母鹅日粮添加甜菜碱通过IGFs信号通路促进子代胸肌肌纤维肥大
2024-01-10庄新娟王文正赵茹茜
马 帅,王 燕,庄新娟,王文正,赵茹茜*
(1.南京农业大学动物医学院,南京 210095;2.农业农村部动物生理生化重点实验室,南京 210095;3.江苏立华牧业股份有限公司,常州 213168)
江南白鹅是由江苏立华牧业有限公司培育出的中型白鹅配套系(三系配套),该品种具有耐粗饲、体型紧凑、肉质鲜美、适应性强等优点,目前已在不同地区广泛养殖。然而,与一些优秀肉鹅品种相比[1-2],江南白鹅的生长发育较慢,屠宰性能较低,因此,需要进一步提高江南白鹅的生产性能,以提高该品种在市场中的竞争力。
家禽骨骼肌约占体重的40%,是体内最大的蛋白质储存库[3],同时,胸肌和腿肌是家禽生产的主要产品之一,肌肉产量与其生产性能密切相关。骨骼肌发育过程受到正向和负向因子的双向调控,其中,生肌调节因子家族(myogenic regulatory factors,MRFs)属于正向调节因子,包括肌源性分化因子1(myogenic differentiation factor 1,MyoD 1)、生肌因子5(myogenic factor 5,Myf5)、肌细胞生成素(myogenin,MyoG)、生肌调节因子4(myogenic regulatory factor 4,MRF4),以上4种生肌调节因子共同促进肌肉的形成、分化和增殖[4-6]。而肌肉生长抑制素(myostatin,MSTN)作为骨骼肌生长发育的负向调节因子,可抑制出生前骨骼肌的发育和出生后骨骼肌细胞的肥大。
肌纤维数目是决定肌肉重量的一个重要因素,且在出生前就已经固定。出生后肌纤维的生长主要与肌纤维类型的转化、肌纤维的增长和肥大有关。其中,肌纤维的增长主要依赖于卫星细胞的分裂以增加新的细胞核[7-8],而肌纤维肥大与肌纤维中蛋白质沉积有关。蛋白质合成受胰岛素样生长因子1/2(insulin-like growth factor-1/2)信号通路的调控[9],而该过程由哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)介导。mTOR是肌肉中蛋白质合成的关键调控蛋白,其中mTOR复合物1(mTORC1)通过磷酸化下游两种关键效应底物p70核糖体S6激酶(p70S6K)和真核翻译起始因子4E结合蛋白1(eukaryotic initiation factor 4E binding protein 1,4EBP1)发挥作用[10-13]。
甜菜碱(betaine,BET)广泛存在于多数生物体中,作为营养型饲料添加剂,添加在家禽日粮中具有促进动物生长[14-16]、调节机体代谢[17-18]等功能。研究表明,饲粮中添加甜菜碱通过激活IGF-1-mTOR信号通路,提高血液和胸肌中IGF-1含量,从而增加青脚麻鸡及樱桃谷鸭的胸肌重量[15-16,19]。饲粮中添加甜菜碱可以显著提高63和70日龄子代鹅的全净膛率,但未影响到胸肌率和腿肌率[14,20]。此外,甜菜碱通过提高MyoD、Myf5和MyoG蛋白水平促进小鼠C2C12成肌细胞的分化[21]。本实验室前期研究发现,在母猪[22]或蛋鸡[23]日粮中补充甜菜碱,可通过表观遗传机制调节子代肝组织中胆固醇代谢相关基因的表达,从而影响血液和肝中胆固醇水平。母鼠在哺乳期摄入较高水平的甜菜碱,还能降低后代小鼠成年时期的肥胖率,并提高机体葡萄糖的稳态[24]。
