湖泊微生物群落构建机制及研究展望
2024-01-05张含笑张靖天翁南燕张佩莲马春子霍守亮
李 怡,张含笑,张靖天,翁南燕,张佩莲,马春子,霍守亮
环境基准与风险评估国家重点实验室,中国环境科学研究院,北京 100020
湖泊是地球表层系统重要组成部分,是水资源的重要储存和调节单元,具有削洪调蓄作用,能够提供饮用水源和生物栖息地等[1-2]。在湖泊生态系统中,微生物是能量流动和生物地球化学循环的参与者,在介导碳、氮、磷等元素循环的过程中发挥重要作用,对维持生态系统的相对稳定和代谢过程具有关键意义[3-4]。湖泊微生物研究是通过解析微生物时空分布规律、群落结构对环境变化的响应及反馈机制、微生物与其他生物之间的复杂相互作用,来认识湖泊生态系统的结构与功能,评估湖泊生态系统状态,探究生态系统稳定性及调节机制的[5-6]。
生物化学法和组学方法的应用为研究人员提供了大量未培养微生物的信息,提高了微生物识别的精度与广度,使微生物生态学研究的核心问题从发现和描述时空分布规律,延伸到探究驱动微生物时空分布的生态过程[7-9]。系列理论与假说曾被用于解释微生物群落时空演变的生态过程,发展至今,主流观点支持将生态位理论与中性理论整合,认为确定性和随机性过程共同作用于微生物群落构建,这些过程涉及环境选择、生物相互作用、扩散限制、物种分化与生态漂变等[8]。群落构建机制研究是对生态过程进行区分和量化,通过统计分析法、模型推断法、网络分析法解释湖泊微生物对干扰的响应,有助于理解湖泊微生物多样性维持机制。笔者综述了湖泊微生物时空分布、群落构建的生态过程,群落构建机制分析方法及研究展望,以期为湖泊微生物群落构建机制研究提供参考。
1 湖泊微生物的时空分布
1.1 湖泊微生物研究的时空尺度
微生物生态模式受研究尺度的严格限定,微生物时空分布规律具有显著的时空尺度依赖性,因此明确研究对象的时间、空间尺度是理解微生物群落构建机制的前提[10]。
时间尺度研究范围可以为天、月、季节、年乃至更久。短时间尺度的研究有利于反映微生物群落敏感的、突如其来的变化,较长时间尺度上微生物的时空分布规律具有季节性或者年份间差异。在长达百年的时间尺度上,古湖沼学为揭示湖泊微生物群落的时间演替规律提供了可行方法,不同历史时期的湖泊信息储存于相应深度的沉积物中[11-12]。因此,对不同沉积物演替序列中微生物群落的分析,可以解析微生物群落组成结构变化及其驱动机制[13-14]。空间尺度的研究范围可以限定在微小尺度、局域尺度,或通过大量数据整合与再分析扩展到区域尺度、洲际尺度和全球尺度等[15-16]。从空间角度研究微生物群落,能够反映多尺度下微生物地理分布格局的差异。
1.2 湖泊微生物研究的时空分布格局
关于湖泊微生物群落在天、月、季节和年际时间尺度动态演变的已有研究包括:①群落总体结构的变化(如微生物群落结构的年间变化[17]、月变化[18]、季节变化[19]);②群落组成变化(如冬季、夏季门类丰度的变化[19]);③物种周转(如物种总数随时间推移而增加[20])。相关研究通过调查微生物在不同时期的群落动态,探讨季节性调水工程对浮游植物群落的影响和贡献[21]、浮游细菌群落对湖泊中氮磷输入的响应等问题[22]。对湖泊沉积物中微生物群落演变信息的挖掘等相关研究,揭示了百年尺度的微生物多样性模式和生态系统变化[13,23-24]。
在空间分布格局上,由于微生物对环境条件变化的敏感性高、适应性强,湖泊微生物群落分布随自然地理条件和水体环境变化呈现相应规律(如浊度等环境因子使抚仙湖固氮微生物群落在丰度、多样性和群落组成上存在明显的空间异质性[25])。同时,由于微生物主动扩散能力差,地理格局产生的扩散限制也可能导致物种组成相似性呈现随地理距离衰减的现象(如扩散限制影响中国东南地区24座水库细菌群落的构建过程[26-27])。
