某化工园区综合废水处理单元中病毒群落的分布特征
2024-01-08梅红汪炎洪文清宋立岩
梅红, 汪炎, 洪文清, 宋立岩
(1.东华工程科技股份有限公司, 合肥 230024; 2.安徽大学 资源与环境工程学院, 合肥 230601)
污水处理厂(WWTP)一般联合应用物理、 化学和生物等多种处理方法有效去除污水中污染物。 在污染物分解过程中, 微生物进行生物降解从而有效去除污染物[1-2]。 病毒是一种专门寄生在细胞内的微生物, 它依靠宿主微生物来生存和复制[3]。 在感染周期中, 病毒通过裂解微生物细胞来释放宿主提供的营养物质, 以及通过转导和表达病毒基因,重新编程被感染宿主的代谢过程, 从而完成自身的繁殖[4-5]。 同时, 细菌群落作为病毒寄生最多的宿主微生物, 其活动和功能受到病毒的直接调控[4]。环境中病毒的数量通常比微生物数量多10 倍以上,并通过侵染微生物对其群落结果和功能产生直接影响, 间接地对污水处理效果产生显著影响[6-7]。
随着宏基因组技术的快速发展, 人们对于病毒群落的组成和多样性有了更深入的理解[8]。 同时,生物信息分析可以进一步解析病毒群落在环境介质中的演化, 例如线性判别LEfSe 分析可以深入分析不同组间显著差异的群落特征, 并且已被用作揭示河流、 湖泊和海洋等不同环境中差异微生物群落的工具[9-11]。 这些研究揭示了不同生境中代表性微生物并阐明了差异微生物的主要特征和功能。 然而,在WWTP 各子系统中差异病毒群落发挥何种功能仍鲜为人知。
病毒通过侵染宿主微生物从而对宿主微生物群落结构和功能产生直接影响, 并间接对污水处理效果产生显著影响, 因此, 研究WWTP 中病毒群落的特征, 及其与宿主的关联可以为提升WWTP 的污水处理效率提供科学依据。 本文以安徽省某化工园区综合废水处理核心单元为研究对象, 采用宏基因组测序和LEfSe 分析, 研究了该WWTP 中原水、厌氧池和污泥池中病毒群落的分布特征及其与细菌群落的相互关系, 阐明WWTP 各子系统中病毒群落分布格局, WWTP 中细菌群落多样性与病毒群落组成间的关系, 以及WWTP 各子系统中病毒群落的差异, 旨在通过调查研究结合生物信息学分析, 揭示WWTP 中病毒群落的分布特征, 明晰宿主(细菌)对病毒群落组成的重要影响作用。 弄清WWTP 中病毒群落分布格局及其发生机制, 这不仅有助于阐明病毒在污水中的分布和迁移规律, 拓展环境微生物学研究范畴, 而且对我国水资源的可持续发展具有重要的指导意义。
