四川贡嘎山国家级自然保护区南北坡小型兽类物种多样性
2024-01-07王旭明李英廖锐刘莹洵郑笑傲乔江王宇王新刘少英
王旭明 ,李英 ,廖锐 ,刘莹洵 ,郑笑傲 ,乔江 ,王宇,王新*,刘少英*
1.四川省林业科学研究院森林和湿地生态恢复与保育四川省重点实验室,四川 成都 610081;
2.四川贡嘎山国家级自然保护区管理局,四川 康定 626000
贡嘎山是我国中低纬度海拔跨度最大的山脉,被称为“蜀山之王”。该区域是我国横断山脉南段生物多样性保护优先区和全球生物多样性保护热点地区。四川贡嘎山国家级自然保护区位于贡嘎山的腹心地带,东经101°29′—102°12′,北纬29°01′—30°05′,主要保护对象为山地生态系统及各类珍稀野生动植物资源。保护区内最高主峰7 556 m,最低处约1 000 m,相对高差逾6 000 m,拥有从亚热带到寒带的山地自然垂直带谱,生物区系复杂,生物资源丰富[1]。小型兽类通常被认为是环境指示物种,对判定生态系统健康与否起到非常重要的作用[2]。关于保护区的小型兽类调查资料始见于吴永杰等[3]对贡嘎山东坡非飞行小型兽类物种多样性的垂直分布格局研究,记录小型兽类3 目6 科25 种;谢淑强等[4]对保护区内兽类物种多样性与区系特征进行了研究,捕获小型兽类3 目4 科17 种。但吴永杰等主要调查保护区东坡,谢淑强等调查了南坡的东、西区,调查区域较小且样本量较少。
在喜马拉雅山脉、横断山脉等区域内,以雨量或温度分析物种密度分布趋势是不适宜的[5],纬度是小型兽类物种多样性的地理分布趋势的主要影响因素[6]。贡嘎山是横断山系大雪山支脉的主峰,呈南北走向,南北跨度长约60 km。因此,为进一步弄清保护区小型兽类多样性,2022 年3—10 月开展了四川贡嘎山国家级自然保护区小型兽类专项调查。调查研究的重点区域设置在四川贡嘎山国家级自然保护区的南坡及北坡,并对保护区南、北坡小型兽类生物多样性行了比较,以探究贡嘎山国家级自然保护区南、北坡小型兽类物种差异。
1 研究方法
1.1 样品采集
为完整覆盖保护区的各海拔段和生境类型,根据保护区地形地貌,在保护区内由北向南布设5 条样线,分别为保护区泸定入口—雅家埂;保护区康定入口烈士陵园—关门石沟—雅家埂;九龙猎塔湖沟口—峰顶;九龙瓦灰沟—峰顶;子梅村—垭口。海拔每上升300 m 设置一个调查区域,在调查区域内的不同植被类型分别布设样方,共计调查48 个样方。