1.3 时空分布特征的主要影响因素
湖泊环境条件是影响微生物时空分布的主要因素,天然水体中光强随深度增加而衰减、湖泊热力及溶解氧等分层现象使湖泊内部环境具有异质性,特殊湖泊可能具有更显著的化学分层,影响着不同环境适应性的水体微生物的垂直分布[28-30]。pH、盐度等环境因素已被证明对湖泊微生物群落结构和组成均具有重要影响,甚至在相距很远但pH相近的湖泊中存在着相似的微生物组基因[31-33]。水位波动、湖泊内部或与外界水交换等水动力条件变化与多种类群联系密切(如浮游藻类群落在湖流速度增加时,优势类群逐渐向绿藻门和硅藻门转变[34-35])。
湖泊微生物分布还受到生长资源及种间关系的制约。自养型微生物分布受光照条件和营养物质(磷酸盐、硝酸盐、铁等)可获得性的影响,同时和异养型微生物一样,在湖泊生态系统中受到正相互作用和负相互作用的影响,在获取资源时存在着合作与竞争关系[36-37]。
地理因素对湖泊微生物分布的影响体现在随着湖泊之间的地理距离增加,微生物群落组成的相似度逐渐降低[31]。但由于微生物扩散能力强、种群密度高,其群落组成的海拔分异较弱,水生微生物的物种丰富度和α多样性随海拔的升高会呈现出不同的规律(如降低、增加、驼峰形、U形变化或无明显变化趋势[38])。
2 湖泊微生物群落构建的生态过程
2.1 微生物生态学基础理论
群落构建是指群落生物多样性形成、维持和变化的生态学过程[38-39]。群落构建机制研究中的生态过程划分基于系列理论与假说,其中,生态位理论和中性理论最具代表性。生态位理论认为,各种物种特征的权衡和组合使其沿着决定适合度的所有变量的梯度占据特定的生态位并得以稳定共存[40-41]。基于生态位理论的群落构建认为,环境选择作用和竞争排斥等确定性因素共同塑造着群落结构。群落中性理论被用来解释群落构建过程中漂变、扩散限制等随机性过程,该理论由种群遗传学中的中性理论衍生而来,认为个体在生态水平上具有概率意义的等价性,并借鉴岛屿生物地理学理论中某个个体的死亡伴随群落外其他随机个体迁入的情形,假设局域群落具有饱和性[42-43]。中性理论由以上假设推论得到物种群落物种多度分布符合零和多项式分布,群落结构由扩散限制决定[43]。
随着高通量测序技术在环境微生物群落中的不断应用和对大尺度下环境微生物分布模式的逐步探索,越来越多的研究发现,微生物群落组成结构不能仅被环境驱动因素、种间互作关系等切实可控的因素来解释,生态随机过程在微生物多样性和群落构建方面发挥重要作用[8]。目前,生态确定性过程和随机性过程共同主导环境微生物群落构建过程的观点已被普遍接受,确定性因素与随机性因素的相对重要性成为影响群落构建过程的核心科学问题[8,44]。
VELLEND将生态过程整合成4个主要过程:物种形成(speciation)、选择(selection)、扩散(dispersal)和生态漂变(drift)[45]。之后,有学者提出使用多样化(diversification)来代替物种形成过程,如图1所示[8,46]。选择过程主要是由生态位理论所主导的生态过程,群落生物多样性主要是由不同物种在适应环境过程中的差异造成的(包括物种生存、生长和繁殖等),驱动上述差异的原因主要是非生物因素(如环境因素)和生物相互作用因素(物种竞争、协作、共生、捕食等)[8]。由于受到环境异质性和环境同质性的影响,相应的生态选择过程分为同质选择和异质选择[47]。扩散过程主要是指群落内物种主动或被动地从一个地点迁移至另一个地点并成功定殖的过程。生态漂变是由物种的随机产生、随机死亡和随机繁殖等一些特有的随机因素所产生的群落内物种差异,进而形成不同的生物多样性模式[43]。多样化指产生新的遗传变异位点的生态进化过程,是介于物种形成和物种灭绝之间的一种状态[8]。一般认为扩散与多样化过程使微生物物种多样性增加,而选择与生态漂变过程使生物多样性降低[48]。这些生态过程不是单一地作用于微生物群落构建,而是相互结合,以某一个过程为主导、多个生态过程共同发挥作用促进环境微生物群落生物多样性模式形成。
