模拟增温对五台山亚高山草甸植物群落与土壤性质的影响
2024-01-06郝爱华罗正明田志杰郑海霞郑庆荣
郝爱华,罗正明,田志杰,郑海霞,郑庆荣*
(1.忻州师范学院地理系,山西 忻州 034000; 2.忻州师范学院生物系,山西 忻州 034000)
IPCC第六次评估报告指出,人类活动导致大气中二氧化碳(CO2)等温室气体排放量增加,近130年来全球地表平均气温已上升约0.85℃,未来气温上升预计有加快趋势[1]。陆地生态系统每年能够吸收大气中大约31%的CO2,由此对气候变暖产生负反馈[2]。植物通过光合作用将大气中CO2以生物量的形式存储,据报道全球大约78%的潜在碳储量存在于植物生物量[3]。土壤是陆地生态系统最大的碳库[4]。植物生物量和土壤有机碳轻微的波动都能够引起大气中CO2含量的波动,进而影响气候变化[5]。全球草地总面积52.5亿hm2,占陆地总面积的 40.5%,其碳储量占全球陆地生态系统的34%,其中约 90%的碳储存在植物根系和土壤中[6]。植物多样性是生态系统结构和功能的基础[7],研究表明植物多样性可以增强草地生态系统的碳汇功能[8]。气候变暖导致植物多样性减少或丧失,必然造成草地生态系统功能下降[9]。因此,草地的生物量、土壤有机碳和植物多样性对气候变暖如何响应备受生态学家关注。
亚高山草甸作为一种特殊的草地生态系统类型,不仅在调节气候、保持水土、维持生物多样性等方面发挥重要的生态功能,而且可以为牲畜提供食物来源,极具经济价值。亚高山草甸是高寒草甸的一种类型,以耐寒冷、密丛短根茎地下芽嵩草以及苔草、禾草、杂类草为建群植物的草甸群落[10]。亚高山草甸地处高寒地带,生态系统脆弱,对气候变化响应极为敏感,是监测气候变化的理想试验场所和研究生物多样性保护的热点地区[11-12]。目前基于增温控制试验监测亚高山草甸植被特征和土壤特征对气候变暖响应的研究已取得一系列的成果。研究发现增温处理下亚高山草甸的地上生物量增加而根系生物量下降[13-15]、植物群落结构发生改变[11,13,16]、植物多样性减少[17]、优势物种基部茎粗与高度由等速增长变为异速增长[18]。增温使亚高山草甸土壤呼吸增强[15,19]、表层土壤有机质显著下降[20]。这些成果为我们深入理解亚高山草甸生态系统响应全球变化提供了坚实的基础。
山西省地处中纬度地区,亚高山草甸分布广泛,在五台山、云顶山、太岳山、恒山和中条山等山系林线以上的高海拔地带多有发育,面积达35.3万hm2,约占山西省草地总面积的10%。五台山素有“华北屋脊”和“华北水塔”之美称,是京津唐及雄安新区乃至整个华北地区的绿色生态安全屏障。五台山亚高山草甸是华北地区海拔最高、面积最大、最典型的高山夏季牧场,面积约10.7万hm2[21]。近年来,由于气候变化和过度放牧、旅游、新能源开发、采矿等人类活动的共同作用,五台山亚高山草甸退化严重,导致生态系统服务功能和生产功能降低,严重威胁着区域的生态安全和人们的生产生活[22-24]。目前关于五台山亚高山草甸植物群落和土壤理化性质响应气候变暖的监测很少,仅有的研究也只是局限于增温对土壤呼吸的影响[19]。这将不利于五台山地区退化草地的治理恢复和地方社会经济的可持续发展。
