广藿香PcGA2ox1基因克隆、VIGS载体构建和表达分析
2024-01-01严雅玲曾晴严寒静何梦玲张宏意


关键词:广藿香;赤霉素2-氧化酶(GA2ox);基因克隆:病毒诱导基因沉默(VIGS);表达分析
广藿香[Pogostemon cablin(Blanco)Benth.]为唇形科刺蕊草属植物,以干燥地上部分人药,原产于东南亚菲律宾、马来西亚等热带地区,宋朝时期传人我国并在南方广泛种植,道地产区主要分布在两广、岭南等,故而有广藿香之称。广藿香少有开花,很难结果,因而主要采用扦插方式繁殖,但扦插繁殖慢且易引起种质退化,难以培育出优质品种。因此,通过促进侧枝发育增加地上部分的比重,从而提高广藿香产量,对选育品质优良的广藿香意义重大。
赤霉素(gibberellins,GAs)是一种重要的植物内源激素,调控植物生命周期的各个阶段,在根茎叶伸长、侧枝发育、花期调控、种子休眠等方面发挥作用。植物体内的GAs分为有活性和无活性两类,其中GA1和GA4为有活性类。GA20x(gibberellin 2-oxidases)作为赤霉素代谢途径中的一个关键酶,可以将活性赤霉素(GAI和GA4)降解为无活性状态,限制有生物活性GAs的含量,从而调节植物发育,如在拟南芥中阻止种子萌发、延迟无性繁殖、改变花期、限制主茎和侧芽的伸长等。一般而言,植物体内GA2ox过表达会使活性GAs含量降低,从而使植株表现为矮化,而将其沉默,则会促进茎的伸长和植株生长。目前已有众多植物GA2ox基因被成功克隆并进行了相关研究,如GA2ox基因在油菜籽、早熟禾、油茶等植物中过表达呈现出矮化表征,生物量减少、开花延迟、叶片颜色呈深绿色等:相反,在烟草中沉默GA2ox可以显著促进烟草生长和提高纤维产量。目前GA2ox基因在烟草、拟南芥、番茄等植物中已有研究,但未见广藿香GA2ox基因的相关研究报道,为此,本研究基于本课题组前期获得的广藿香转录组数据,对其GA2oxl基因进行克隆和生物信息学分析,以探寻GA2ox基因在广藿香中的作用,为后续研究矮化广藿香、促进侧枝发育和增加产量等提供有力依据。
病毒诱导基因沉默(VIGS)是一种利用植物的病毒防御机制来抑制特定侵袭性病毒转录物的基因沉默方式,是研究植物基因功能的强有力工具,与传统的基因功能分析方法相比,VIGS具有沉默单个或多个基因家族成员的潜力,已被应用于众多研究领域,如在木薯、大豆、番茄等植物中利用VIGS技术进行抗病性研究:Wang等发现通过VIGS敲除GA中主要DELLA基因,可显著增强棉花茎木质部和韧皮部次生细胞壁的形成:利用VIGS沉默百合花药的赤霉素上调基因LoMYB65,可导致花粉发育不正常,花粉量减少等。为探寻GA2ox基因在广藿香中的沉默效应,本实验采用烟草脆裂病毒载体对广藿香叶片进行VIGS侵染,以降低GA2oxl基因的表达水平,探究其对广藿香表型的影响,以期为广藿香优良品种选育奠定理论基础。
1材料与方法
1.1供试材料
组培苗培养:摘取经广东药科大学中药学院严寒静教授鉴定为唇形科刺蕊草属植物广藿香Pogostemon cablin(Blanco)
Benth.的叶片接种于丛芽诱导培养基(MS+0.2mg/L 6-BA+0.1 mg/LNAA)上培养40d,然后选取生长健壮、长势一致的不定芽接种于生根培养基(1/2MS)上培养60d,统一采用日光灯为光源,在25℃恒温培养。