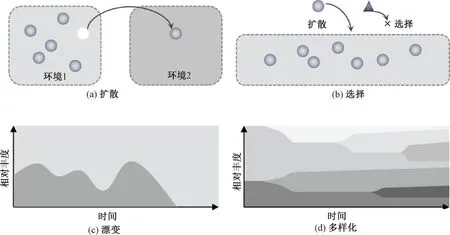
图1 影响微生物群落构建的4个生态过程[8,46]Fig.1 Four ecological processes affecting the construction of microbial communities[8,46]
2.2 湖泊微生物群落构建机制分析方法
目前,在湖泊生态系统中研究微生物群落构建机制的分析方法主要有统计分析法、模型推断法和网络分析法。
1)统计分析法
环境选择与扩散限制因素共同驱动湖泊微生物群落的动态变化和生物地理分布格局形成,研究2种过程对群落结构影响的多元统计方法主要有3种。
第一种方法是直接比较不同组间群落结构的差异,使用探索性的降维排序方法,如主成分分析(PCA)、主坐标分析(PCoA)、非度量多维标度(NMDS)和去趋势对应分析(DCA),以及统计检验方法,如置换多元方差分析(PERMANOVA)和相似性分析(ANOSIM)。此类方法可以揭示多种选择作用造成的群落结构差异,但难以评价随机性过程的重要性。第二种方法是探究群落结构和环境变量之间的关系,选择解释性的数据分析方法,如冗余分析(RDA)、典范对应分析(CCA)和Mantel检验[49]。若环境变量能够解释很大比例的群落结构变化,则可以证明选择过程的重要性,反之则可能表明随机过程的重要性。然而,选择作用和扩散限制可能产生相似的距离衰减模式[50-52]。因此,第三种多元统计方法扩展到方差分解分析(VPA),确定环境选择和空间效应对群落结构差异的相对重要性[49,53]。同时,该方法量化了环境变量与空间距离之间的相互作用,表征了环境异质性造成的影响。其余未解释部分的变化可能由未测定的环境变量、生物相互作用、物种分化、漂变和人为误差引起[7,54-55]。
2)模型推断法
模型推断的方法包括中性群落模型(NCM)和零模型(null model)。
中性群落模型最早应用于大型动植物群落构建机制研究,而后被推广应用于探讨微生物群落构建过程。目前普遍使用的Sloan模型沿用了早期中性模型的符号和核心概念,改进之处在于允许物种有竞争优势或劣势,并将模型转化为连续的形式[42]。中性模型从随机性或确定性层次评估群落构建过程,假设群落构建完全由偶然性和扩散驱动,通过拟合观测得到群落结构数据,得出拟合程度(R2)、群落规模(N)和迁移率(m)等参数,由拟合优度判断群落构建中随机性过程的强度,由群落迁移率的大小判断物种受到扩散限制的程度[56-58]。采用该方法对太湖光合微生物的研究发现,在短至3 d的时间尺度上,随机过程主导光合微生物群落构建过程[59]。
零模型方法通过比较随机生成群落与观测群落的差异,推断关注的生态过程是否对观测群落产生显著影响。基于零模型的标准化随机比例(NST)方法仅在随机性或确定性的层次划分生态过程,但能够量化确定性和随机过程在不同群落构建中的相对重要性。如果一组群落的NST值主要为0.5~1,则认为该组群落由随机性过程主导,反之,若NST值主要为0~0.5,则认为该组群落由确定性过程主导。该方法基于模拟生态数据,具有更高的准确度和检测精度。有研究使用NST指数衡量浮游细菌群落、底栖细菌群落在冬季、夏季不同的群落构建机制,结果发现在2个季节中,浮游细菌群落受到随机性过程的影响大于底栖细菌群落[60]。