北坡调查6 个海拔段,分别为:2 400—2 700 m(植被类型均为阔叶林)、2 700—3 000 m(阔叶林、针阔混交林)、3 000—3 300 m(阔叶林、针叶林)、3 300—3 600 m(针阔混交林、针叶林、高山灌丛)、3 600—3 900 m(阔叶林、针阔混交林、针叶林、高山灌丛)、3 900—4 200 m(高山灌丛、流石滩植被);南坡因保护区范围内整体海拔较高,共调查4 个海拔段,分别为:3 300—3600m(植被类型包括阔叶林、针阔混交林、针叶林)、3 600—3 900 m(阔叶林、针阔混交林、针叶林、高山灌丛)、3 900—4 200 m(针阔混交林、针叶林、高山灌丛)、4 200—4 500 m(流石滩植被)。
采用铗夜法、桶捕法和笼捕法相结合的方式进行调查,总计下铗9 226 铗次,布设陷阱桶1 687 桶次,布设鼠笼60 笼次。
1.2 标本测量
捕获的样品经杀灭体表寄生虫后进行测量,外形测量包括体重(g),体长(mm),尾长(mm),后足长(mm)及耳高(mm)。测量后取肝脏、肌肉组织块,于99%分析纯乙醇保存,标本胴体用10%福尔马林溶液固定。带回实验室后,组织块于-80 ℃超低温冰箱保存,浸制标本制作成假剥制标本和头骨标本,用于标本鉴定。
1.3 物种鉴定
浸制标本制作成假剥制标本和头骨标本后,根据形态特征进行鉴定,主要参考依据为《中国兽类野外手册》[7],同时参考其他研究及专著[8-11]。
1.4 生物多样性计算
参照《基础生态学(第2 版)》[12]。不同群落内的物种组成按照鉴定结果统计,分别计算贡嘎山国家级自然保护区南、北坡的马格列夫指数(Margalef index),辛普森指数(Simpson index),香农威纳指数(Shannon-Wiener index)和佩洛均匀度指数(Pielou’s evenness index),公式如下:
马格列夫指数:
式中,R 为马格列夫指数,S 为物种数,N 为群落中全部物种总个体数;
辛普森指数:
式中,D 为辛普森指数,S 为物种数,Ni 为种i 的个体数,N 为群落中全部物种总个体数;
香农威纳指数:
式中,H 为香农威纳指数,S 为物种数,Ni 为种i 的个体数,N 为群落中全部物种总个体数;
佩洛均匀度指数:
式中,E 为佩洛均匀度指数,H 为香农威纳指数,S 为物种数。
2 结果
2.1 物种组成
本次调查共采集到小型兽类标本1 299 号,经鉴定包含3 目6 科16 属27 种(见表1)。其中劳亚食虫目2 科8 属12 种;啮齿目3 科7 属12 种;兔形目1 科1 属3 种。可见分布的27 种小型兽类中,劳亚食虫目和啮齿目最多,均为12 种,兔形目最少。

