动物早期胚胎发育中表观重编程的机制
2024-01-10张宸艺博任斌斌郑睿智朱文治苏建民
张宸艺博,余 彤,任斌斌,郑睿智,朱文治,苏建民
(西北农林科技大学动物医学院,家畜生物学重点实验室,杨凌 712100)
最近几年,表观遗传学研究在发育生物学、再生医学等领域备受关注,特别是胚胎发育过程中的表观遗传现象。不同于有丝分裂中子细胞直接继承母细胞的表观遗传印记,在动物早期胚胎发育期间,胚胎表观基因组的重编程是一个极其重要的表观遗传学事件。重编程的目的是清除受精前雌雄配子既有的分化记忆,从而使受精后的合子获得全能性。在早期胚胎发育过程中,表观遗传重编程的关键事件包括消除附植前胚胎雌雄原核中的大部分表观遗传标记,以及在附植后建立胚胎表观遗传模式。哺乳动物胚胎表观遗传重编程过程受到多种机制的严格调控。如果该过程发生异常,常导致胚胎发育相关基因的异常表达,继而阻滞胚胎发育,甚至导致胎儿发育异常或死亡[1]。因此,了解哺乳动物胚胎重编程过程中的动态变化及其调控机制具有十分重要的意义。本文将综述近年来国内外在哺乳动物胚胎重编程方面的研究进展,以小鼠的重编程模式为主要线索,同时比较不同物种间重编程机制的差异。
1 DNA甲基化
5-甲基胞嘧啶(5-methyl-cytosine, 5-mC)是一种可逆的表观遗传标记,通过DNA甲基转移酶(DNA methyl-transferase, DNMT)将S-腺苷甲硫氨酸(SAM)提供的甲基基团转移到胞嘧啶第五个碳原子而形成,常出现在CpG岛并抑制基因的表达[2]。在哺乳动物早期胚胎中,DNA甲基化主要由DNMT3A/B和DNMT1催化,而TET酶则通过一系列反应去除胞嘧啶上的甲基基团[3]。
1.1 受精后发生的大范围去甲基化
在小鼠早期胚胎第一次卵裂之前,除了印迹控制区(imprinting control regions,ICRs)和一些逆转录转座子外,父本与母本基因组都经历了广泛的主动和被动去甲基化,其中父本基因组的去甲基化过程更加迅速。在受精卵DNA第一次复制之前,TET3会将DNMT3A和DNMT1新合成的5 mC转化为5-羟甲基胞嘧啶(5-hydroxy-methyl-cytosine, 5hmC),使得父本基因组中5hmC的含量逐渐增加,而5 mC含量急剧减少[3]。相比之下,由于DNMT1无法进入细胞核,母本基因组在DNA复制的过程中发生被动去甲基化,甲基化水平变化程度相对于父本基因组较为缓和[3-4],这种变化可能与H3K9me2招募母源因子STELLA有关[5],但一些新发现使这种机制的有效性存疑。例如,当缺少STELLA时,由DNMT1介导的从头甲基化导致小鼠受精卵中5 mC水平显著升高[6],而STELLA过表达则会导致胚胎干细胞中的DNA低甲基化[7]。因此,STELLA对母体基因组DNA甲基化的保护作用有待进一步验证。此外,在卵裂过程中,从成熟精子中遗传而来的父本基因组高CpG甲基化水平迅速下降,表明其发生主动去甲基化[4]。这一过程由DNA甲基氧化酶Tet1、Tet2或Tet3联合胸腺嘧啶DNA糖基化酶(thymine DNA glycosylase, TDG)及DNA碱基修复途径共同作用,最终去除5 mC标记[8-9]。与之相反,母本基因组初始甲基化水平较低[10],在随后的卵裂过程中,由于维持DNA甲基化的酶DNMT1表达降低,母本基因组中新合成的DNA无法被甲基化,5 mC水平随着细胞分裂而降低,即被动去甲基化[3]。这种差异化的去甲基化模式使得早期胚胎基因组呈现明显不对称的甲基化变化模式,并且在一定程度上受到H3K9me3的调节[11]。在猪的早期胚胎中,其整体的去甲基化模式呈现与小鼠相似的变化[12],而牛与人类的去甲基化模式更接近[13-14](图1)。