基于零模型的Stegen生态框架在判断随机性或确定性过程的基础上,将确定性过程划分为同质选择(HoS)和异质选择(HeS),将随机性过程划分为同质扩散(HD)、扩散限制(DL)和无主导过程(UD)[55]。该框架结合系统发育研究和分类学研究内容,主要使用基于系统发育β多样性指数的β最相邻物种指数(βNTI)和基于Bray-Curtis距离的Raup-Crick指数(RCBray)进行划分,如ZHANG等使用该方法判断出环境选择过程是主导青藏高原地区湖泊微生物群落构建的主要生态过程,特定的局部环境条件决定了微生物群落生物多样性的形成[61]。在以上方法的基础上,iCAMP执行基于系统发育分箱(bin)的零模型分析,以量化不同生态过程的相对重要性。iCAMP框架同样将群落构建划分为5个过程,将无主导过程延伸到漂变等过程,并可以进行群落组内各过程强度的比较。首先将观察到的分类群根据其系统发育关系分为不同的bin,随后使用净种间亲缘关系指数(βNRI)对每个bin的系统发育多样性进行零模型分析,并使用修正的Raup-Crick指数分析β多样性[62]。相关研究采用此方法将观察到的物种分组到92个bin中,再进行零模型分析,证明了人类活动加剧会对某些微生物物种选择过程产生同质化效应[63]。
基于零模型的PER-SIMPER方法和扩散-生态位连续体指数方法(DNCI)将生态过程划分为生态位/扩散过程[64]。PER-SIMPER方法由GIBERT等基于相似性百分比(SIMPER)提出,计算扩散模型、生态位模型、混合模型的E值分布,定性推断主要的生态过程,扩散/生态位模型E值分布较低,则对应生态过程主导群落构建,混合模型E值分布较低,则表示2个过程共同影响群落构建。主要原理为对经验数据进行重排获得3种不同的零分布,并基于SIMPER分析,计算经验SIMPER和3种不同的零SIMPER。随后,对于每个零模型,计算零SIMPER与经验SIMPER之间的平方偏差之和的对数(E),该方法仅能用于生态过程的定性判断[65]。VILMI等在此基础上计算了效应量差值,提出了扩散-生态位连续体指数(DNCI),以DNCI值的正或负推断群落构建是由生态位主导还是扩散过程主导,以其绝对值大小表示驱动因素的强度,可以进行群落组间的比较[66]。相关研究使用该方法证明了扩散过程在多个空间尺度上对浮游生物群落组成相似性具有不同的影响[67]。以上基于模型推断的分析方法由于构建原理不同,每种方法都有其适用性(表1)。使用单一方法进行分析得到的结论往往不能准确反映群落构建中各生态过程的相对重要性,需要根据具体的研究问题选择多种方法综合分析。此外,统计分析和模型推断方法对于推断生物相互作用的影响程度和作用机制能力较弱,需要结合生态网络分析方法进行解析。

表1 模型推断方法比较Table 1 Comparison of model extrapolation methods
3)网络分析法
在复杂的生态系统中,物种之间具有普遍的联系,将物种之间潜在的相互作用关系用网络模型简化形成生态网络[69]。由于传统微生物识别技术难以汇集生态系统水平上的所有物种数据,微生物生态网络大多基于DNA或蛋白质分子标记技术获取组学数据构建而成,被称为分子生态网络(MENs)[70-71]。构建生态网络的方法众多,常见的生态网络基于差异度、相关性、多元回归、概率图模型等方法构建而成。网络构建方法的选择应综合考虑物种多样性、物种组成、数据类型(截面/时间序列数据)以及研究目的[72]。网络分析有助于揭示环境选择、生物相互作用的影响以及群落构建过程中的优先效应。但如果在所选择的研究尺度上群落构建过程完全由随机性因素主导,网络就不再能提供有效信息[73-74]。因此,为避免建立不具有解释性意义的网络,需要预先从群落生态的角度判断构建过程的随机性或确定性[75]。