表1 贡嘎山国家级自然保护区小型兽类群落组成Tab.1 Community composition of small mammals in Gongga Mountain National Nature Reserve
从区系成分来看,分布的27 种小型兽类中,东洋界种类24 种,占88.89%,剩下的3 种均为古北界种类,占11.11%。从分布型来看,27 种小型兽类共有5 种分布型,分别是喜马拉雅横断山区型15 种、南中国型5 种、东洋型4 种、高地型2 种、东北-华北型一种。
采集数量占比超过10%的物种有6 个,分别为:小纹背鼩鼱、康定绒鼠、高原松田鼠、中华姬鼠、大耳姬鼠、川西白腹鼠。这6 个物种占比总和超过70%,表明这些物种为贡嘎山自然保护区小型兽类优势种。占比低于1%的物种有12 个,分别为:长吻鼹、少齿鼩鼹、川西缺齿鼩、斯氏缺齿鼩、甘肃鼩鼱、藏鼩鼱、珀氏长吻松鼠、大林姬鼠、大足鼠、黄胸鼠、滇攀鼠和中国鼠兔,表明这些物种在保护区内相对稀少。
2.2 南、北坡小型兽类物种组成比较
在贡嘎山自然保护区南坡采集到小型兽类标本22 种共586 个个体,捕获率为15.65%。其中劳亚食虫目2 科7 属9 种;啮齿目3 科7 属11 种;兔形目1 科1 属2 种。北坡采集到小型兽类标本19 种共713 个个体,捕获率为12.4%。其中劳亚食虫目2 科7 属10 种;啮齿目2 科4 属6 种;兔形目1 科1 属3 种(见表2)。南坡小型兽类物种数更多,捕获率更高。南、北坡均采集到的物种仅14 种,占物种总数的51.85%,分别为:长尾鼹、等齿鼩鼹、淡灰黑齿鼩鼱、斯氏缺齿鼩、小长尾鼩鼱、小纹背鼩鼱、藏鼩鼱、高原松田鼠、中华姬鼠、大耳姬鼠、北社鼠、川西白腹鼠、间颅鼠兔和藏鼠兔;仅在南坡采集到的物种有8 种,分别为:长吻鼹、川西缺齿鼩、珀氏长吻松鼠、康定绒鼠、大林姬鼠、大足鼠、黄胸鼠和滇攀鼠;仅在北坡采集到的物种有5 种,分别为:少齿鼩鼹、川鼩、甘肃鼩鼱、黑腹绒鼠和中国鼠兔。根据捕获率分析,南坡的优势物种是康定绒鼠、中华姬鼠、大耳姬鼠和川西白腹鼠,北坡的优势物种是小纹背鼩鼱、高原松田鼠、中华姬鼠、川西白腹鼠和藏鼠兔。

表2 南、北坡小型兽类捕获情况Tab.2 Capture of small mammals on the south and north slopes
