressed proteins(DEPs) were involved in the ABA signal transduction.These DEPs and the differentially regulated phosphoproteins (DRPPs) were selected atP< 0.05(t-test) and > 1.2- or < 0.83-fold change.The proteomic and phosphoproteomic analysis datasets are available from the ProteomeXchange Consortium (http:// prote omece ntral.prote omexc hange.org) using the dataset identifier PXD032890.
Determination of ROS level
To visually display the accumulation of ROS in different treatments, we performed histochemical assays as described by Li et al.(2022).Briefly, freshly germinated seeds were collected after 24 h of germination.The seeds were then placed in 50 mmol·L–1Tris–acetate buffer (pH 5.0) that contained 1 mg·mL-13,3’-diaminobenzidine (DAB) or 10 mmol·L–1potassium phosphate buffer (pH 7.8) that contained 0.25 mg·mL-1nitro tetrazolium blue chloride (NBT) for 1 h in the dark.The samples were then noted for their degree of staining.The contents of hydrogen peroxide (H2O2) and superoxide anion (O2.-) of the samples were analyzed using assay kits from Solarbio (BC3595 and BC1295) as described by Wang et al.(2022).
Quantification of the activities of antioxidant enzymes
After 24 h of germination, 0.1 g of freshly prepared seed samples were homogenized in phosphate-buffered saline (PBS) (pH = 7.8) that was supplemented with 1.34 mmol·L–1EDTA-Na2in an ice bath.The supernatants were obtained by centrifugation at 4 ℃ for 15 min at 8 000gas described by Wang et al.(2022).The supernatant was used to measure the activities of SOD, peroxidase (POD), CAT, APX, DHAR, GR, and GPX using their respective enzyme activity assay kits (BC0170, BC0090,BC0200, BC0220, BC0665, BC1160, and BC1195) from Solarbio.
Determination of the contents of antioxidants
To determine GSH and GSSG contents of the seeds,0.1 g of thoroughly ground powder of the germinated seeds were homogenized with 1 mL of 0.2 mmol·L–1potassium phosphate buffer that contained 5 mmol·L–1EDTA, pH 8.0, as described by Alpert and Gilbert(1985).The homogenate was then centrifuged for 10 min at 4 ℃ at 8 000g, and the supernatant was collected to analyze the contents of antioxidants.The contents of GSH and GSSG were measured based on the reaction between GSH and 5,5’-dithiobis-2-nitrobenoic acid, respectively, using their respective assay kits(BC1175 and BC1180) (Solarbio).Furthermore, another 0.1 g of sampled seeds were homogenized with 1 mL of 5% trichloroacetic acid as described by Ji et al.(2018)and centrifuged for 20 min at 4 ℃ at 16 000g.The supernatant was used to measure the contents of AsA and DHA of the germinated seeds under different treatments.The contents of AsA and DHA were defined based on the redox reaction of AsA using their respective assay kits (BC1230 and BC1240) (Solarbio) according to the manufacturer’s instructions.
Determination of the contents of ABA and GABA and the activity of GAD
Approximately 0.1 g of frozen cotton seeds sampled at 24 h after imbibition were ground into powder to determine their contents of ABA and GABA and activity of GAD.The ABA was quantitatively analyzed using an ABA ELISA kit (MM-1185O1, Mmbio, Jiangsu, China).The content of GABA and activity of GAD were measured as described by Wang et al.(2022) using their respective assay kits (GABA-1-W and GAD-1-W, Omin,Shanghai, China).
Quantitative real-time polymerase chain reaction(qRT-PCR)
The expression levels of the genes engaged in hormone metabolism, GABA synthesis, and ROS metabolism were defined by qRT-PCR (RR420A, TaKaRa, Dalian,China).The primers that are shown in Table S1 were designed based on the gene sequences obtained from the NCBI GenBank database (https:// www.ncbi.nlm.nih.gov [accessed on August 30, 2022]) and the Cotton Functional Genomics Database (https:// cotto nfgd.org/[accessed on August 30, 2022]).
Total RNA was extracted from the germinated cotton seeds at 12 h after sowing using a RNeasy kit (Tiangen,Dalian, China).The quantity and purity were measured using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).A volume of 1 μL of the RNA samples was then reversed transcribed into cDNA using a cDNA synthesis kit (RR036A,TaKaRa).The relative gene expression levels were determined using the 2-ΔΔCTmethod with theActingene as the internal reference.There were three biological replicates per gene.
Statistical analysis
Principal component analysis (PCA) of indicators were analyzed by Origin 2021.The data were analyzed in three biological replicates using a one-way analysis of variance(ANOVA) on SAS software (SAS 9.4, Cary, NC, USA).The ANOVA was performed to test for significant differences between treatments and utilized the least significant difference (LSD) test atP< 0.05.
Supplementary Information
The online version contains supplementary material available at https:// doi.org/ 10.1186/ s42397- 023- 00162-x.
Additional file 1: Fig.S1.The analysis results of the energy dispersive spectrometer for (A) WCK (B) WS and (C) MCS treatments of hypocotyl regions after 24 h of salt stress.
Additional file 2: Table S1.Primer sequences required for qRT-PCR.
Acknowledgements
Thank Institute of Cotton Research of Chinese Academy of Agricultural Sciences and Western Agricultural Research Center of Chinese Academy of Agricultural Sciences for providing germplasm for this work.We thank the Mogo Internet Technology Co., Ltd.(http:// www.mogoe dit.com/) for its linguistic assistance during the preparation of this manuscript.
Authors’ contributions
Wang XR, Wang N, Yan G, and Zhang XL conceived and designed the experiments.Qi Q and Wang N performed the experiments, analyzed the data, and wrote the the manuscript.Ruan SJ, Zhang HH, Shi JB, Dong Q, and Xu QG contributed the reagents and materials.Muhammad N improved the written language.Wang XR, Yan GT, Zhang XL, and Song MZ reviewed and edited the manuscript.All authors read and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China(32001481), the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences, the China Agriculture Research System, the National Modern Agricultural Industry Technology System of China (CARS-18–05), the Provincial Key R&D and Promotion Special Projects in Henan (232102110178), the Program for Key Areas of Science and Technology of Xinjiang Production and Construction Corps Third Division and Tumsuk City (KY2021GG08), and the Central Public-interest Scientific Institution Basal Research Fund (1610162023019).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Author details
1National Key Laboratory of Cotton Bio-Breeding and Integrated Utilization,Institute of Cotton Research of Chinese Academy of Agricultural Sciences,Anyang 455000, Henan, China.2Institute of Western Agriculture, the Chinese Academy of Agricultural sciences, Changji 831100, Xinjiang, China.
Received: 8 October 2023 Accepted: 28 November 2023
杂志排行

Journal of Cotton Research的其它文章
- Are yarn quality prediction tools useful in the breeding of high yielding and better fibre quality cotton (Gossypium hirsutum L.)?
- Risk of control failure to insecticides malathion, profenophos + cypermethrin mixture, and fipronil in boll weevil (Coleoptera:Curculionidae) populations from Bahia, Brazil
- Effects of mepiquat chloride and plant population density on leaf photosynthesis and carbohydrate metabolism in upland cotton
- Feasibility study of assessing cotton fiber maturity from near infrared hyperspectral imaging technique
- Enhancing waterlogging tolerance in cotton through agronomic practices
- Biocontrol potential of entomopathogenic nematode, Heterorhabditis indica against pink bollworm, Pectinophora gossypiella (Saunders)(Lepidoptera: Gelechiidae)