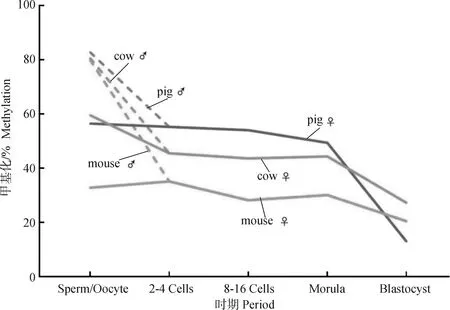
图1 不同物种从配子到囊胚的全基因组甲基化的变化(改自文献[13])Fig.1 Whole-genome methylation changes in various animals from gametes to blastocysts (adapted from[13])
1.2 甲基化的重新建立
在早期胚胎发育过程中,维持基因组的初始低甲基化状态确保了将来分化调控的准确性,同时也是维持原始多能态的必要条件。进入囊胚阶段后,DNMT3和TET酶的表达共同影响DNA甲基化的重新建立;随着DNA甲基化水平的增加,胚胎的全能性逐渐消失,并开始出现细胞分化。DNMT3A能够促进父本DNA甲基化的增加,这与它快速进入雄原核的时间一致;如果父本基因组未能正常建立甲基化,就会导致父本等位基因的表达异常[4]。在牛和猪的胚胎中,除了DNMT3L的转录物缺失外,DNMT3A、DNMT3B的mRNA以及DNMT1都有明显表达[13]。TET酶能够通过将5 mC转变为5hmC来轻微降低合子基因组的甲基化水平,但是如果缺乏TET酶,胚胎发育就会停滞在2细胞阶段[15]。在猪中,TET3主要存在于卵母细胞和第一卵裂阶段,并逐渐被TET1和TET2替代,这与小鼠胚胎发育中的情况相似[13]。此外,在小鼠中,缺失DNMT1会导致多能性标记无法被正常抑制,而3种TET酶均缺失的细胞表现出造血干细胞调控缺陷[16],这表明DNA甲基化的动态变化可能影响早期细胞命运决定。
显而易见的是,DNA甲基化重编程异常可以导致胚胎发育障碍甚至死亡。例如,在体细胞核移植(somatic cell nuclear transfer, SCNT)的过程中,由于现有技术手段难以完全去除供体细胞核中原有的表观遗传修饰,克隆胚胎的死亡率通常高于体内受精胚胎,其中一个重要的原因是克隆胚胎中的DNA甲基化长时间维持在较高水平[17-18](图2)。因此,研究SCNT胚胎中异常的DNA甲基化模式及其纠正措施,将有助于加深对于自然受精胚胎基因组甲基化重编程机制的理解。

图中展示了哺乳动物个体发育周期中全基因组DNA去甲基化和再甲基化的两个波。受精后不久,从配子遗传的甲基化标记被清除,父系基因组经历主动去甲基化,母系基因组经历被动去甲基化。附植后,从头甲基化建立了最初的胚胎甲基化模式The graph shows the two waves of whole-genome DNA demethylation and re-methylation during the mammalian ontogeny cycle. Shortly after fertilization,methylation markers inherited from gametes are eliminated with the paternal and maternal genome undergoing active and passive demethylation, respectively. After implantation, the initial embryo methylation pattern was established by de novo methylation图2 哺乳动物发育过程中DNA甲基化的动态变化(引自文献[19])Fig.2 Dynamic changes of DNA methylation during mammalian development (quoted from[19])
2 组蛋白修饰