不同物种垂直分布区大小存在差异,相同物种在南、北坡垂直分布区大小亦有差异(见图1)。在南、北坡均采集到的14 个物种中,除北社鼠和高原松田鼠外,其余物种在南坡的分布海拔上限均比北坡更高。北社鼠、藏鼠兔、川西白腹鼠、大耳姬鼠、淡灰黑齿鼩鼱、等齿鼩鼹、高原松田鼠、小纹背鼩鼱、小长尾鼩鼱、中华姬鼠等10 个物种在北坡的分布海拔下限低于南坡。长尾鼹、藏鼩鼱、间颅鼠兔、斯氏长尾鼩等4 个物种在南坡的垂直分布区完全覆盖了其在北坡的垂直分布区。仅小长尾鼩鼱在南、北坡的垂直分布区完全不重叠。除康定绒鼠和黑腹绒鼠外仅南坡或北坡采集到的物种垂直分布海拔范围均较窄(见图2),捕获的数量也较少(见表1)。
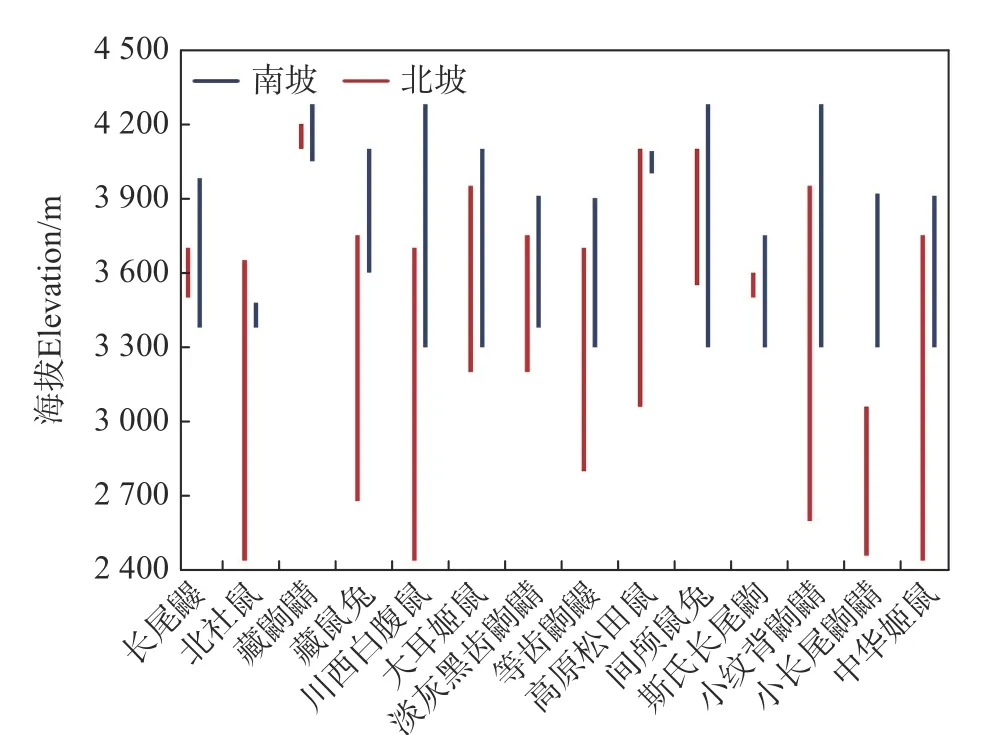
图1 南、北坡共同采集到的物种垂直分布海拔范围Fig.1 Elevational distribution of species captured both on the north and south slopes
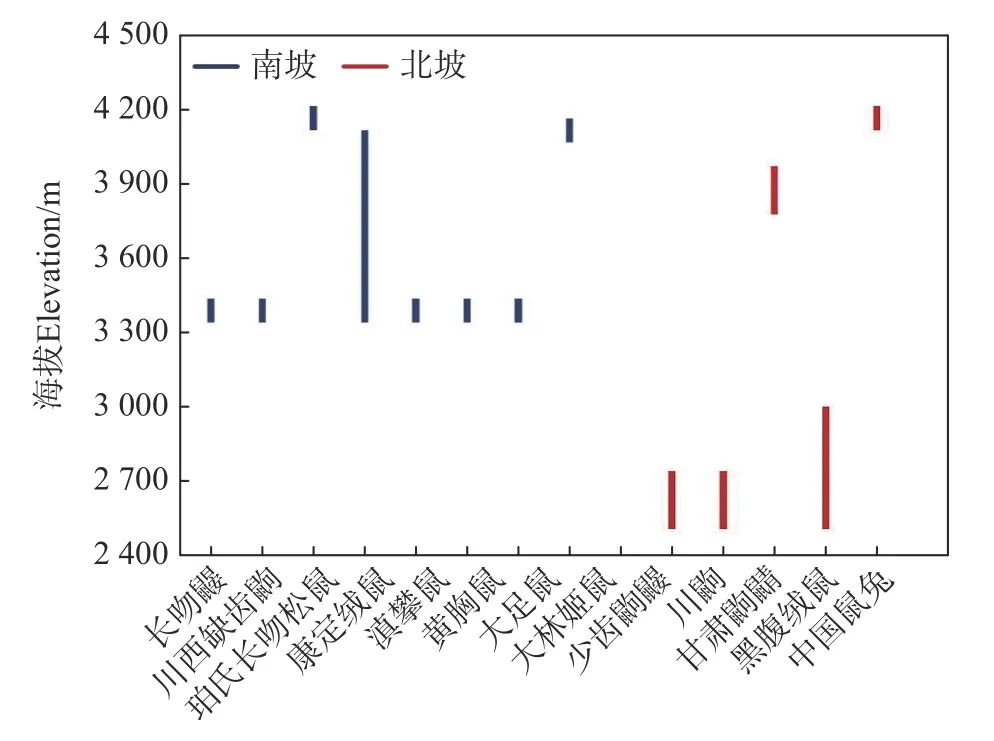
图2 仅南坡或北坡采集到的物种垂直分布海拔范围Fig.2 Elevational distribution of species captured only on the south or north slope
2.3 南、北坡小型兽类多样性指数
根据多样性指数计算结果(见表3),北坡的香农威纳指数和佩洛均匀度指数均高于南坡,表明北坡的小型兽类物种分布更均匀;南坡的马格列夫指数大于北坡,且辛普森指数小于北坡,表明南坡的小型兽类物种丰富度更高。
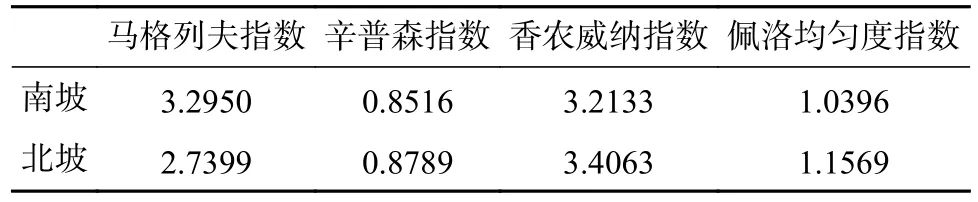
表3 南、北坡小型兽类多样性指数计算Tab.3 Calculation of small mammal diversity index on the south and north slopes
为了进一步探究南、北坡生物多样性差异的形成原因,对不同海拔段和不同生境分别进行生物多样性指数计算和比较。结果表明:南、北坡共同调查的3 个海拔段(3 300—3 600 m、3 600—3 900 m、3 900—4 200 m)中,南坡的捕获物种数、捕获个体数和马格列夫指数均大于北坡(见图3a、b、c);辛普森指数、香农威纳指数和佩洛均匀度指数呈现相似的趋势,3 300—3 600 m 与3 600—3 900 m 海拔段南坡低于北坡,3 900—4 200 m 海拔段南坡高于北坡(见图3,d、e、f);