目前,关于甜菜碱的研究主要集中于动物的生长发育以及通过改变母体营养状况来调控子代机体的代谢。然而,关于甜菜碱对不同家禽胸肌产量的影响存在不一致的研究结果。至今尚未见有关母鹅日粮添加甜菜碱对子代鹅胸肌生长发育影响的报道。本试验主要目的是研究在母鹅日粮中添加甜菜碱对子代鹅胸肌生长发育及相关分子机制的影响,以期为甜菜碱在鹅业生产应用提供参考。
1 材料与方法
1.1 试验动物
试验选用540只体重相近的39周龄江南白鹅(由江苏立华牧业股份有限公司提供),公母比例1∶5(90只公鹅、450只母鹅)。
1.2 试验设计及饲粮
将试验鹅随机分为3组,每组5个重复,每个重复36只,对照组(CON)饲喂基础日粮,处理组在基础日粮中分别添加0.25% (2.5 g·kg-1, LBT)、0.5%(5 g·kg-1HBT)甜菜碱(纯度≥75%,购自北京昕大洋科技发展有限公司),试验期为7周。第7周开始连续收集7 d种蛋并进行孵化。出雏后,各组选取160只体重相近的健康雏鹅,分为5个重复,每个重复32只(雌雄各半)。所有组别的子代鹅均饲喂相同的基础日粮,试验期为63 d。种鹅和子代鹅基础日粮组成及营养水平见表1。

表1 基础日粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of basal diets (air-dry basis) %
1.3 饲养管理
试验于2020年8月~2021年1月在金坛市四季禽业种鹅场进行,试验鹅全程舍内漏缝塑料板高床架饲养,自由采食、饮水,种鹅每日16 L∶8D、子代鹅每日24 h光照。按常规免疫程序注射疫苗和消毒。
1.4 样品采集与处理
1.4.1 生长性能测定 分别于21、42和63日龄时,按重复称量试验鹅的空腹体重(称重前禁食8 h),统计1~21日龄、22~42日龄、43~63日龄的采食量,计算各组平均日采食量(ADFI)、平均日增量(ADG)和料肉比(F/G)。
1.4.2 胸肌样品采集 在35和63日龄时,从每个重复随机选取2只雄性子代鹅称重后屠宰,剥离两侧的胸肌并称重,计算胸肌率。采集左侧胸肌,一部分经液氮速冻后,转移至-80 ℃保存,另一部分置于4%多聚甲醛溶液中用于制备石蜡切片,并进行苏木精和伊红(HE)染色。
1.5 测定指标与方法
1.5.1 胸肌率的计算 根据胸肌的重量和宰前活体重计算35和63日龄的胸肌率,胸肌率=胸肌重/活体重×100%。
1.5.2 胸肌核酸含量和蛋白质含量测定 采用优化过的Johnson和Chandler[25]的方法对胸肌中DNA和RNA含量进行测定,利用DNA和RNA不同的水解特性将二者分开,经分光光度计紫外吸收法测定各自的浓度(单位:μg·mg-1)。用BCA蛋白质试剂盒(DQ111,全式金)测定胸肌中蛋白浓度。