1 材料与方法
1.1 采样点信息和样品采集
试验研究在安徽省某化工园区综合废水处理厂(WWTP)内进行, 该WWTP 的日处理水量为1.5 万t。 WWTP 的工艺流程如图1 所示, 废水经澄清池混凝沉淀后进入缓冲池, 出水进入水解池使其可生化性得以改善, 然后经厌氧池厌氧处理后流入好氧生物池进行好氧处理, 最后经过二沉池沉淀后排放。

图1 WWTP 工艺流程Fig.1 Process flow of WWTP
从WWTP 的进水处(RW)、 厌氧池(IAT)和回流的活性污泥(AS)中分别平行采集了3 份500 mL样品置于灭菌的超净HDPE 瓶中, 样品置于冰上运回实验室, 并在8 h 内完成预处理, 采用0.22 μm无菌亲水膜过滤水样, 同时以5 000 r/min 离心污泥样品20 min 并取0.1 g 浓缩污泥。 预处理后的样品用于DNA 提取。
1.2 理化指标的测定
使用YSI 6600 V2 多参数水质分析仪测量水样的pH 值, 按标准方法测定水样的COD、 NH3-N、TN 和TP[12]。
1.3 DNA 提取与宏基因组测序
使用FastDNA Spin Kit(MP Biomedicals, Irvine,CA)提取DNA, 并用NanoDrop 2000(Thermo Fisher Scientific, USA)定量, 然后用1% 琼脂糖凝胶电泳检测[13]。 利用Covaris M220 将DNA 片段化到约400 bp, 最后利用NEXTFLEXTMRapid DNA-Seq Kit 构建PE 文库。 配对端测序在上海某公司的Illumina HiSeq4000 平台上进行。
1.4 宏基因组序列数据的处理
首先对原始序列进行拆分、 质量剪切以及去除污染等优化处理, 质控在Fastp v0.20.0 上进行; 然后使用优化序列进行拼接组装和基因预测, 对得到的基因进行物种和功能上的注释以及分类, 注释包括基因物种注释NR。 病毒的丰度以NR 物种注释后RPKM 标准化后的绝对丰度表示, 即每百万条序列中, 每个基因以1 000 个碱基为单位, 比对上的reads 条数。
1.5 统计分析