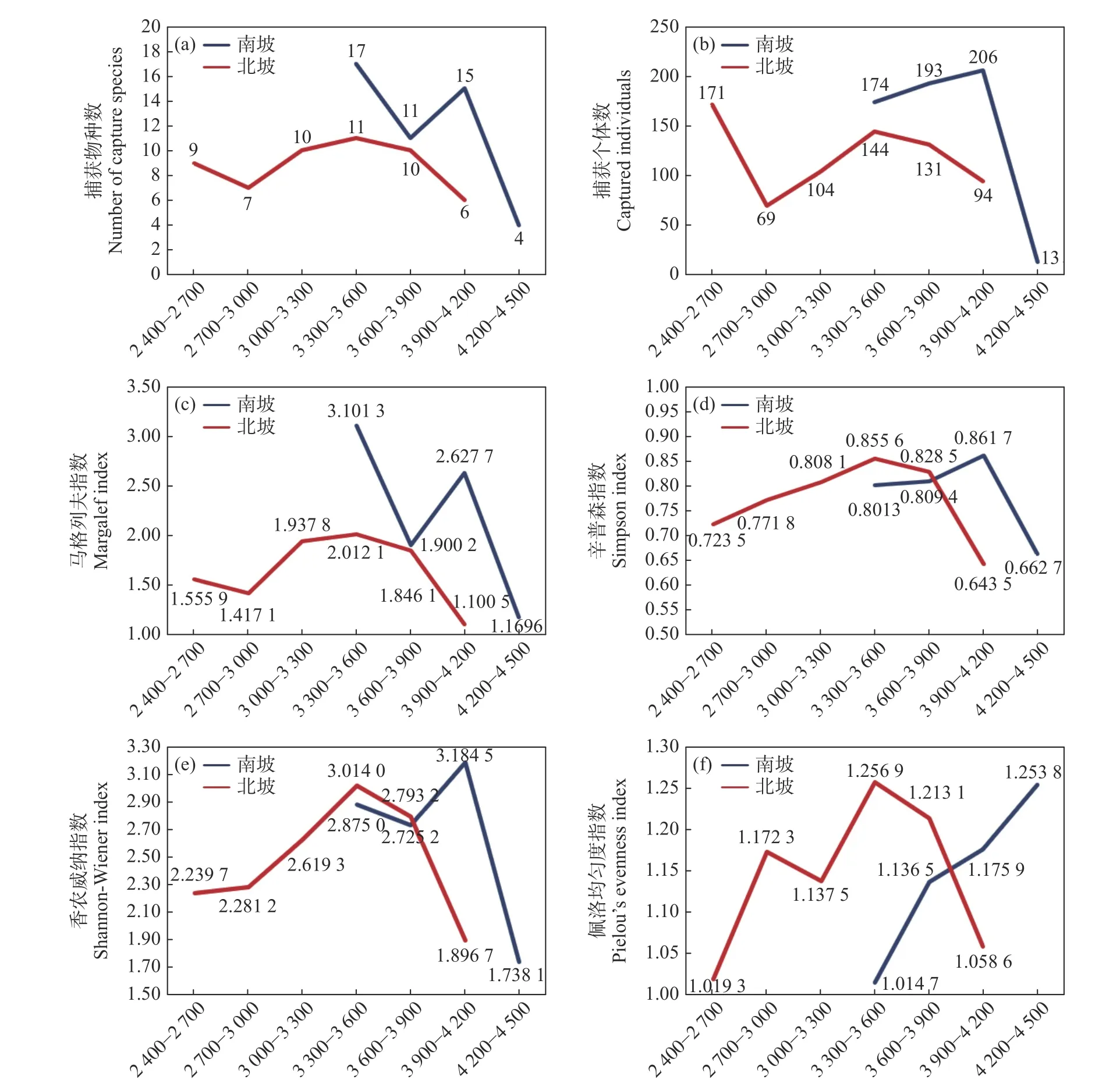
图3 南、北坡各海拔段生物多样性指数比较Fig.3 Comparison of biodiversity index at different elevations on the south and north slopes
保护区内南、北坡小型兽类调查的生境一致,均包含阔叶林、针阔混交林、针叶林、高山灌丛和流石滩植被5 种。南坡的捕获物种数和捕获个体数趋势均呈“M”型,北坡的捕获物种数和捕获个体数整体呈下降趋势(见图4,a、b);南、北坡马格列夫指数和香农威纳指数呈现一致的趋势,阔叶林生境南坡低于北坡,其余4 种生境南坡均高于北坡(见图4,c、e);南坡的辛普森指数均显著小于北坡(见图4,d),且南、北坡的佩洛均匀度指数趋势与捕获个体数趋势完全相反(见图4,f)。
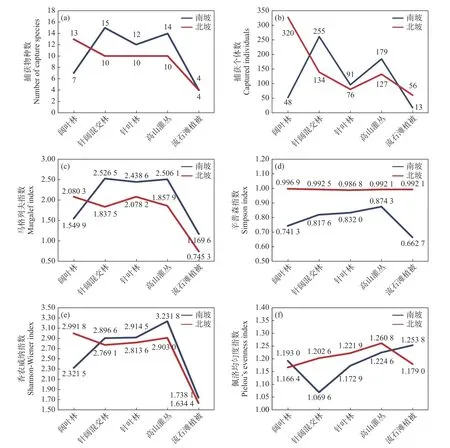
图4 南、北坡各生境生物多样性指数比较Fig.4 Comparison of diversity index of different habitats on the south and north slopes
3 讨论
3.1 小型兽类物种组成
共采集到小型兽类标本1 299 号,经鉴定包含3 目6 科16 属27 种,是历次保护区小型兽类调查采集标本数量最多的一次,也是物种数量最多的一次,对保护区小型兽类编目有重要意义。与此前调查研究比较[3-4],小型兽类增加了11 种:等齿鼩鼹、淡灰黑齿鼩鼱、斯氏缺齿鼩、川西缺齿鼩、藏鼩鼱、甘肃鼩鼱、小长尾鼩鼱、黑腹绒鼠、黄胸鼠、滇攀鼠和中国鼠兔。根据小型兽类的分类与分布研究[7-8],原来报道的甘肃鼹(Scapanulus oweni)、四川短尾鼩(Anourosorex squamipes)、蹼足鼩(Nectogaleelegans)、隐纹花鼠(Tamiops swinhoei)、华南针毛鼠(Niviventer huang)、高山姬鼠(Apodemus chevrieri)、小泡巨鼠(Leopoldamys edwardsi)7 种可能在保护区有分布,但本次调查未采集到。此外,此前报道的印度长尾鼩(Episoriculus leucops)、大绒鼠(Eothenomys miletus)、安氏白腹鼠(Niviventer andersoni)、黄毛鼠(Rattus losea)等可以确定是鉴定有误[13-15],贡嘎山并不是这些物种的分布区域。