核小体是染色体结构与功能的基本单位,由一个组蛋白八聚体、连接组蛋白H1和约150 bp的DNA构成。组蛋白八聚体由H2A、H2B、H3和H4各两个分子组成,其N末端突出于核小体外,可发生多种蛋白质翻译后修饰,包括甲基化、乙酰化、磷酸化、泛素化、类泛素蛋白化、ADP-核糖基化以及生物素化等,本文将主要讨论在胚胎发育过程中最为重要的两种修饰:组蛋白甲基化和乙酰化。这些修饰可以影响转录因子与染色质的结合,进而调控发育相关基因的表达。在附植前胚胎的发育过程中,父系和母系遗传的组蛋白修饰会经历不同的重编程过程:父源组蛋白修饰通常在受精卵DNA复制之前被快速清除,而母源组蛋白修饰则有部分通过母源遗传而得以保留。
2.1 H3K4me3
H3K4me3是组蛋白第三亚基四号赖氨酸的三甲基化,主要由混合系白血病(mixed lineage leukeMia, MLL)蛋白家族的组蛋白赖氨酸甲基转移酶(histone methyltransferase,HKMTs)催化,包括MLL1~MLL4、SET1A和SET1B[20-23],能够促进染色质维持松散状态,从而活化发育相关基因的启动子。精卵结合后,小鼠父本基因组中的H3K4me3水平迅速降低,在合子基因组激活(zygotic genome activation, ZGA)期间逐渐恢复,并在胚胎植入后达到与母本基因组相似的水平[5];而在小鼠的母本基因组中,启动子和远端位点存在大量非典型H3K4me3(noncanonical H3K4me3, ncH3K4me3)[24-25]。这些非典型标记与成熟卵母细胞中部分DNA甲基化区域重叠[26];直到受精卵的ZGA主波阶段,ncH3K4me3才被典型H3K4me3替换,并在4细胞时期基本消失[24]。ncH3K4me3水平的变化似乎仅出现在卵母细胞和早期胚胎中,其调控可能与多种组蛋白去甲基化酶(lysine(K)demethylase, KDM)有关。例如,KDM5A和KDM5B过表达会激活成熟卵母细胞的转录组,而KDM5A敲降会导致胚胎发育异常[27]。近期另一项研究指出,牛早期胚胎中组蛋白去甲基化酶KDM5A、KDM5B和KDM5C的缺失均会增加H3K4me3水平,从而降低8/16细胞期胚胎的发育率[28]。在猪附植前胚胎中,KDM5B表达与H3K4me3去甲基化有关,其敲低不仅会增加H3K4me3水平,还会破坏囊胚期H3K4me3与H3K27 me3的平衡,这表明KDM5B对早期胚胎H3K27 me3重编程至关重要[29]。上述研究表明,ncH3K4me3可能引起胚胎基因组沉默,但在猪SCNT胚胎中,存在过量H3K4me3的位点似乎并未出现基因表达异常,表明猪可能存在不同的机制维持这些位点的转录[9]。此外,在小鼠的父本基因组中还存在一类富集于拓扑关联域(topologically associating domains,TADs)的稳定的H3K4me3,其与ZGA期间胚胎转录激活有关。考虑到CCCTC结合因子(CCCTC-binding factor, CTCF)在染色质高级结构的形成中具有重要作用[30],这类稳定的H3K4me3可能通过对TADs中的CTCF结合位点提供正向调控,进一步促进染色质高级结构的形成,从而促进胚胎的转录激活[31]。