基于r 3.6.3 和RStudio 1.1.463 软件, 并利用Spearman 相关性分析检验细菌群落丰富度和多样性与病毒群落相异度的相关性。 通过线性判别(LEfSe)分析甄别了各样点间的差异病毒, 同时基于非参数Kruskal-Wallis 秩和检验与Wilcoxon 秩和检验判别了差异病毒的一致性。 在LEfSe 分析中,选定log10(LDA)的值为3.6。
2 结果与讨论
2.1 病毒的总体消减特征
COD、 TN 和NH3-N 的浓度 在 经 过WWTP 的子系统运行后得到了有效控制, 各子系统中污染物浓度存在显著性差异(P <0.05), 同时由于TP 进水浓度较低, 故浓度变化并不规律(表1)。

表1 WWTP 研究单元的水质指标Tab.1 Water quality indicators in research unit of WWTP
WWTP 中各流程病毒丰度如下: 原水124 680±8 112、 厌氧池87 585±11 928、 活性污泥136 988±15 514。 当废水经水解和厌氧处理后, 厌氧池中病毒浓度相较于进水处下降了29.75%(以均值计算),表明厌氧处理对病毒的去除有一定作用; 污泥池中病毒浓度相较进水处上升了9.87%, 这是因为活性污泥富集了大量微生物成为病毒适宜的宿主。
2.2 病毒群落分布特征
基于宏基因组技术, 本研究在病毒门水平上展示了各样点中相对丰度为1% 以上的病毒(图2),通过热图展示了相对丰度前50 的病毒属(图3)。WWTP 中主要病毒门包含Uroviricota 和Nucleocytoviricota 门, 其在各样点平均占比为53.42% 和26.10%, 其中Uroviricota 门在活性污泥的样本中占比68.98%, 而Nucleocytoviricota 门在原水中占比52.59%。 Uroviricota 门可能会影响人体的新陈代谢或免疫系统[14], 而Nucleocytoviricota 门由于其特殊的构造而只能侵染环境中的真核生物[15], 同时真核生物会分泌大量干扰素应对病毒的侵染, 但干扰素会对人类的生命健康产生多重影响和作用[16-17]。
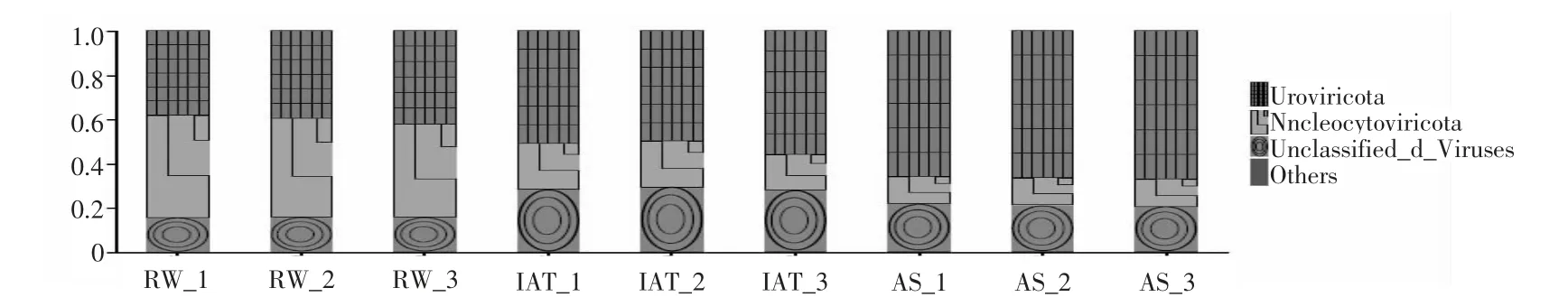
图2 病毒群落在门水平上的组成Fig.2 Viral community composition at phylum level
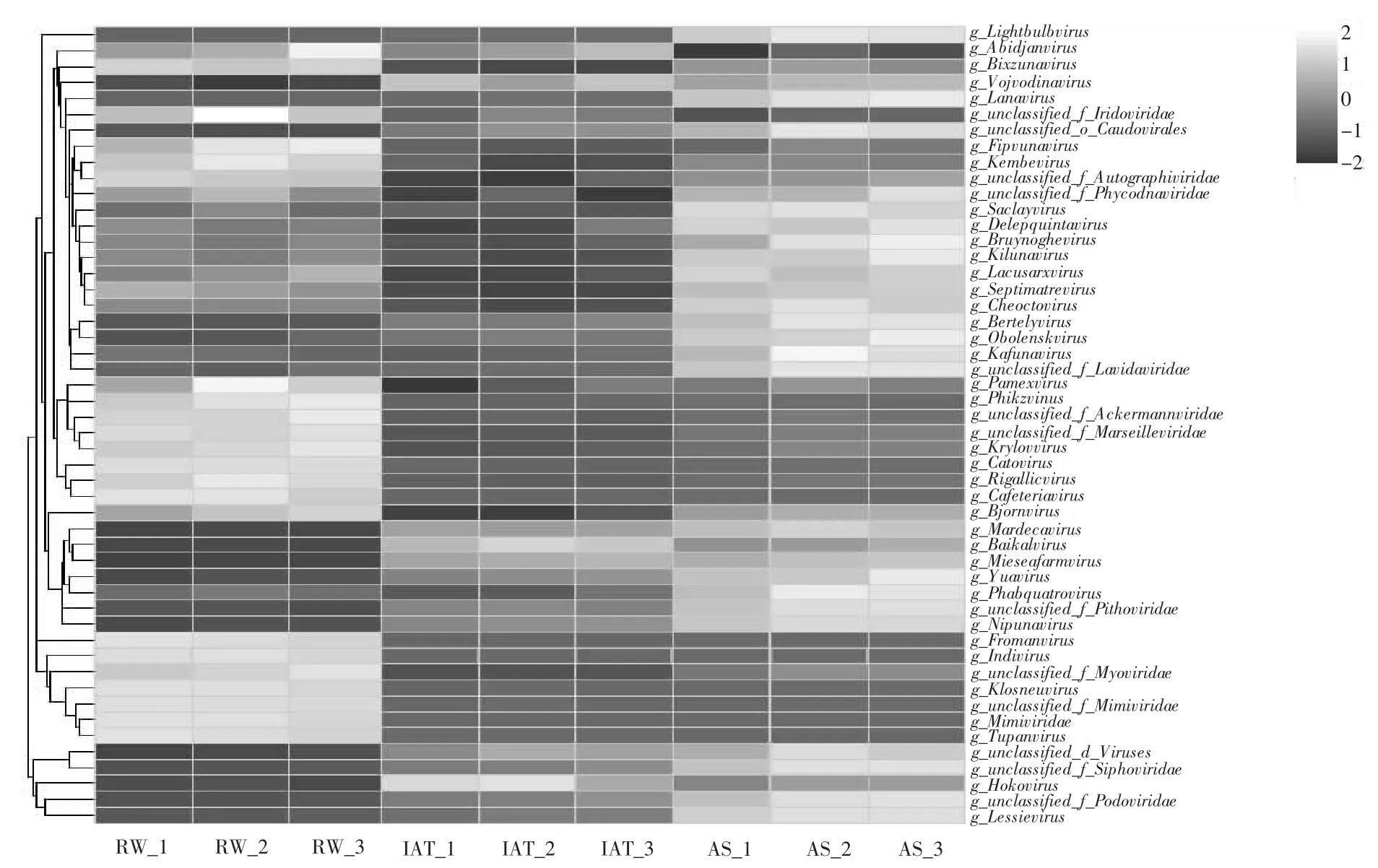