为此,本研究以五台山亚高山草甸为研究对象,采用开顶式(Open top chambers,OTCs)增温控制试验,探讨模拟增温对植物多样性、群落结构、地上生物量(Aboveground biomass,AGB)和地下生物量(Belowground biomass,BGB)的影响;分析模拟增温对土壤温度(Soil temperature,ST)、土壤水分(Soil moisture,SM)、土壤有机碳(Soil organic carbon,SOC)、总氮(Total nitrogen,TN)、pH值、有效磷(Available phosphorus,AP)和速效钾(Available potassium,AK)的影响;阐明植物群落与土壤理化性质的关系。本研究不仅有助于理解高山、亚高山草甸对气候变化的响应机制,而且可为区域生态系统长效增汇提供重要的理论依据。
1 材料与方法
1.1 研究区概况
研究区五台山位于山西省忻州市东北,地理坐标38°27′~39°15′ N,112°48′~113°55′ E,海拔610~3 061 m,具有相对完整的高山和亚高山草甸生态系统(图1)。试验样地选择五台山中台亚高山草甸地区,海拔2 500~2 800 m之间。该区地貌以石质山为主,土壤生成结构不完整,属于亚高山草甸土,土壤较薄,厚度一般在20~30 cm。该区有典型的冰缘地貌,有些地方常年积冰,属于温带大陆性季风气候,四季分明。冬季寒冷,春季干旱多风,夏季雨量集中,秋季短暂晴朗。年均温差6℃~8℃,年均降水量500~650 mm,无霜期130天。五台山亚高山草甸优势物种以莎草科的矮生嵩草(Kobresiahumilis)为主,伴生种有珠芽蓼(Polygonumviviparum)、直梗高山唐松草(Thalictrumalpinumvar.elatum)、铃铃香青(AnaphalishancockiiMaxim)、蓝花棘豆(Oxytropiscoerulea)等杂类草。

图1 五台山研究区域及增温试验样点定位图Fig.1 Location map of Wutai Mountain study area and warming experimental sites
1.2 试验设计
相比于红外辐射增温,开顶式小室增温模拟增温效果好,稳固性较强,能够降低空气流动对装置的影响,开放式增温更接近自然,而且成本低廉,节能环保,因此更适合坡地地形、地处偏远和通电困难的研究区域。根据研究需要,课题组于2019年8月在五台山亚高山草甸设置开顶式小室增温控制试验,于海拔2 544,2 631和2 764 m设置了3个观测样点(图1)。每个样点设置开顶式小室5~6个,相邻小室间隔3~5 m,随机选择4个小室进行植被和土壤参数调查。对照样地选择邻近增温调查样地,群落特征类似对照样地,每个对照样地设置4个重复观测样方。增温和对照样地共设置24个调查样方,每个样方大小为0.5 m×0.5 m。开顶式小室设计为六边形,上边长40 cm,下边长75 cm,垂直高度60 cm。样地周边设置围栏,以防止动物破坏。
1.3 样品采集与测试
1.3.1植物群落调查 2022年8月中旬课题组于植被生长季高峰期进行样地调查,获取植被数据。调查内容包括每个样方内植被群落高度(Community height,CH)、群落盖度(Community coverage,CC)、物种多度、频度、物种名称,同时记录每个样地的经纬度、海拔及周边地形地貌信息。物种高度用钢卷尺测量,盖度和频度用网格法获得。