研究环境驱动因素的第一种方法是直接将环境因子作为附加节点,计算与微生物类群之间的关联,探究对微生物群落组成的调控机制[75]。通过建立物种-环境因子网络,ZENG等揭示了浮游细菌和沉积物细菌群落网络结构改变受不同环境因子影响,浮游细菌群落中物种相互作用主要受总磷和叶绿素a影响,而沉积物细菌群落网络连接主要由硝态氮、总磷和亚硝酸盐氮决定[22,76]。第二种方法是通过逐个样本聚类或根据关键变量(如水深或pH)将样本分组,并分别为每个样本组建立网络,不同环境条件下微生物群落内呈现出不同的物种相互作用关系。如MO等使用不同盐度水平下的真核微生物生物群落的扩增子测序数据构建网络,发现真核微生物网络模块结构和特性对盐度变化敏感,表明环境异质性可诱导微生物生态网络模块化[18]。
生态网络综合了群落中物种的相互作用关系,物种之间会表现出共同出现、彼此排斥或无相互作用等情形。这些情形除了代表潜在的相互作用关系或信息传递关系,也可能与生态过程有关。物种的共现可能是由于共同的环境选择或扩散限制,物种的互斥可能是生态位分化或空间隔离造成的[77-78]。基于差异度/相关性的方法建立的网络可以揭示物种之间的成对关系,在网络中由连接2个节点的边表示。相关研究解析了湖泊浮游细菌的种间关系,网络结构呈现出物种之间显著负相关的共同特征,表明竞争在浮游细菌群落构建机制中的作用[63]。基于回归和关联规则方法建立的网络可以预测更为复杂的生物相互作用关系,如一个微生物类群受到多个其他类群的影响,推断出的复杂关系在网络中表示为定向连接2个以上节点的边[79]。使用域内生态网络可以分析域内物种之间的相互关系,而跨域生态网络方法(IDEN)关注域间的相互关系而忽略域内物种之间的相互关系[80-81]。XUE等使用跨域生态网络分析方法探究了蓝藻水华的发生对不同微生物类群之间相互作用关系的影响,发现在水华消退后,水体中营养物的减少导致营养级之间关联增强,使网络复杂性显著增加[82]。
网络分析还有助于认识群落构建过程中随机因素的历史偶然性。以概率图模型为构建原理,SAZAL等将贝叶斯网络和共现网络结合实现了仅从微生物丰度数据推断定殖顺序,这一研究检测了物种加入群落的特定顺序和时间,即群落构建过程中的优先效应[83-84]。其中,贝叶斯网络用节点表示多维变量,边表示节点之间的条件依赖关系,这种数学关系在特定情况下近似于因果关系[75]。构建的网络包含有向和无向的边,当变量高度相关时,边显示为无向或双向。该方法可以根据边的方向推断微生物组中微生物类群之间的定向关系,并区分直接和间接的条件依赖关系,但该方法计算成本高,不适用于大样本量数据[76]。
2.3 湖泊微生物群落构建机制影响因素
2.3.1 尺度效应
确定性过程、随机性过程对微生物群落构建的相对重要性与研究尺度密切相关[85]。空间尺度上,在同质环境条件中,局域尺度群落构建过程主要是随机的,服从中性模型所做的预测,而在最大尺度上扩散限制起主导作用。在异质环境条件中,在最小空间尺度上可能会发生环境对群落多样性分布的选择作用,在局域尺度上,物种迁入和迁出对群落动态产生更大的影响,生物相互作用会对物种进行选择,随空间尺度进一步增大,物种选择主要由气候因素驱动(如降水和温度),扩散限制过程也将变得越来越重要[86]。相关研究为该理论提供了依据,如LIU等对南极、北极、青藏高原地区的湖泊微生物群落构建机制分析发现,在区域尺度上,微生物群落结构与当地气候因素明显相关,说明群落构建过程主要由确定性选择过程驱动,而在跨区域尺度上扩散限制起到更重要的作用[87]。时间尺度上,不同时间尺度内微生物群落构建过程往往由不同的生态过程主导,可能是由于生态过程在长时间跨度内发生方向与强度的变化,或是由于时间本身可以作为群落构建的驱动因素(如随机或确定性事件强度随着时间积累而增加,使微生物群落受到更多随机或确定性过程的影响[88])。