1.5.3 胸肌组织形态学分析 将固定好的胸肌样本经酒精脱水、透明、浸蜡包埋后连续4 μm切片进行HE染色。每张切片置于显微镜(AX70,Olympus, 日本)下观察和拍照,选取5个清晰的视野,使用Image-Pro-Plus测量肌纤维直径(μm)、肌纤维横切面积(μm2)和肌纤维密度(个·mm-2)。
1.5.4 胸肌IGF-1和IGF-2含量测定 使用鹅IGF-1和IGF-2(YB-IGF1-Go、YB-IGF2-Go,上海钰博生物科技有限公司)ELISA检测试剂盒。根据说明书步骤检测胸肌中IGF-1和IGF-2含量。即30 mg胸肌样品经PBS匀浆、离心后取上清。将稀释后的上清液经温育、酶标试剂、显色和终止反应后,由酶标仪测定OD值。以标准品的浓度为横坐标,OD值为纵坐标,绘出标准曲线,根据样品OD值由标准曲线计算相应的浓度。
1.5.5 胸肌总RNA提取和qPCR 采用TRIzol法提取胸肌总RNA,根据反转录试剂盒使用说明书(AU341-02-V2,全式金)反转录为cDNA。在GenBank上找到目的基因的序列并设计靶基因的引物(表2),引物由南京擎科生物公司合成。采用实时荧光定量PCR检测相关基因的转录表达水平,通过2-ΔΔCT法比较组间基因的相对表达量。
1.5.6 胸肌总蛋白提取和Western Blot RIPA总蛋白裂解液的配置(50 mL):1.5 mL 5 mol·L-1NaCl、2.5 mL 1 mol·L-1Tris(pH=8.0)、0.25 g 脱氧胆酸钠、0.5 mL 10% SDS、0.5 mL NP-40以及1%蛋白酶抑制剂(现用现加)。取30 mg研磨过的组织样置于2 mL EP管中,加入研磨珠和600 μL RIPA总蛋白裂解液,在组织匀浆仪上低温匀浆后冰上静置10 min,之后4 ℃ 6 000 r·min-1离心20 min,取上清,并用BCA法测定上清液蛋白浓度。通过SDS-PAGE凝胶电泳分离目标蛋白质。将NC膜置于5%脱脂奶粉中室温封闭2 h,加入一抗(GAPDH抗体,A19056,ABclonal,1∶12 000;MyHC-I抗体,BA-F8,DSHB,1∶2 000;AKT抗体,AP0059,Bioworld,1∶200;p-AKT抗体,BS4009,Bioworld,1∶200;mTOR抗体,2938,Cell Signaling Technology,1∶1 000;p-mTOR抗体,3398,Cell Signaling Technology,1∶1 000),4 ℃过夜孵育,次日经TBST缓冲液洗3次,常温孵育二抗1 h,将NC膜浸泡在ECL发光液(bl523,Biosharp)中2 min后使用蛋白质成像仪(VeralDoc 4000 MP,BIO-RAD,美国)进行拍照,并用Image J分析蛋白条带灰度值。
1.5.7 统计分析 采用IBM SPSS 20.0统计软件中One-way ANOVA对组间试验数据进行单因素方差分析,并用LSD法进行组间多重比较,所有数据以“平均值±标准误”表示,以P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 母鹅日粮添加甜菜碱对子代鹅体重、胸肌重和胸肌指数的影响