在2细胞阶段,典型H3K4me3在启动子区建立[31],启动子区广泛的H3K4me3标记可能有助于关键细胞类型特异性因子的稳定转录[32]。这一过程的调控机制可能涉及多种信号分子。最近的研究显示,在小鼠胚胎干细胞中,Dppa3敲降会导致H3K4me3水平下降,并对异染色质标记物(如H3K9me3和H3K27 me3)也产生负调节作用,表明Dppa3可能参与促进包括H3K4me3在内的表观遗传重塑[7]。另外,由于DNMT的功能与SAM密切相关,SAM由甲硫氨酸经甲硫氨酸腺苷转移酶(methionine adenosyltransferase, MAT)催化生成[33],因此MAT可能间接影响组蛋白甲基化水平。在哺乳动物中,MAT存在3种不同的表达形式,分别是MAT1A、MAT2A和MAT2B[34]。在小鼠胚胎中敲降MAT2A并未显著改变H3K4me3和H3K9me3水平,尽管这会使胚胎发育停滞在2细胞期[35],这可能与受精卵胞质中的母源性SAM尚未完全消耗有关。此外,共激活因子相关的精氨酸甲基转移酶1(coactivator-associated arginine methyltransferase 1, CARM1)可以催化H3第26位精氨酸残基的对称性二甲基化(H3R26 me2s)[36],当CARM1被抑制时,小鼠附植前胚胎中的H3K4me3水平降低[37],牛和猪的早期胚胎也显示出类似结果[38-39],这可能是由于CARM1影响了核组织结构,并且对染色质结构有重要作用[40]。WD重复蛋白5(WD repeat-containing protein 5, WDR5)作为MLL/SET甲基转移酶复合物的中心成分,在猪的早期胚胎发育过程中起到重要作用。WDR5敲降会增加H3K4me3水平,并导致囊胚质量下降和胚胎发育潜力减弱[41]。虽然MLL3可能介导了WDR5蛋白对H3K4me3的调节作用,但是MLL3缺失并不影响小鼠早期胚胎中H3K4me3水平或HKMTs活性,这表明WDR5与催化H3K4me3相关的HKMTs之间的关系仍需进一步阐释[42]。
2.2 H3K27me3
H3K27me3是一种由多梳抑制复合体2(polycombrepressive complex 2,PRC2)催化的甲基化修饰,它主要富集于母本基因组上,能够抑制编码多能性因子、发育因子和谱系特异性转录因子等基因的表达[43-44]。例如,在哺乳动物克隆胚胎中,H3K27me3异常高表达导致重编程不完全和胚胎发育障碍。小分子抑制剂GSK126能够特异性抑制EZH2的甲基化功能,另一个小分子抑制剂GSK-J4能够特异性抑制KDM6B,有研究表明,经过低浓度(0.1 μmol)GSK126抑制H3K27甲基化酶处理后,猪的克隆胚胎中H3K27me3水平显著降低,同时囊胚率也有所提高,而相同浓度GSK-J4抑制去甲基化酶处理的猪胚呈现相反的变化[45]。卵母细胞中PRC1介导的组蛋白H2A赖氨酸119位点单泛素化(H2AK119ub1)修饰也参与了H3K27me3在受精卵中的遗传[46]。在小鼠早期胚胎发育过程中,遗传自配子的H3K27me3早在PN5时期就出现广泛的丢失,直到植入后才重新建立[47]。这一过程涉及父本基因组中H3K27me3的整体去除和母本基因组上启动子区域H3K27me3的选择性去除[5]。而在猪早期胚胎发育过程中,从1细胞期到桑椹胚阶段,H3K27me3丰度逐步下降至最低水平,但在孵化后的囊胚中重新增加[48]。有研究指出,在小鼠胚胎单细胞期后,启动子区域富集了H2AK119ub1修饰,并可能促进了多梳结构域的形成和H3K27me3的重新建立[49,46]。然而,在去除H2AK119ub1后,4细胞期前并未出现H3K27me3的明显减少[50],表明H3K27me3和H2AK119ub1在小鼠早期胚胎重编程中的相互作用值得进一步研究。此外,H3K27me3的水平与组蛋白甲基化转移酶PRC2和组蛋白去甲基化酶KDM6A的动态变化相一致[51-52]。例如,在牛和猪的SCNT胚胎中,过表达KDM6A可以降低过高的H3K27me3水平从而提高SCNT胚胎的重编程效率[53]。在猪的胚胎中,过表达KDM4A有利于降低SCNT胚胎的H3K27me3水平,而KDM5B的缺失也可能通过影响KDM6A的表达而降低H3K27me3的丰度[9,29]。总之,虽然H3K27me3及其相关酶在胚胎重编程中的具体作用机制还不清楚,但已有研究证明了H3K27me3对于维持和改变染色质状态的重要性。