滇攀鼠是非常珍稀的物种,其分布范围较广但种群数量极小,本次发现对该种的分类研究有较大意义。中国鼠兔也是分布区域非常狭窄的珍稀种类,贡嘎山国家级自然保护区是其模式产地,本次采集到标本是该种命名100 多年来的首次再度发现,意义重大。
3.2 南、北坡小型兽类比较
由于青藏板块和扬子板块的挤压作用,使得大雪山系呈南北走向,并沿贡嘎山主脊线分成了在地质构造和地貌形态均有较大差异的东、西两坡[14],以往的小型兽类调查研究也主要集中在贡嘎山的东、西两坡。首次对贡嘎山南、北坡小型兽类物种多样性进行比较,结果与既往研究云南横断山区小型兽类物种多样性由南向北呈递减的趋势[7]是一致的。
南、北坡物种多样性差异主要体现在啮齿目和兔形目。南坡啮齿目物种比北坡多5 种,占本次调查所有啮齿目物种的42%,即近半的啮齿目物种仅在南坡采集到。其中大林姬鼠为古北界物种,在北坡没有采集到标本却在南坡有分布,其种群扩散路线与方式值得进一步研究。兔形目方面,南坡显然更适合其生存,物种数与个体数均大于北坡。仅在南坡采集到的康定绒鼠是狭域分布的物种,仅分布于九龙、康定和冕宁一带,Liu 等[13]通过研究将其由西南绒鼠康定亚种(Eothenomys custos hintoni)提升为种,贡嘎山南坡的独特地理特性可能是其物种形成的主要因素。仅在北坡采集到的少齿鼩鼹、川鼩、黑腹绒鼠3 物种是低海拔分布的物种,保护区南坡普遍高于其最高分布海拔,是不利于其种群扩散的因素。南、北坡共同分布的14 个物种中,有11 个物种在南坡分布的海拔上限更高,这与南坡的植被有关,南坡在海拔4 000 米以上还普遍存在针阔混交林和针叶林,这在北坡是极少的。南北坡共同调查的3 个海拔段中,南坡的捕获物种数与马格列夫指数均高于北坡,且南坡各生境辛普森指数均低于北坡,捕获物种数和马格列夫指数仅阔叶林生境低于北坡。表明南坡物种多样性高于北坡,根据实地调查推测,可能与南坡整体的湿度和温度更高,更适合啮齿目与兔形目物种繁殖有关。
3.3 贡嘎山国家级自然保护区的小型兽类研究现状
尽管贡嘎山国家级自然保护区的小型兽类研究已有十多年之久,但由于公众对小型兽类关注度很小,研究支持力度不够等原因,基础调查覆盖区域仅占保护区的很小范围,基础调查工作任重道远。总而言之,保护区的小型兽类还未完全厘清,贡嘎山周的小型兽类起源演化机制还未摸透,其研究价值依然巨大。