由图1A可知,21日龄LBT和HBT组以及42日龄和63日龄HBT组全群体重显著高于对照组(P<0.01)。由图1B可知,1~21日龄时,两个处理组平均日增重显著高于对照组(P<0.01),HBT组在1~63日龄平均日增重仍显著高于对照组(P<0.05)。由图1C可知,1~21日龄两个处理组子代鹅的平均日采食量显著高于对照组(P<0.05),该阶段料肉比显著低于对照组(P<0.05,图1D)。对每组随机选取的10只子代鹅生产性能分析发现,35日龄HBT组子代鹅体重和胸肌重量显著高于对照组(P<0.01,图1E、1F)。63日龄时,LBT和HBT组子代鹅体重和胸肌重量显著增加(P<0.05,图1 H、1I),HBT组子代鹅胸肌指数也显著高于对照组(P<0.05,图1J)。

A. 子代鹅体重;B. 子代鹅平均日增重;C. 子代鹅平均日采食量;D. 料肉比;E. 子代鹅35日龄体重;F. 子代鹅35日龄胸肌重量;G. 子代鹅35日龄胸肌指数; H. 子代鹅63日龄体重;I. 子代鹅63日龄胸肌重量;G. 子代鹅63日龄胸肌指数。*. P<0.05,**. P<0.01,下同A. Body weight of offspring goslings; B. Average daily gain of offspring goslings; C. Average daily feed intake of offspring goslings; D. Feed conversion ratio of offspring goslings; E. Body weight of goslings at 35 days of age; F. Breast muscle weight of goslings at 35 days of age; G. Breast muscle index of goslings at 35 days of age; H. Body weight of goslings at 63 days of age (n=10); I. Breast muscle weight of goslings at 63 days of age; J. Breast muscle index of goslings at 63 days of age.*. P<0.05,**. P<0.01, the same as below图1 母鹅日粮添加甜菜碱对子代鹅体重、胸肌重量和胸肌指数的影响(n=10)Fig.1 Effect of maternal betaine supplementation on body weight, breast muscle weight and index of offspring goslings(n=10)
2.2 母鹅日粮添加甜菜碱对子代鹅胸肌组织形态的影响
如图2所示,35日龄时,LBT和HBT组子代鹅胸肌肌纤维直径和横截面积显著增加(P<0.05,图2C、2D),并且肌纤维密度随着肌纤维直径的增加而降低,HBT组子代鹅胸肌肌纤维密度显著低于对照组(P<0.05,图2E)。63日龄HBT组子代鹅胸肌肌纤维直径和横截面积仍然显著高于对照组(P<0.05,图2F、2G),各组间肌纤维密度没有显著差异(P>0.05,图2H)。