2.3 H3K9me3
H3K9me3修饰在小鼠植入前胚胎中调控重复序列元件和部分蛋白编码基因的表达。与H3K4me3和H3K27me3不同的是,H3K9me3主要富集于早期胚胎中的LTR,这表明其可能参与LTR的调节[54]。在2细胞期,双亲基因组中从头建立的H3K9me3结构域显著增多,但在LTR与启动子上呈现出不同模式:LTR上以H3K9me3标记的区域逐渐增多并维持在较高水平,而启动子上的H3K9me3结构域则在受精后减少,在植入后恢复,表明不同位点上存在不同的机制调控H3K9me3的重编程[54]。此外,在牛的受精卵中,亲本基因组上的H3K9me3呈不对称分布,并且在雌原核上水平更高[55]。这说明亲代之间不对称的组蛋白信号既可能源于配子的表观遗传,也可能受合子阶段的重编程影响。
H3K9me3通常被认为是细胞命运变化的障碍之一,因为它可以阻止转录因子Oct4、Sox2、Klf4、c-Myc与胚胎干细胞中一些涉及多能性的染色质区域结合,并且抑制附植前胚胎中富含CpG的基因组位点发生去甲基化,从而阻碍正常的重编程进程[11,56-57]。H3K9me2/3由组蛋白甲基转移酶GLP、G9a催化形成,与异染色质蛋白1(heterochro matin protein 1, HP1)的染色质结构域结合,从而促进胚胎干细胞中异染色质的形成。而在构建克隆胚胎时,H3K9me3重编程不完全会导致早期克隆胚胎囊胚发育率降低[58-59]。因此,调控H3K9me3修饰水平对于提高克隆效率具有重要意义。研究表明,在小鼠和猪等物种中,过表达KDM4d、KDM4e或敲降供体细胞中的Suv39h1/h2(H3K9me3甲基转移酶)均可降低H3K9me3对发育基因转录的抑制[59-62]。本课题组前期研究发现,在牛的早期胚胎中,microRNA-125b通过直接结合SUV39H1,从而抑制其表达产物介导的H3K9me3生成[63];在羊早期克隆胚胎中,注射人源重组KDM4d蛋白也可以降低H3K9的甲基化水平[64]。但在猪早期克隆胚胎中,KDM4d的过表达并不能改变高H3K9me3修饰状态[62],而过表达KDM4A则能显著增加猪早期克隆胚胎的囊胚率[62,65]。敲除KDM4A会导致H3K4me3所属位点被异常的H3K9me3入侵,使得大多数修饰的基因表达下调,表明KDM4A对H3K9me3的去甲基化是确保合子基因在ZGA期间正确激活的必要机制[66]。另一方面,尽管过表达KDM4b有利于提高克隆效率[61],但注射过高剂量的KDM4b mRNA会抑制滋养外胚层(trophectoderm, TE)的分化[67],这表明H3K9me3在适度水平上的维持对于保障胚胎的正常发育至关重要[5]。值得注意的是,有研究指出,在附植前胚胎中,由SUV39H2催化从头合成的H3K9me3不具有调控整体基因表达水平的作用,且为非抑制性标记;随着卵裂的进行,亲本基因组间的转录活性差异逐渐消失,表明早期的H3K9me3标记在发育后期倾向于产生抑制作用[68](图3)。
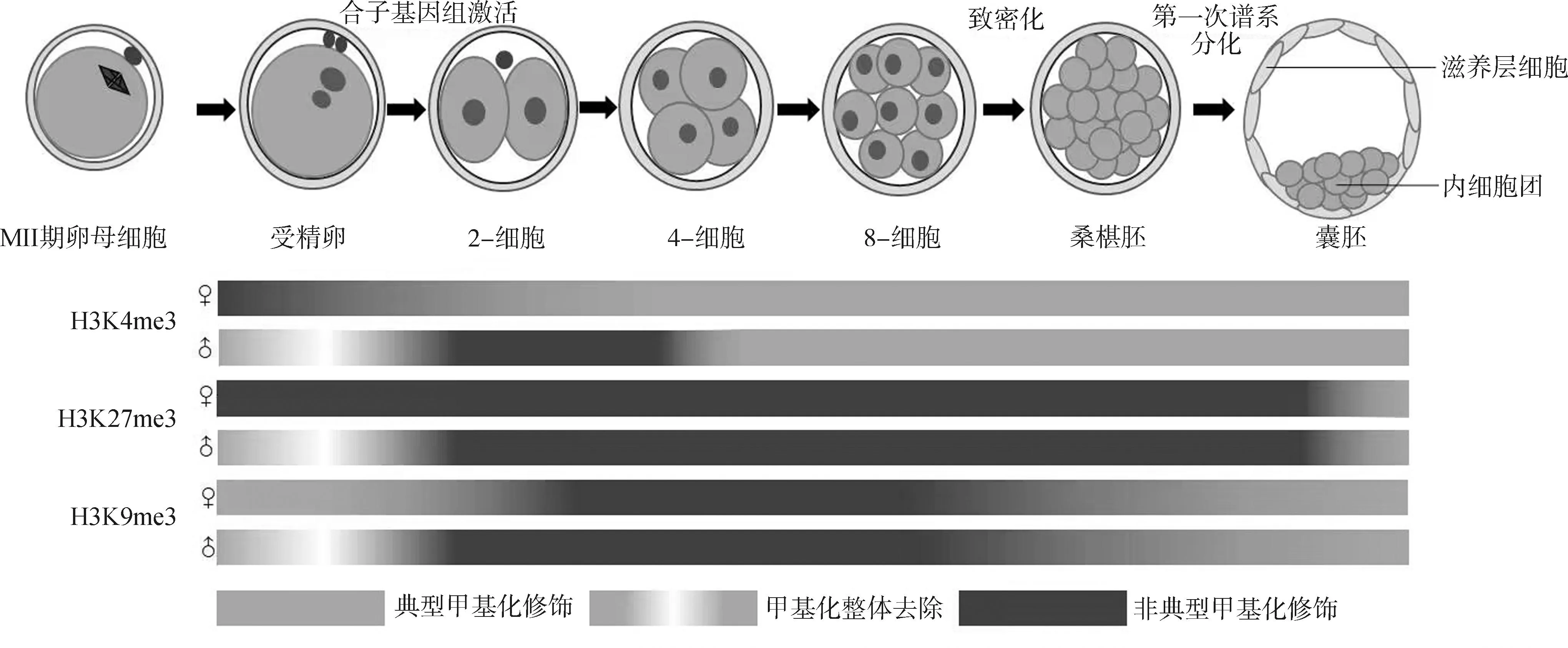