图3 病毒群落在属水平上的组成Fig.3 Composition of viral communities on genus level
在病毒的属水平上, 各病毒在不同样点中的分布存在差异。 具体而言, 原水中的Kembevirus、Krylovvirus 和Phikzvirus 属占比较高, 且在后续流程中相对丰度明显下降; 厌氧池中相对丰度较高的病毒并不多, 仅Hokovirus 属最显著, 这可能是因为厌氧环境中产生能量的途径急剧减少, 限制了病毒增殖和感染宿主[18], 同时这解释了厌氧池中病毒丰度下降了29.75%, 而在活性污泥中Phabquatrovirus、Lightbulbvirus、 Obolenskvirus 和Lanavirus 等病毒属得到了富集(图3)。 研究结果表明活性污泥中微生物浓度较高, 改变了物种间的竞争关系, 导致病毒的寄主细胞数量增加, 从而促进了病毒的增殖。
2.3 病毒在各单元中的差异
通过LEfSe 分析对比原水、 厌氧池和活性污泥三者中的差异病毒, 并将log10(LDA)的值设置为3.6, 结果表明, Megaviricetes 纲、 Mimiviridae 科、Myoviridae 科和Mimivirus 属是原水中差异显著的病毒。 Hokovirus 属、 Caudoviricetes 纲、 Caudovirales目和Siphoviridae 科是活性污泥中差异显著的病毒(图4)。 原水中多数差异病毒均与肺炎病毒相关,以往研究认为Mimiviridae 科、 Myoviridae 科和Mimivirus 属可能会引起严重的肺炎[19]。 相对而言,活性污泥中的差异病毒繁多, 如Hokovirus 属是一种猪病毒, 而Siphoviridae 科是感染金黄色葡萄球菌的病毒[20-21]。 总之, 三者间的差异病毒的功能和感染的对象均不同, 表明WWTP 中的病毒群落分布具有空间异质性。 病毒通过侵染特定的宿主微生物进行繁殖, 宿主微生物的组成会直接影响病毒的种类, 因此, 本研究中WWTP 的病毒群落分布空间异质性主要是由于各处理单元宿主微生物种类不同而导致的。 探究不同处理单元中病毒的分布特征可以了解各单元的宿主微生物的组成, 从而可以评估各单元的生物处置效能。

图4 基于LEfSe 分析的原水、 厌氧池和活性污泥三者间的差异病毒Fig.4 Differential virus among raw water, anaerobic tanks, and activated sludge based on LEfSe analysis