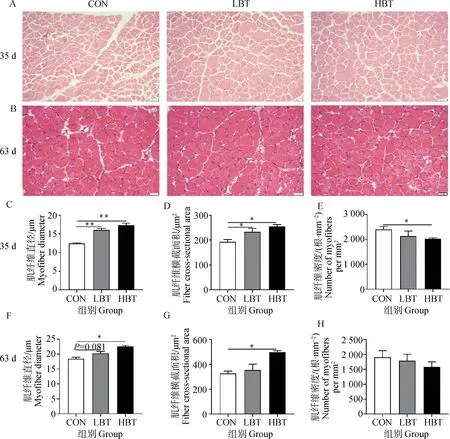
A. 35日龄子代鹅胸肌HE染色分析(400×,比例尺=20 μm);B. 63日龄子代鹅胸肌HE染色分析(400×,比例尺=20 μm);C~E. 35日龄子代鹅胸肌肌纤维直径、横截面积、肌纤维密度;F~H. 63日龄子代鹅胸肌肌纤维直径、横截面积、肌纤维密度A. HE staining analysis of breast muscle of offspring goslings at 35 days of age (400×, scale bar=20 μm); B. HE staining analysis of breast muscle of offspring goslings at 63 days of age (400×, scale bar=20 μm); C-E. Diameter, cross-sectional area and density of breast muscle of offspring goslings at 35 days of age; F-H. Diameter, cross-sectional area and density of breast muscle of offspring goslings at 63 days of age图2 母鹅日粮添加甜菜碱对仔鹅胸肌组织形态的影响Fig.2 Effects of maternal betaine supplementation on the breast muscle morphology of offspring goslings
2.3 母鹅日粮添加甜菜碱对子代鹅胸肌核酸和蛋白质含量的影响
由表3所示,与对照组相比,母鹅日粮中添加甜菜碱显著提高35日龄子代鹅胸肌中RNA含量(P<0.05)以及HBT组RNA/DNA的比值(P<0.05)。此外,母源甜菜碱的影响可以维持到63日龄,HBT组子代鹅胸肌中RNA和蛋白含量显著高于对照组(P<0.05)。
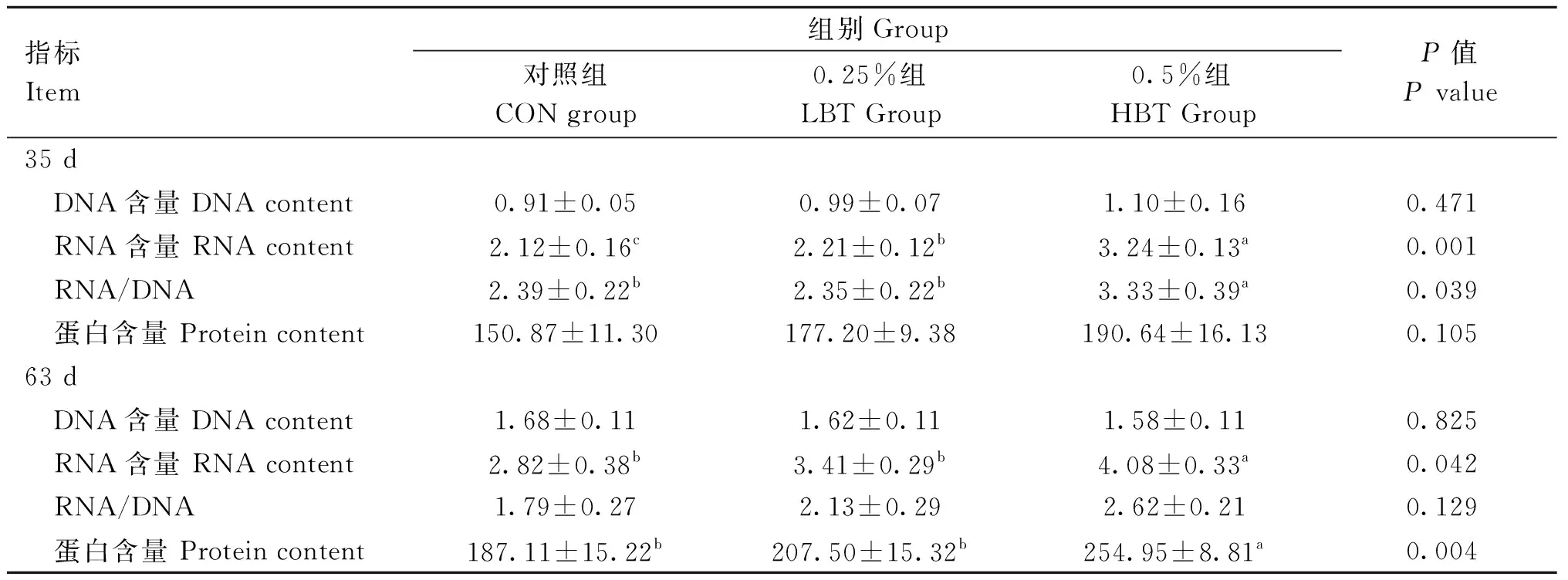
表3 母鹅日粮添加甜菜碱对子代鹅胸肌核酸和蛋白含量的影响Table 3 Effect of maternal betaine supplementation on the nucleic acid and protein contents of breast muscle in offspring goslings (μg·mg-1)
2.4 母鹅日粮添加甜菜碱对子代鹅胸肌肌纤维类型及发育相关基因表达的影响
如图3所示,母鹅日粮添加甜菜碱未显著影响35和63日龄子代鹅胸肌肌纤维类型(P>0.05,图3A~3D)。对生肌调节因子基因表达水平进行检测,结果显示,母鹅日粮中添加不同水平的甜菜碱均可以显著提高35和63日龄子代鹅胸肌中Pax7 mRNA的表达水平(P<0.05,图3E、3F),而对其它指标无显著性影响。
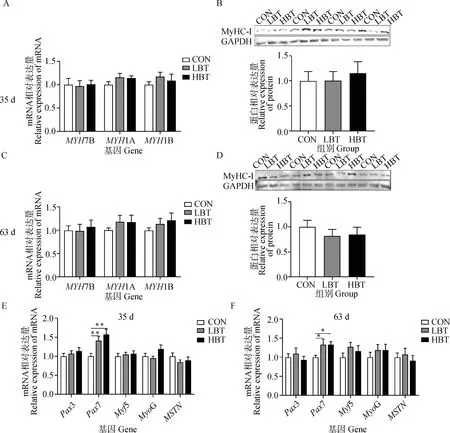