地上生物量采用刈割法收获。将每个样方内植物地上部分全部刈割,分成莎草、禾草和杂类草,分别置入信封内,放入75℃烘箱烘干至恒重并称重。地下生物量采用土钻法获取。因为高寒草甸的根系集中分布于土壤表层,因此本研究只钻取了0~10 cm土层根系。根系样品带回实验室,冲洗干净。根据根的颜色、断面、柔韧性及是否附着须根,分出活根和死根。再将活根放入信封内,置入75℃烘箱烘干至恒重并称重,记入地下生物量。每个样方钻取的根系生物量均被换算成单位面积的地下生物量。每个样地的地上和地下生物量取各个小样方生物量的平均值。
1.3.2土壤样品收集 采用土钻进行土壤样品采集。每个样方多点采集,混合为一份土壤样品,共收集24个混合土样。所采土壤样品尽早带回实验室用以分析土壤水分、有机碳和总氮、pH值等理化性质。分析前要去除土壤中的植物、石块等并过2 mm钢筛。
1.3.3土壤理化性质测定 所有收集的土壤样品及时送至中国科学院西北生态环境资源研究院沙漠与沙漠化重点实验室进行测定。pH值采用pH计法获得(HZP-T502,Huzhi,福建),土壤水分测定采用烘干法。有机碳测定采用浓硫酸-重铬酸钾氧化滴定法[25],全氮测定采用凯式定氮法[26],硝态氮和铵态氮测定采用紫外可见光分光光度计(UV-5100,元析,上海)。
到了宋代,民间养狗已为常见,城市中出现了专门的宠物市场,宋人孟元老《东京梦华录》说,开封府的大相国寺,“每月五次开放万姓交易,大三门上皆是飞禽猫犬之类,珍禽奇兽,无所不有”。市场上还有猫粮、狗粮出售:“凡宅舍养马,则每日有人供草料;养犬,则供饧糠;养猫,则供鱼鳅;养鱼,则供虮虾儿。”
1.4 数据分析
1.4.1物种重要值 物种重要值(Importance value,IV)能够反映物种在群落中的优势程度,是由相对高度(Relative height,RH)、相对盖度(Relative coverage,RC)和相对频度(Relative frequency,RF)三者之和的平均值计算得出,公式如下:
(1)
注:相对盖度=样方内各物种的分盖度/物种总盖度;
相对高度=各物种的平均高度/样方内所有物种的平均高度之和;
相对频度=样方内各物种的频数/所有物种的频数之和。
1.4.2植物多样性 采用α多样性指数对五台山亚高山草甸增温处理下的植物多样性进行分析,其指标包括:物种丰富度(Species richness,SR)、香农-威纳(Shannon-Wiener)多样性、辛普森(Simpson)优势度和Pielou均匀度指数。计算公式如下:
(2)
(3)
(4)
(5)
式中,IV为物种重要值,H′为Simpson指数,H为Shannon指数,E为Pielou指数;S为样方内物种数总和,i为样方内第i个物种,Pi为第i个物种的相对重要值,T为样方内所有物种重要值的总和。
1.4.3统计分析 采用ArcGIS 10.5和Origin 2017软件绘制地图;统计分析在IBM SPSS Statistics (SPSS 23.0)软件中完成;通过冗余分析(Redundancy analysis,RDA) 和Pearson相关性检测植被指标与土壤理化性质的关系,冗余分析在Rstudio软件中进行,Pearson相关性通过SPSS软件完成。
2 结果与分析
2.1 增温对植物多样性的影响