小鼠胚胎中全基因组沉默从配子发育晚期开始,一直持续到ZGA时期。卵母细胞中ncH3K4me3、H3K27me3与H3K9me3的广泛结构域可遗传至胚胎,其中ncH3K4me3在ZGA之前被H3K4从头甲基化取代,H3K27me3可维持至囊胚期并最终被重编程为其典型模式,H3K9me3则被重编程为非典型形式并一直持续到囊胚期。父源性组蛋白修饰H3K4me3、H3K27me3与H3K9me3则在2细胞期前被从头甲基化的广泛结构域取代,其中H3K4me3在ZGA前被重编程为典型模式,H3K27与H3K9的甲基化结构域则可以维持到囊胚,并在附植后重编程为典型模式In mouse eggs, genome-wide silencing begins during late gametogenesis and lasts until ZGA. Broad domains of maternal ncH3K4me3, H3K27me3 and H3K9me3 are inherited to the embryo, where ncH3K4me3 is replaced by H3K4 de novo methylation before ZGA, H3K27me3 persists until blastocyst and is eventually reprogrammed to its canonical pattern, and H3K9me3 is reprogrammed to a noncanonical form and maintains until blastocyst. Paternal histone modifications H3K4me3, H3K27me3 and H3K9me3 are replaced by broad domains of de novo methylation before 2-cell stage, where H3K4me3 is reprogrammed to a canonical pattern before ZGA, while H3K27 and H3K9 methylation domains endure until blastocyst and are reprogrammed to canonical patterns after implantation图3 小鼠早期胚胎发育过程中的H3组蛋白甲基化变化(改自文献[69])Fig.3 Changes in H3 histone methylation during early mouse embryo development (adapted from[69])
2.4 组蛋白乙酰化
组蛋白乙酰化是一种发生在组蛋白N端赖氨酸残基的ε-氨基上的表观遗传修饰,主要集中在H3和H4组蛋白上[70]。组蛋白乙酰化在基因转录、DNA复制及细胞周期调控等方面发挥着重要作用。通常情况下,组蛋白乙酰化如H3K122ac和H3K64ac与转录激活有关,而组蛋白去乙酰化与基因沉默相关[71]。根据Ziegler-Birling等[72]的研究,在附植前胚胎发育过程中,3个关键的组蛋白乙酰化位点(H3K56、H3K64、H3K122)均有表达,但是表现出不同的重编程模式。其中,H3K56ac和H3K64ac在附植前便富集于核心核小体中,而H3K122ac虽然在成熟卵母细胞中存在,但水平极低,这可能与增强子使用或调节频率较低有关。