A. 35日龄胸肌肌纤维类型相关基因表达;B. 35日龄胸肌MyHC-I蛋白表达;C. 63日龄胸肌肌纤维类型相关基因表达;D. 63日龄胸肌MyHC-I蛋白表达;E. 35日龄胸肌发育相关基因表达;F. 63日龄胸肌发育相关基因表达A. Expression of breast muscle fiber type related genes at 35 days of age; B. Protein expression of MyHC-I in breast muscle at 35 days of age; C. mRNA expression of breast muscle fiber type related genes at 63 days of age; D. Protein expression of MyHC-I in breast muscle at 63 days of age; E. Expression of breast muscle development related genes at 35 days of ages; F. Expression of breast muscle development related genes at 63 days of ages图3 母鹅日粮添加甜菜碱对子代鹅胸肌肌纤维类型及肌肉发育相关基因的表达Fig.3 Effect of maternal betaine supplementation on breast muscle fiber type and expression of breast muscle development related genes in offspring goslings
2.5 母鹅日粮添加甜菜碱对子代鹅胸肌IGFs含量及生长相关基因表达的影响
如图4所示,35日龄时HBT组子代鹅胸肌中IGF-1含量有增加的趋势(P=0.076,图4A),而IGF-2含量在HBT组显著升高(P<0.05,图4B),同时也显著提高了两个处理组中IGF-2、IGF-1R的mRNA相对表达水平,而mTOR的mRNA相对表达水平仅在HBT组显著升高(P<0.05,图4C)。63日龄时,LBT和HBT组胸肌中IGF-1和IGF-2含量均显著高于对照组(P<0.05,图4 D、4E),与之相对应的IGF-1、IGF-2、IGF-1R的mRNA表达水平也明显升高(P<0.05,图4F),而mTOR的mRNA表达量仅在HBT组显著升高(P<0.05,图2F)。
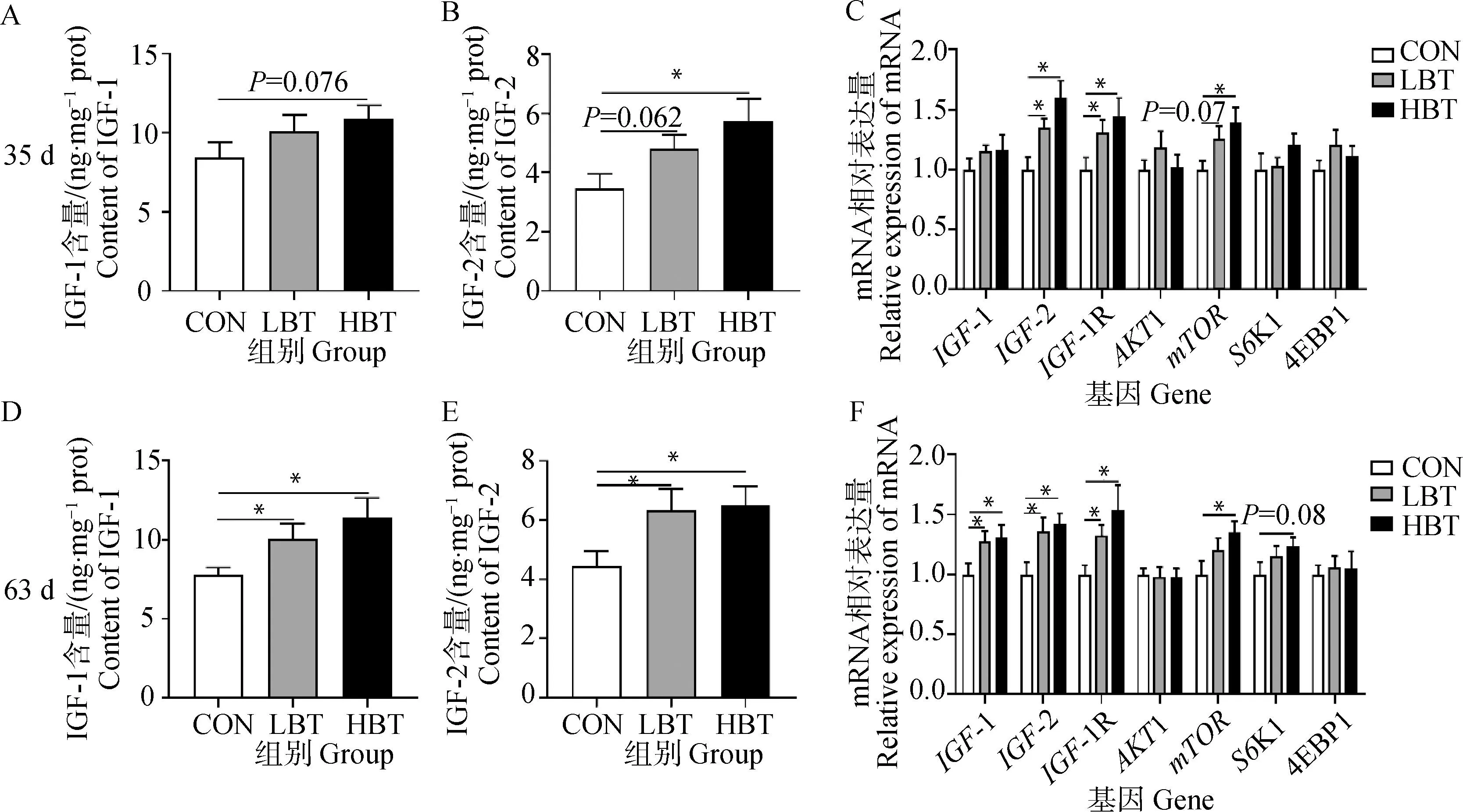
A~C. 35日龄胸肌IGF-1含量、IGF-2含量、生长相关基因的表达;D~F. 63日龄胸肌 IGF-1含量、IGF-2含量、生长相关基因的表达A-C. Contents of IGF-1, IGF-2 and growth related genes expression in breast muscle of offspring goslings at 35 days of age; D-F. Contents of IGF-1, IGF-2 and growth related genes expression in breast muscle of