2.4 病毒群落与细菌群落间的关系
基于Mantel 检验并利用Spearman 相关性分析, 检验了WWTP 中细菌群落多样性和丰富度与病毒群落的Bray-Curtis 相异度之间的关系。 Bray-Curtis 相异度可以衡量样本中不同物种组成的数量特征[22]。 在WWTP 中, 随着细菌群落的丰富度和多样性增加, 各样点中病毒群落越来越相似(图5), 尤其是细菌群落丰富度与病毒相异性之间的关系更显著(P=0.000 2, R=0.942)。 当细菌群落丰富度和多样性增加时, 可能会为病毒提供更多的生态位, 进而增加病毒数量, 并使得各样点间病毒组成越来越相似。 另一方面, 细菌群落丰富度和多样性的增加会使得菌落内部竞争加剧, 促进细菌产生更多抗病毒机制, 这将导致某些病毒消失, 从而使各样点间病毒群落越来越相似。 综上, 细菌群落丰富度和多样性的增加导致病毒群落的相似性增加, 这是宿主选择、 竞争和进化等因素综合作用的结果。 通过分析细菌群落α 多样性与病毒间的相关性, 阐明了污水处理过程中病毒群落增殖可能受到宿主(细菌)多样性调控, 这为污水处理过程的优化提供了一个新的视角, 如何减缓甚至阻断病毒利用WWTP 中宿主增殖是未来的重要研究课题。
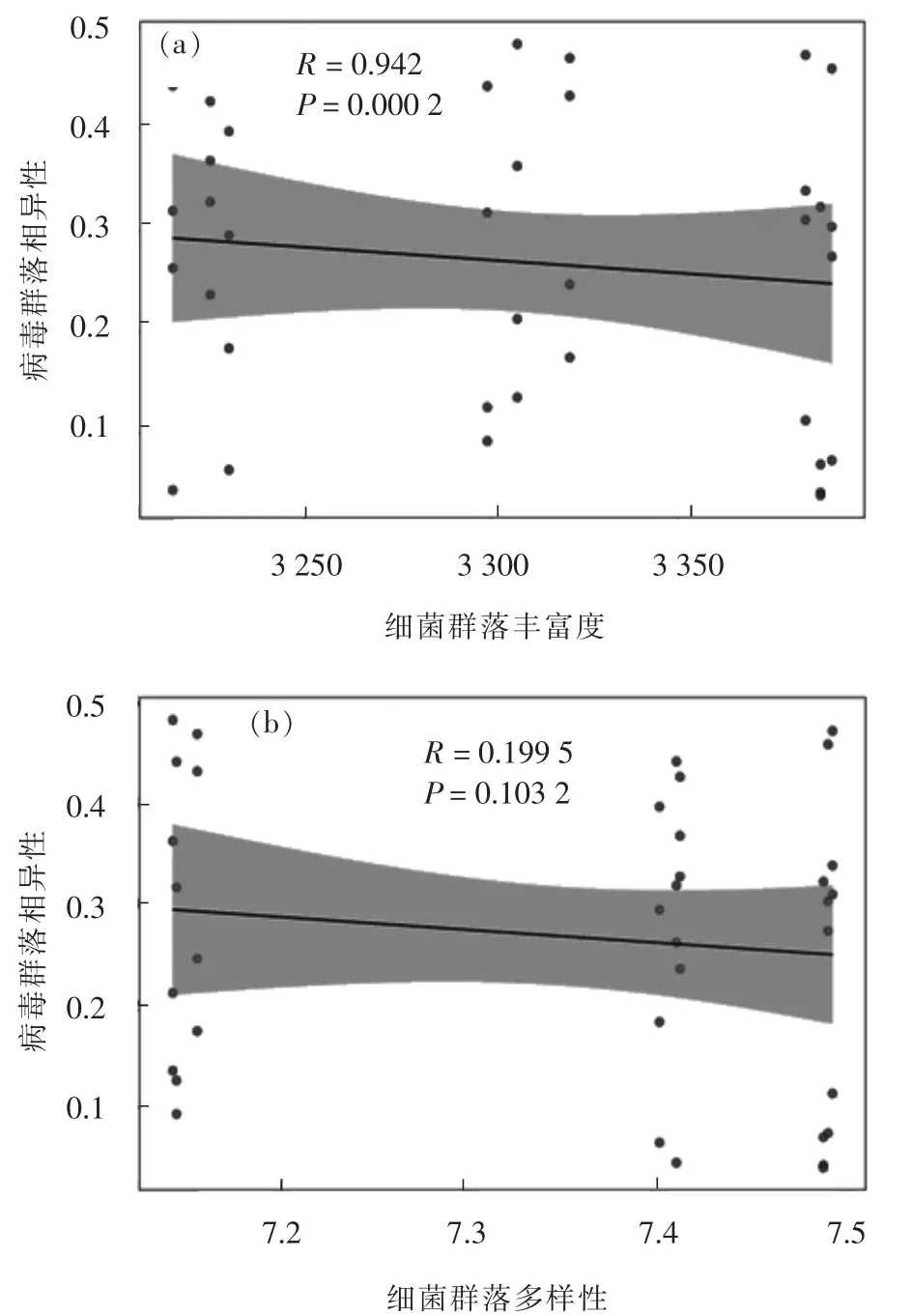
图5 细菌群落丰富度和多样性对病毒群落的相关性Fig.5 Correlation of bacterial community richness and diversity with viral communities
3 结论
通过宏基因组测序技术并结合LEfSe 差异分析, 对WWTP 核心单元中病毒群落的分布格局、 差异病毒群落和病毒-细菌间的相关性进行了深入讨论。 研究结果表明WWTP 中病毒群落分布具有空间异质性, 并广泛存在对人体或环境具有危害的病毒群落; 厌氧池的环境会限制病毒增殖和感染宿主,而活性污泥中数量众多的微生物会促进病毒群落的增殖; 病毒群落的相似性会随着细菌群落丰富度(P <0.05)的增加而增加, 表明病毒对宿主微生物具有直接调控作用以及对污水生物降解效能的间接调控。 本研究为WWTP 中病毒生态学提供了新的视角, 为污水处理效能的提升提供了科学依据。