2.3.2 非生物因素
微生物群落构建与湖泊水动力条件有关,如孤立湖泊与连通湖泊具有不同的水文水动力条件,导致这2类湖泊在水质特征、生境格局、水生生物迁移扩散、物质循环过程等方面存在差异,微生物会采取不同的群落构建模式[68,89]。有关研究对比了孤立、连通湖泊浮游植物物种多样性和功能多样性模式,发现环境因素(如电导率、浑浊度等)与连通性湖泊浮游植物多样性格局的形成相关性更强,表明确定性过程对连通性湖泊群落构建有影响[90]。孤立湖泊中稀有微生物类群对环境因子更为敏感,而在相互连通的湖泊中,稀有类群和丰富类群可能对扩散过程或环境变化做出相似的响应[68,91]。湖泊环境条件(如pH、溶解氧含量和营养状态等)对微生物群落结构有直接作用[90]。环境条件变化的剧烈程度影响着确定性和随机性过程的相对重要性。对其他生态系统的研究表明,在环境条件发生剧烈或突然变化时,确定性过程占主导地位,而在轻度和渐进变化中,随机过程占主导地位[92]。
2.3.3 生物因素
由于不同的微生物类群对环境和空间变量具有不同的响应模式,不同相对丰度和出现频率的物种群落构建过程可能存在差异[7]。基于物种相对丰度将微生物划分为丰富、稀有类群,不同丰度的类群对确定性和随机性过程的敏感程度不同。此外,由于微生物对环境波动的适应能力不同,生态位宽度的分布也会影响群落构建过程,对于西藏湖泊沉积物中泛化种与特化种的环境适应性研究表明,确定性过程主导泛化种群落构建,而特化种的群落演替更多由随机性过程解释[93]。
3 总结与展望
在基础理论方面,用生态位理论和中性理论解释群落构建过程,是采取从现象到理论机制的研究思路,是从群落格局出发推测群落的形成过程,2种理论在不断更新和发展的过程中整合形成了多种假说。生态群落理论框架从群落的形成过程出发,认为选择、漂变、成种和扩散4种高层级过程在群落构建中都发挥作用。未来的微生物群落构建机制研究需要进一步结合当代物种共存理论,将多时空尺度的生态过程纳入统一的理论框架,揭示不同时空尺度下微生物群落特征和生态学模式。
在技术方法方面,目前对微生物群落的调查多依赖于高通量测序技术,该方法存在偏好度、精度等测量误差,对随机性和确定性的判断与量化仅是一种统计结果,不能完全作为群落多样性形成与维持过程的最终结论。普遍使用的零模型方法也面临着多方面的挑战,从机理解释上看,零模型作为统计学的随机模型,本身不具备生态学的机理,忽略了生物属性及其相互作用,更无法区分混合的生态过程;从逻辑推理上看,零模型没有设置对立的零假设,随机作用只是物种分布符合零模型预测的一种可能,多种确定性作用也会使物种产生随机的分布模式;从实际应用上看,零模型方法算法和相似性度量方法的选择可能导致随机性估计的结果出现较大差异,结果的可靠度往往取决于具体生态问题和群落。因此,解析群落的构建机制仍需要设计控制实验进行验证,或进行调查研究及因果分析。
在影响因素方面,对确定性过程的研究中,环境过滤、生物相互作用、物种分化等过程的相对贡献率仍不清楚,未来研究应注重构建多种过程耦合关系模型,加强确定性过程的定量分析研究;对随机性过程的研究中,虽然随机过程本身难以解析,但影响随机过程的因素值得关注,未来研究应集中探讨在何时、何种条件下随机过程对微生物更加重要。此外,还需要将湖泊微生物群落时空分布规律与湖泊生态系统服务结合,解析微生物组-环境-生态系统的关联,最终服务于生态问题的评估、预防、调控和管理。根据环境因素和空间因素对微生物群落多样性分布的影响,可以得到微生物群落的物种分布模型,在此基础上应尽可能丰富湖泊微生物样本数据集,涵盖多种湖泊类型,开展时间序列上的研究,在全球背景下探究湖泊微生物群落的时空演变规律,将有助于保护湖泊微生物多样性及其生态功能。