offspring goslings at 63 days of age图4 母鹅日粮添加甜菜碱对子代鹅胸肌IGFs含量及生长相关基因的表达Fig.4 Effect of maternal betanine supplementation on the contents of IGFs and expression of growth related genes in breast muscle of offspring goslings
2.6 母鹅日粮添加甜菜碱对子代鹅胸肌蛋白质合成的影响
胸肌重量的增加与胸肌蛋白质合成速率有关,本研究对两个日龄子代鹅胸肌中AKT、p-AKT、mTOR和p-mTOR的蛋白表达进行了检测。如图5所示,母鹅日粮添加甜菜碱对35日龄LBT和HBT组子代鹅胸肌中AKT、p-AKT的蛋白表达以及p-AKT/AKT的比值无显著影响(P>0.05,图5A),但p-mTOR蛋白表达量和p-mTOR/mTOR比值显著高于对照组(P<0.05,图5B)。此外,显著提高了63日龄子代鹅胸肌中p-AKT、p-mTOR的蛋白表达量(P<0.05,图5C、5D)以及p-AKT/AKT、p-mTOR/mTOR的比值(P<0.05,图5C、5D)。
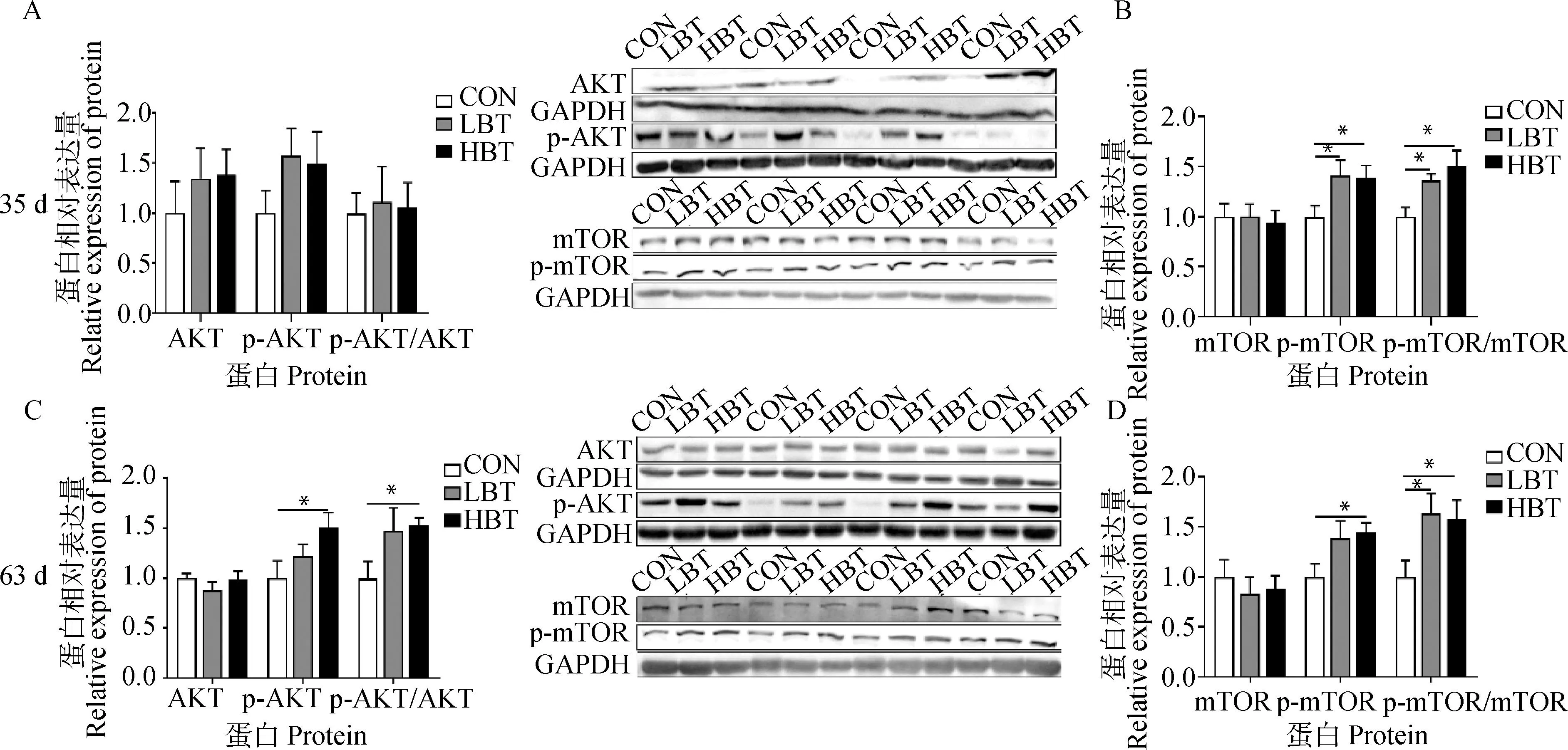
A. 35日龄AKT、p-AKT蛋白表达;B. 35日龄胸肌mTOR、p-mTOR蛋白表达;C. 63日龄胸肌AKT、p-AKT蛋白表达;D. 63日龄胸肌mTOR、p-mTOR蛋白表达A. AKT, p-AKT protein expression at 35 days of age; B. mTOR, p-mTOR protein expression at 35 days of age; C. AKT, p-AKT protein expression at 63 days of age; D. mTOR, p-mTOR protein expression at 63 days of age图5 母鹅日粮添加甜菜碱对子代鹅胸肌中AKT、p-AKT、mTOR 和p-mTOR的蛋白表达Fig.5 Effect of maternal betaine supplementation on the protein expression of AKT, p-AKT, mTOR and p-mTOR in breast muscle of offspring goslings
3 讨 论
本研究显示,在母鹅基础日粮中添加不同水平的甜菜碱,均可以显著增加其子代鹅的体重和胸肌指数,并且饲喂母鹅0.5%水平的甜菜碱还可以显著提高子代鹅63日龄的胸肌指数。前人的研究表明,在饲粮中添加0.05%、0.08%和0.1%的甜菜碱能显著提高肉仔鸡的体重[15,26]。另外,在藏鸡基础日粮中添加3 g·kg-1的甜菜碱,能显著提高子代雏鸡的出雏重,但对子代的胸肌指数和腿肌指数无显著影响[27]。杨雨等[14]研究发现,在江南白鹅饲粮中添加0.06%、0.12%、0.18%和0.24%的甜菜碱对63日龄仔鹅的胸肌率和腿肌率均没有显著影响。这些结果与母体日粮添加甜菜碱试验结果有所不同,原因可能与以往对甜菜碱的研究多集中在直接添加对家禽生产性能的作用,而本试验关注母源性甜菜碱对子代鹅生长特性的研究。母体通过沉积在蛋内的甜菜碱等活性物质影响子代胚胎发育。此外,本研究中的甜菜碱添加剂量与前人的研究也不同。