由图2可知,增温以后物种丰富度、香农多样性和辛普森优势度指数均显著增加(P<0.05),其中物种丰富度和香农多样性指数显著增加了20.23%和34.35%。Pielou均匀度指数增温处理下无明显变化。结果表明:增温处理下五台山亚高山草甸植被α多样性呈现不同的变化特征。

图2 增温处理下植物多样性指数变化Fig.2 Changes of plant diversity index under warming treatment注:不同小写字母表示不同处理间差异显著(P<0.05),下同Note:Different lowercase letters indicate significant difference between different treatments at the 0.05 level,the same as below
2.2 增温对群落结构和物种组成的影响
增温以后群落平均高度和平均盖度均显著增加(P<0.05)。增温样地相比对照样地群落平均高度增加了3.29倍,平均盖度则增加了28.86%(图3)。由表1可知,对照样地亚高山草甸建群种矮生嵩草在群落中占据绝对优势,其重要值为0.31,明显高于群落中其它物种的重要值,伴生种珠芽蓼、翻白草(Potentilladiscolor)、蓝花棘豆、直梗高山唐松草的重要值分别为0.13,0.07,0.07和0.06。增温样地矮生嵩草的重要值为0.08,明显小于优势种珠芽蓼(0.15)、翻白草(0.11)和紫花地丁(Violaphilippica,0.09)。对照样地的物种毛茛(Ranunculusjaponicus)和假水生龙胆(Gentianapseudoaquatica)在增温样地中消失,取而代之的是尖嘴苔草(Carexleiorhyncha)和高山早熟禾(Poaalpina)。草地退化的标志性物种火绒草(Leontopodiumleontopodioides)也在增温样地出现。由以上结果可知:矮生嵩草在五台山亚高山草甸对照样地群落中占据绝对优势,而增温处理下其优势地位被杂类草取代,增温以后五台山亚高山草甸群落结构和物种组成发生改变。

表1 增温处理下物种重要值变化Table 1 Changes of species importance value under warming treatment

图3 增温处理下群落高度和盖度变化Fig.3 Changes of the community height and coverage under warming treatment
2.3 增温对地上和地下生物量的影响
五台山亚高山草甸地上和地下生物量对增温响应敏感。由图4可知,亚高山草甸增温样地地上生物量为154.91 g·m-2,相比对照样地显著增加了约2.83倍(P<0.05)。对照样地的地下生物量为2 465.01 g·m-2,增温以后显著下降了约35.96%(P<0.05)。3年增温以后,功能群禾草地上生物量由0.81 g·m-2显著增加至18.94 g·m-2(P<0.05),嵩草和杂类草地上生物量显著增加了1.0倍和2.18倍(P<0.05)。对照样地功能群嵩草的地上生物量值最高(30.12 g·m-2),增温样地功能群杂类草地上生物量值最大(75.74 g·m-2)。
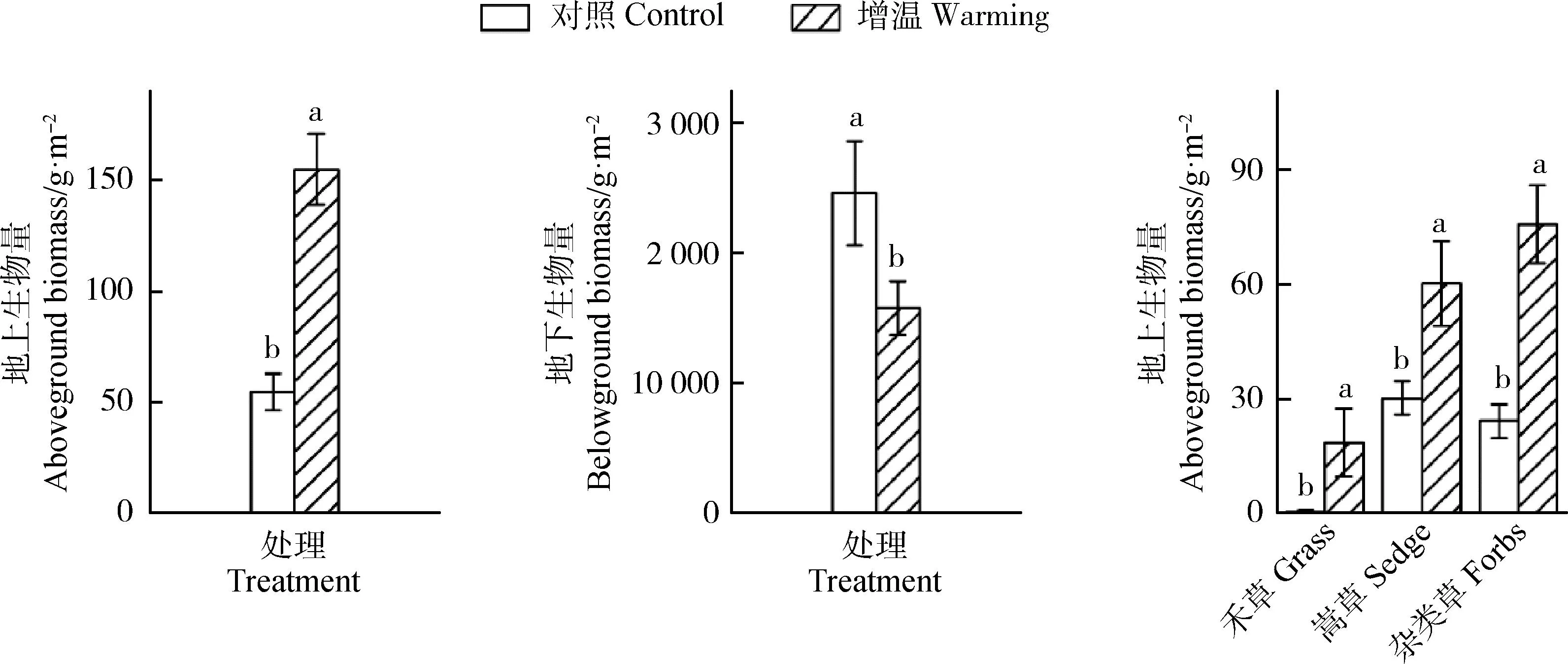