组蛋白乙酰化与去乙酰化分别由两种作用相反的酶进行调控,即组蛋白乙酰化转移酶(histone acetyltransferases, HATs)和组蛋白去乙酰化酶(histone deacetylases, HDACs)。HATs通过乙酰辅酶将一个乙酰基连接到赖氨酸侧链的ε-氨基上,从而中和了赖氨酸的正电荷,降低了组蛋白和DNA结合的紧密程度,使染色质结构松弛,便于转录因子结合。与之相反,HDACs去除组蛋白尾部乙酰化修饰后,赖氨酸的正电荷恢复,使得染色质结构重新稳定[73]。在小鼠受精卵中,H3K27ac等组蛋白乙酰化位点在雌雄原核中大量存在,其信号在ZGA主波发生前急剧减弱。这种过程在牛、猪等多个物种的早期胚胎中均有发生,表明组蛋白乙酰化对于哺乳动物早期胚胎的正常发育具有重要意义[74-77]。在小鼠和牛的早期胚胎中,HDAC1与HDAC2是高度同源且表达量最高的两种组蛋白去乙酰化酶。尽管相比于HDAC1对附植前胚胎的重要作用而言,HDAC2主要调控卵母细胞的正常发育,但二者对早期胚胎发育的调控呈现出互补作用[74,78]。单独敲降HDAC1或HDAC2均不会对小鼠附植前胚胎发育产生明显影响,但当两者均被敲降后,小鼠胚胎发育停滞在桑椹胚期。这可能是由于高乙酰化水平导致TRP53上调、TE与内细胞团(inner cell mass, ICM)谱系特异性分化受阻,以及全基因组DNA甲基化水平升高的共同作用所致[79]。此外,HDACs还可以将广泛分布的H3K27ac还原成典型分布模式,从而通过阻止发育基因过早表达确保ZGA的正常进行[80]。综上所述,在囊胚前阶段,通过直接或间接地调控胚胎干细胞谱系特异性分化、DNA甲基化等过程,组蛋白乙酰化模式对于维持正常的胚胎发育至关重要。
3 染色质开放性
染色质开放性(chromatin accessibility)又称为染色质可及性。在真核细胞的细胞核中,DNA与组蛋白结合形成核小体,并经过进一步折叠在细胞核中形成高度压缩的染色质,这对DNA的复制与表达有一定的限制作用。开放的染色质在早期胚胎发育过程中发挥重要的调控作用,不仅影响ZGA的进程,还参与细胞命运决定与谱系特异性分化。
早期胚胎的发育过程伴随着染色质开放性的显著变化,不同物种的早期胚胎也呈现出不同的染色质可及性重编程模式。例如,小鼠受精卵中较高的染色质可及性在1细胞期末期降低,然后在2~4细胞期重新升高[16]。在牛的早期胚胎中,2~4细胞期胚胎的染色质可及性较低,但在8细胞期明显增高[81]。精卵结合后,父本基因组上包裹的鱼精蛋白被置换为组蛋白,雄原核的染色质相对于雌原核更加紧密,呈现出不对称的染色质开放模式[82]。随后父本染色质被迅速重编程,在PN3期与母本基因组的染色质开放性接近。这个过程与亲本基因组之间DNA甲基化和组蛋白修饰的重编程模式非常不同[83],这种差异的产生可能与不同表观遗传方式的来源有关。在小鼠2细胞期自然受精胚胎中,转录因子Nfya促进了ZGA的发生,以及开放染色质在活跃转录基因启动子和转录起始位点的建立[82,84];同时逆转录转座子LINE1的表达也高度上调,过早敲降或长时间激活LINE1都会对染色质可及性的正常形成产生不利影响[85]。在8细胞期染色质开放性变化最为剧烈,此时Oct4表达也高度上调,表明Oct4可能促进染色质开放[82]。除此以外,还有多种因素被认为参与早期胚胎松弛染色质结构的形成,包括转录因子DUX、染色质组装因子CAF-1等[86-88]。这些研究揭示了复杂的染色质开放性调控网络,但其潜在的调控机制以及各种因素间的相互关系仍需进一步研究。
除了转录因子外,表观遗传修饰也被证明与染色质开放性有关。一般来说,去甲基化或具有中等程度羟甲基化水平的DNA以及乙酰化的组蛋白通常具有更高的可及性[84,89]。在小鼠原始生殖细胞中,H3K4me3标记的区域比H3K27ac位点的染色质具有更高的开放性,而H3K27me3与染色质开放性关联较弱[84]。在牛早期胚胎中,microRNA-125b过表达导致H3K9me3水平降低,并伴随着异染色质的松弛[63]。上述研究表明,在哺乳动物早期胚胎中,DNA甲基化和组蛋白修饰协同参与染色质开放性重编程的正常进行。