由于母源性甜菜碱组其后代仔鹅胸肌的重量显著高于对照组,本研究进一步通过组织形态学的方法对胸肌肌纤维直径和横截面积进行统计和分析。结果显示,胸肌纤维的变化趋势与胸肌重量的变化一致,表明母鹅日粮添加甜菜碱显著促进了子代鹅胸肌肌纤维肥大,且这种促进作用可持续至上市日龄。当骨骼肌中蛋白质的合成速率超过分解速率时,肌肉质量就会增加。在骨骼肌中每个细胞核与其周围的细胞浆称为一个DNA单位(RNA/DNA比值)。当肌纤维肥大时,肌纤维中每个细胞核周围的细胞浆会进一步扩大,此时DNA浓度下降,RNA/DNA比值越大,表示肌纤维肥大程度越高,蛋白质合成能力越强[28]。本试验中发现子代鹅胸肌中RNA/DNA比值以及蛋白含量显著高于对照组,在肉鸡上也发现了类似的结果[29]。此外,研究发现甜菜碱对肉鸡的促生长作用与胸肌中MyoD1 mRNA表达水平的增加有关[15],然而在本研究中,母鹅日粮添加甜菜碱并未影响子代鹅胸肌中部分生肌调节因子的表达,但显著增加了子代鹅胸肌中Pax7基因的表达,提示卫星细胞在一定程度上使子代鹅胸肌肌纤维有进一步肥大的能力。
肌肉组织以自分泌或旁分泌的方式产生IGF-1[30]和IGF-2[31]并通过IGF-1R受体激活AKT/mTOR信号通路,促进肌肉中蛋白质的合成。研究表明,生长速度快的肉鸡胸肌中IGF-1含量以及IGF-1的mRNA相对表达量明显高于生长速度慢的肉鸡[28]。在日粮中添加甜菜碱不仅能增加子代鹅血液中IGF-1的水平[14],还能通过激活IGF-1-mTOR信号通路,增加肉鸡的胸肌产量[15]。另外,母猪妊娠期饲粮中添加甲基供体通过提高仔猪骨骼肌中IGFs家族和生肌调节因子基因的表达,促进骨骼肌的分化和成熟,从而增加仔猪的体重[32]。本研究与上述结果一致,母鹅日粮添加甜菜碱显著提高了子代鹅胸肌中IGF-1和IGF-2的含量。同时,显著提高了生长相关基因IGF-1、IGF-2、IGF-1R和mTOR的mRNA相对表达量,以及p-AKT和p-mTOR的蛋白表达水平,此外,在63日龄子代鹅胸肌中,mTOR下游的关键效应底物S6K1的表达水平有增加的趋势。因此,本试验表明,母源性甜菜碱通过激活IGFs-AKT-mTOR信号通路来增强子代鹅胸肌中蛋白质的合成,从而促进胸肌生长,使子代鹅的胸肌产量增加。此外,母体的营养状态可以通过染色质表观遗传学修饰,在后代胚胎发育过程中影响子代出生后的生长发育。甜菜碱可通过一碳代谢途径提供甲基,因此可能还存在胸肌中相关基因启动子区域DNA甲基化修饰的变化及机制,这需要进一步探究。
4 结 论
综上所述,母鹅日粮中添加不同水平的甜菜碱均可增加子代鹅的体重,对胸肌的生长有促进作用,这种作用与激活IGFs-AKT-mTOR信号通路,促进子代鹅胸肌肌纤维肥大和蛋白质合成有关。因此,在实际生产中,根据生产需求,可在母鹅日粮中选择性添加0.25%或0.5%水平的甜菜碱,以提高子代鹅的生产性能,从而获得更好的经济效益。