将生根培养60d的组培苗移栽水培两周,然后移栽至混合土壤(普通土与营养基质的体积比为1:1)继续培养60d。选取生长良好的广藿香叶片液氮速冻后用于提取总RNA;选取生长良好的广藿香植株进行VIGS实验,取第2.3对叶用于沉默基因表达量分析;选取广藿香的根、茎(第2对叶与第3对叶之间)、叶(第2或第3对叶)、芽(顶芽),液氮速冻后存于-80℃冰箱,用于组织特异性表达分析。
所用菌株DH5a、农杆菌GV3101感受态细胞,购自北京庄盟国际生物基因科技有限公司:EcoRI和Xhol快切酶购于宝日医生物技术(北京)有限公司;pTRV1、pTRV2载体为实验室留存。
1.2实验方法
1.2.1总RNA提取及cDNA合成根据不同实验要求选用广藿香不同组织部位材料,使用天根植物RNA prep Pure Plant试剂盒提取广藿香总RNA,用超微量紫外分光光度计UV2450检测总RNA纯度和浓度,并使用琼脂糖凝胶电泳法对其完整性进行检验。使用TaKaRa生物公司的Pri-meScript RT reagent Kit with gDNA Eraser试剂盒反转录合成广藿香cDNA,于-20℃贮存备用。
1.2.2广藿香PcGA2oxl基因的克隆
基于课题组前期获得的转录组测序数据,使用PrimerPremier5.0软件设计特异引物(表1)。以反转录得到的广藿香cDNA为模板,设定PCR反应体系:cDNA1uL,上游引物PcGA20xl-F,下游引物35个循环;72℃延伸2 min。产物经胶回收后检测纯度和浓度,采用北京庄盟国际生物基因科技有限公司的克隆载体One step ZTOPO-Blunt/TA进行连接并转化大肠杆菌DH5a感受态细胞,将获得的阳性单菌落送至北京擎科生物科技有限公司测序,检查该序列是否为目标序列。
1.2.3PcGA2oxl的生物信息学分析
利用相应的在线分析软件(表2)对PcGA2oxl基因进行生物信息学分析,利用BLAST对不同植物的GA2ox氨基酸序列进行同源比对,运用MEGA-X软件利用NJ法对不同植物GA2ox基因序列进行系统进化树分析。
1.2.4VIGS实验基于克隆得到的PcGA2oxl基因CDS序列,选取其中第344~558bp片段,利用SnapGene软件设计引物(表1),引入EcoR I和Xhol两个酶切位点,扩增PcGA2oxl基因215 bp片段,构建VIGS载体。PCR扩增体系50PCR反应条件同1.2.2,回收得到目的基因片段。
使用TaKaRa QuickCut限制酶进行pTRV2载体上的EcoRI和Xhol双酶切,37℃反应15min,切胶回收得到酶切产物,检测浓度和纯度后,将酶切产物与目的基因用庄盟生物的SE无缝克隆试剂盒进行无缝克隆,转化DH5ce感受态细胞,测序正确后提取得到pTRV2-PcGA20xl质粒。
分别将pTRV1、pTRV2和pTRV2-PcGA20xl质粒转化农杆菌GV3101感受态细胞,涂布在LB培养基平板(含50mg/LKan,50 mg/LRif)上,恒温培养箱28℃倒置培养24h,挑取单克隆菌经PCR鉴定,随即活化扩大培养阳性克隆菌,培养至菌液OD600值为1.0~1.5,重悬后用侵染液(10mmol/L MES、10mmol/L MgCl2、200umol/L AS)沉淀悬浮,于28℃恒温培养箱遮光静置3h,然后将pTRV1侵染液分别与pTRV2和pTRV2-Pc-GA2oxl侵染液按1:1混合,混匀后从叶背面注射侵染广藿香第2对、第3对叶,于28 0C恒温培养两周后,按照不同组别采集广藿香叶片用于基因表达量测定。