图4 增温处理下地上、地下生物量及禾草、嵩草和杂类草地上生物量的变化Fig.4 Changes of the above- and belowground biomass and the aboveground biomass of grass,sedge,and forbs under warming treatment
2.4 增温对土壤理化性质的影响
由表2可知,增温处理下五台山亚高山草甸表层土壤温度显著上升了1.25℃(P<0.05),土壤水分含量显著下降了24.58%(P<0.05)。增温以后表层土壤有机碳由27.92 g·kg-1下降至24.61 g·kg-1,显著下降了11.86%(P<0.05),总氮由6.06 g·kg-1下降至4.86 g·kg-1,显著下降了19.80%(P<0.05)。对照样地的速效钾为167.72 mg·kg-1,增温处理下略微增加了21.73 mg·kg-1。pH和有效磷增温处理下则无明显变化。因此,增温处理下五台山亚高山草甸表层土壤温度上升,土壤水分减少,表层土壤趋于暖干化,表层土壤有机碳和总氮发生流失,其中总氮流失量大于有机碳流失量。

表2 增温处理下土壤理化性质变化Table 2 Changes of the soil physicochemical properties under warming treatment
2.5 植物群落与土壤理化性质的关系
图5展示了五台山亚高山草甸对照样地和增温样地群落高度、盖度、物种丰富度、香农多样性、辛普森优势度、Pielou均匀度、地上生物量、地下生物量与土壤温度、水分、有机碳、总氮、pH值、有效磷、速效钾的RDA排序结果。对照和增温样地RDA排序轴第一主成分分别解释了82.69%和91.49%。
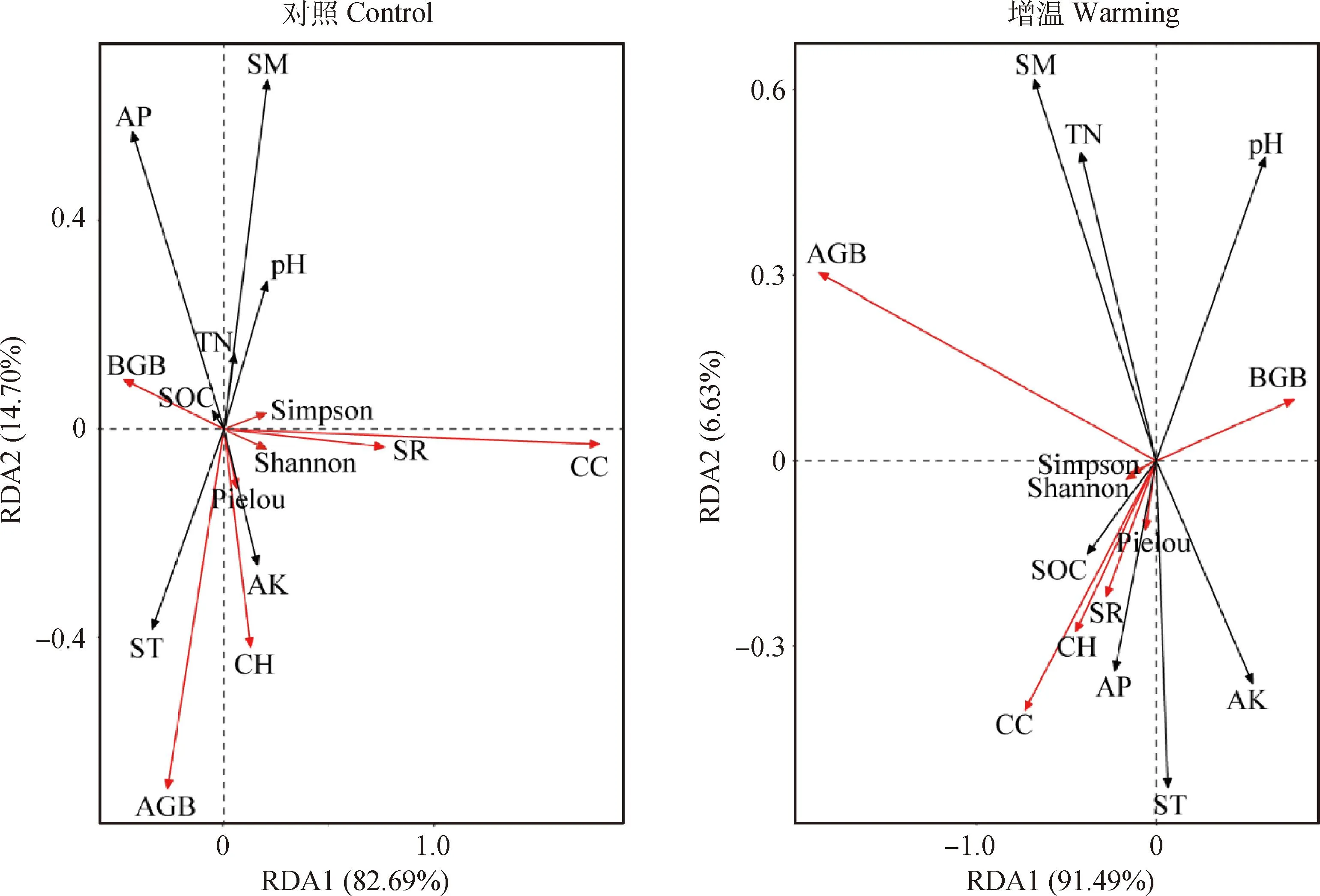
图5 对照样地和增温样地植被要素和土壤因子RDA排序Fig.5 RDA ordination biplot of plant variables and soil factors at the controlled and warmed plots注:图中红色箭头表示植被要素,黑色箭头表示土壤因子。AGB,地上生物量;BGB,地下生物量;CC,群落盖度;CH,群落高度;SR,物种丰富度;Shannon,香农多样性;Simpson,辛普森优势度;Pielou,Pielou均匀度;ST,土壤温度;SM,土壤水分;SOC,土壤有机碳;TN,总氮;pH,pH值;AP,有效磷;AK,速效钾Note:The red and black arrows denote plant variables and soil factors in the figure,respectively. AGB,Aboveground biomass;BGB,Belowground biomass;CC,Community coverage;CH,Community height;SR,Species richness;Shannon,Shannon diversity;Simpson,Simpson dominance;Pielou,Pielou uniformity;ST,Soil temperature;SM,Soil moisture;SOC,Soil organic carbon;TN,Total nitrogen;pH,pH value;AP,Available phosphorus;AK,Available potassium