4 染色质三维结构
真核细胞中,染色质形成复杂而有序的三维结构,包括染色质疆域(chromosome territories)、区室(compartments)、拓扑关联域(topologically associating domains,TADs)、染色质环等层级[90-91]。染色质疆域是指不同的染色质在核内折叠并占据的特定区域;区室是指具有相似基因表达活性的染色质区域,分为活性区室和惰性区室;TADs是指染色质上一段线性连续且内部相互作用更为频繁的区域;染色质环是指由CTCF和cohesin蛋白介导的DNA上的环状结构,通常包含调控元件如启动子、增强子和沉默子等[91]。这些染色质三维结构在受精后也经历了显著的重编程。精子基因组中与体细胞相似的TADs和区室在鱼精蛋白被置换后消失,卵母细胞基因组中原先存在的三维结构也在卵泡发育的过程中逐渐均一化,受精后两个亲本基因组都呈现相似的松散状态[92-94]。随着小鼠早期胚胎发育,受精卵中分散的染色质逐渐形成且聚集成致密的区块,并在核膜附近堆积,在8细胞期出现了较为典型的区室和TAD结构[94-95]。值得注意的是,父本基因组中的染色质区室更加明显,而母本基因组上则几乎不存在,表明染色质高级结构重编程具有等位基因特异性,这与DNA甲基化和组蛋白修饰类似[92-93]。在猪的早期胚胎中,拓扑关联域等三维结构以与小鼠相似的模式在附植前建立[96]。综上所述,受精后染色质结构高度松弛,之后随着早期胚胎发育而缓慢地重新变得紧密,但是诱导染色质折叠的因素有待更深入研究。
与染色质开放性类似,早期胚胎染色质三维结构的重编程与表观遗传修饰的重编程密切相关。例如,基因组整体低甲基化水平的状态与染色质结构重塑在时间上重合,且活性区室中DNA去甲基化速率大于惰性区室[10,92]。同时,非典型的组蛋白标记也逐渐被典型标记替代[97]。这些现象表明,表观遗传修饰可能是调控染色质三维结构重编程的重要因素。但是,这种调控是否必需?是否有特定的分子机制将这两种过程耦合起来?这些问题有待进一步研究。
5 展 望
在哺乳动物早期胚胎的发育过程中,亲本基因组经历了多种表观修饰的重编程,包括DNA去甲基化、组蛋白修饰重塑、染色质开放性增加以及染色质高级结构的变化。近期的研究揭示了这些重编程过程的潜在调控机制,然而许多这样的机制有待进一步研究与验证。早期哺乳动物胚胎发育中与全能性转变和细胞命运决定之间相互作用的分子细节,将是未来研究的一个重要方向。
早期胚胎发育过程中的表观遗传重编程是一个精细调控的过程。随着基因组学技术的不断深入发展,研究者对附植前发育的表观遗传重编程景观的理解有了很大的提高。然而,重编程是如何在不同的基因组位点上被调控的仍然是未知的。对这些因素和潜在机制的识别将提高人们对细胞命运转变和哺乳动物早期胚胎发育的理解。尽管基因组学技术的发展提高了人们对表观遗传重编程景观的认识,但是不同基因组位点上重编程的调控机制仍不清楚。未来研究需要采用多组学方法来阐明表观遗传重编程与全能性转变和细胞命运决定之间的分子联系。
表观遗传重塑的机制在不同物种中遵循相似的模式,但在一些细节上有所不同。揭示不同哺乳动物早期胚胎发育的表观遗传调控机制将是一个重要的研究领域,其结果将有助于体细胞核移植技术的发展,进而提高良种畜禽种质资源的利用效率。遗憾的是,目前大多数研究集中在小鼠与人类的早期胚胎,而其它家畜早期胚胎重编程的研究则相对较少。随着单细胞多组学技术的发展,以及单细胞转录组、染色质组图谱的绘制,表观遗传研究已逐渐深入到单细胞水平,这有助于更微观地解析其作用机理[98]。这些突破也可能为附植前胚胎发育的研究提供新的见解。