就对照样地而言,土壤温度和速效钾与平均高度呈正相关,而土壤水分与平均高度呈现显著负相关(P<0.01);土壤总氮与平均盖度显著正相关(P<0.05),土壤温度与平均盖度呈负相关;土壤温度与物种丰富度呈显著负相关(P<0.01),皮尔森相关性系数为-0.79,土壤速效钾与物种丰富度呈显著正相关(P<0.01),皮尔森相关性系数高达0.80;土壤温度与香农多样性呈显著负相关(P<0.05),皮尔森相关性系数为-0.70,总氮与香农多样性和辛普森优势度呈显著正相关(P<0.05),土壤水分与辛普森优势度呈正相关,但不显著;土壤温度和速效钾与地上生物量呈显著正相关(P<0.05);土壤有机碳、总氮、有效磷和土壤水分与地下生物量正相关,其中有效磷与地下生物量的相关性达显著性水平(P<0.05)。
就增温样地而言,土壤有机碳和有效磷与平均高度呈显著正相关(P<0.05),土壤温度和速效钾与平均高度呈正相关但不显著;土壤温度、有机碳、有效磷及速效钾与平均盖度、物种丰富度、香农多样性和辛普森优势度均呈正相关,但不显著,而土壤水分和总氮与平均盖度、物种丰富度、香农多样性和辛普森优势度均呈负相关,但不显著;土壤水分和总氮与地上生物量呈显著正相关(P<0.01),皮尔森相关性系数分别达0.87和0.63;pH值与地下生物量呈显著正相关(P<0.05)。
3 讨论
本研究发现五台山亚高山草甸植物多样性对模拟增温响应敏感,增温以后群落物种丰富度和香农多样性显著增加(图2)。在海拔2 544 m试验样地,增温以后物种数由14增加至17。云顶山亚高山草甸增温试验则发现,植物多样性呈现下降趋势,与本研究结果不一致[17],主要是由于增温时长差异造成的。晁倩等的增温时间仅有一年,而本研究的增温时间为3年。与本研究相近的增温时长条件下,青藏高原高寒草甸植物多样性在红外辐射增温条件下略微增加[27],而在OTCs增温处理下则显著下降[28]。出现这些差异的结果,主要是由于生态系统不同组分和过程对温度的敏感性不同,导致植物多样性在温度上升时呈现不同的变化,而且不同增温装置的加热机制不同,可能会引起陆地植物和生态系统特异性的反应[29]。另外,增温控制试验属于点上的数据,受微地形、微地貌、微气候条件限制,其试验结果往往具有局限性。
增温处理下五台山亚高山草甸的群落结构发生明显改变,建群种矮生嵩草的优势地位被杂类草取代(表1)。类似的研究结果前人也曾报道[13,30-31]。这一现象主要受到植物本身生理生态特征的调控[32],其中的机理主要表现在嵩草类植物和杂类草植物根系的分布深度和构建特征不同[33]。嵩草类植物拥有庞大的根系,但分布较浅,绝大部分在表层土壤0~10 cm。嵩草草甸多在冷湿的生境条件下发育,对土壤水分依赖性很强。增温显著降低了高寒草甸表层土壤水分,从而引起表层土壤干旱,对浅根系的嵩草类植物生长不利[34]。另外,嵩草类植物根系构建特征表现为根系多为横向根系,而且毛根发达,彼此纠结缠绕,形成结实而富有弹性的草毡层,极耐畜牧践踏。在表层土壤干旱胁迫条件下,缠绕的特征不利于根系向土壤深层延伸,因而很难在短期内吸收土壤深层水分以缓解表层土壤干旱胁迫,因而面临死亡。直根系的杂类草则可以向土壤深层延伸,从而能够吸收土壤深层水分以缓解表层土壤干旱[33]。此外,随着表层土壤的干旱化,群落物种组成也发生了明显变化,对照样地中一些喜湿的物种如毛茛、假水生龙胆在增温样地中消失,取而代之的是一些耐旱的禾草和苔草(表1)。
温度上升使五台山亚高山草甸地上生物量显著增加,但0~10 cm根系生物量却显著下降(图4)。前人研究也证实了这一结果的正确性[34-35]。一直以来温度都是高寒草甸植物生长的限制性因子,增温处理下,空气温度和土壤温度均上升,从而缓解了低温对高寒草甸植物生长胁迫,刺激了植物地上部分的生长。随着全球气候变暖,温度不再是高寒生态系统植被生长的限制性因子,水分则发挥了更大的调控作用[36]。研究表明近年来青藏高原高寒草甸植被生长过程中,温度上升和降水增加的地区植被指数呈现增加态势,而温度上升降水减少的地区植被呈现退化态势[37-38]。这一机制也解释了本研究中亚高山草甸地上生物量在对照处理下与土壤温度呈正相关,而在增温处理下则与土壤水分呈正相关(图5)。增温以后根系生物量减少主要是由于增温导致的表层土壤水分减少,引起表层土壤干旱造成的[34]。植物的资源分配权衡策略(Trade-offs)很好地解释了增温以后五台山亚高山草甸地上生物量增加而根系生物量减少的现象,即植物生长受到土壤条件胁迫时,为了实现生存效率最大化,植物会将更多的生物量分配给地上的叶子和茎而非地下的根系,以获取更多的光和CO2资源[39]。前人的研究表明高寒草甸增温以后由于直根系的杂类草增加,导致土壤深层根系增加[23]。本研究只关注0~10 cm土壤层根系生物量的变化。因此,五台山亚高山草甸深层根系生物量对气候变暖如何响应有待进一步研究。
增温以后五台山亚高山草甸表层土壤温度上升、水分含量减少、有机碳和总氮明显下降(表2),前人对类似生态系统的研究也得出过以上结论[34]。温度上升使植物蒸腾和土壤蒸散发作用加强,导致土壤中可利用的水分含量减少。土壤有机碳的减少主要有两方面原因造成的,一是有机碳的来源减少,二是有机碳的分解速率加快。土壤有机碳主要来源于地上植物凋落物和根系的输入,本研究发现增温以后群落地上生物量显著增加,必然引起地上凋落物的增多,这与土壤有机碳减少逻辑不符,而且凋落物的分解需要一个漫长的过程。增温以后表层根系生物量减少是导致土壤表层有机碳损失的原因之一。另外,土壤有机碳的损失有可能是由于温度上升加快了土壤呼吸作用[19,40],同时增温促进了土壤微生物的活动,从而导致土壤有机碳的分解速率加快,土壤有机碳减少[41]。本研究发现增温以后总氮的下降速率高于有机碳,可知总氮对增温的敏感程度大于有机碳,而且物种多样性在对照样地与总氮呈正相关而在增温样地则呈负相关,增温处理下地上生物量与总氮显著正相关(图5)。这些结论表明总氮在高寒草甸生态系统植被响应气候变暖过程中的调控作用不容忽视。
4 结论
3年增温处理下五台山亚高山草甸植被α多样性呈现不同的变化特征。增温使亚高山草甸群落结构和物种组成发生改变,矮生嵩草的优势地位被杂类草取代,草地退化的标志性物种火绒草出现,亚高山草甸发生了一定程度的退化。增温使亚高山草甸表层土壤趋向干旱化,表层根系减少。增温条件下表层土壤养分流失,呈现贫瘠化态势。总氮对增温的敏感程度大于有机碳,总氮在高寒草甸生态系统植物群落响应气候变暖过程中的调控作用不容忽视。