1.2.5PcGA20xl基因表达量测定使用PrimerPremier 5.0软件设计VIGS沉默及组织特异性表达的qRT-PCR引物(表1),以18SrRNA为内参基因,以反转录得到的cDNA为模板,使用SYBRGreen Premix Pro Taq PCR试剂盒和荧光定量PCR仪CFX96进行实时荧光定量PCR。使用20扩增体系:TB Green⑩Premix Ex TaqⅡ(TliRNaseH Plus)(2x),上、下游引物各0.4,模板cDNA 1.6 VL,双蒸水(ddH20) 7.6。扩增程序:95℃ 30s;95℃ 5s,60℃ 30s,40个循环。每个样品3次重复,采用2法计算VIGS沉默及不同组织部位的基因表达量,使用Prismchs软件进行分析绘图。
2结果与分析
2.1PcGA20xl基因克隆
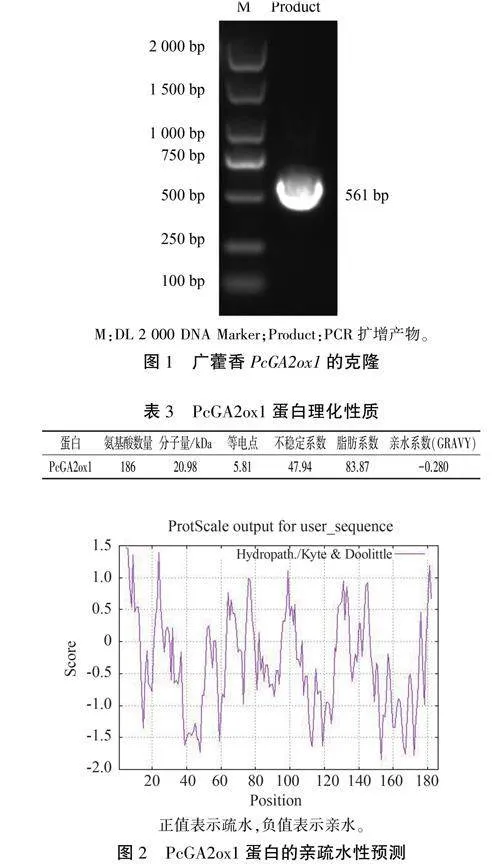
根据前期获得的转录组数据,克隆得到Pc-GA20xl的CDS序列,并设计了特异性引物;以广藿香cDNA为模板进行PCR扩增,通过1%琼脂糖凝胶电泳检测,观察到在500 bp左右位置形成了清晰的条带(图1)。经过对比分析,目的片段与预期大小相符,由SnapGene测序验证确认所得序列为完整的561bp的正确CDS序列。
2.2PcGA20xl基因的生物信息学特征
2.2.1PcGA2oxl蛋白理化性质及亲疏水性分析
ProParam预测结果(表3)显示,PcGA20xl蛋白的分子式为C930 H1455 N255 0278 S10.分子量为20.98kDa,脂肪系数为83.87,为亲水性的不稳定弱酸性蛋白;酸性氨基酸残基(Asp+Glu)总数为22,碱性氨基酸残基(Arg+Lys)总数为19。ProtScale分析结果(图2)表明,PcGA20xl氨基酸多肽链的亲水性最大值为1. 467,最小值为-1.844.主要表现为亲水性,与ProParam预测结果一致。
2.2.2PcGA2oxl蛋白跨膜结构、信号肽及亚细胞定位预测TMHMM分析结果表明,PcGA20xl蛋白不存在跨膜结构。SignalP6.0未预测到信号肽裂解位点,推测PcGA2oxl蛋白不具有信号肽,属于非分泌类蛋白。WoLF PSORT亚细胞定位预测结果表明.PcGA2oxl编码蛋白主要定位在细胞核中。
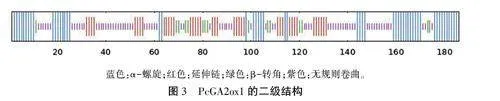
2.2.3PcGA2oxl蛋白二级结构结果(图3)显不,PcGA2oxl蛋白的二级结构由55个a-螺旋、39个延伸链、15个转角、77个无规则卷曲组成,占比分别为29.57%、20.97%、8.06%、41.40%。
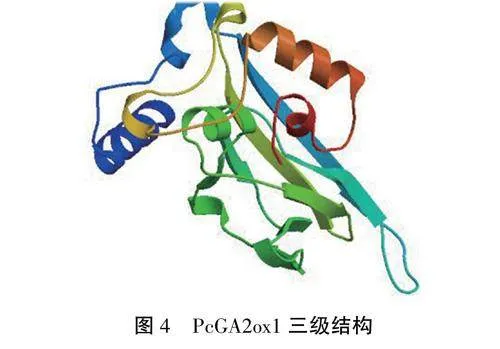
2.2.4PcGA2oxl蛋白三级结构利用SWISS-MODEL在线分析平台构建了PcGA2oxl蛋白可能的三级结构模型(图4),该模型的GMQE值为0.94,其氨基酸序列与珙桐(Nyssaceae)的模型序列相似度高达83.33%,表明对PcGA2oxl蛋白三级结构的预测结果较为可靠。
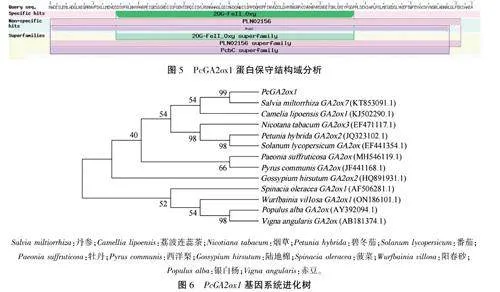
2.2.5PcGA20xl保守结构域及系统发育分析在NCBI的CDD(Conserved Domain Database)数据库搜索上传的PcGA20xl氨基酸序列,发现其具有GA20x家族保守的20G-FeⅡ_Oxy结构域,表明其属于GA20x基因家族(图5)。
运用MEGA -X软件的NJ法建立不同植物GA20xl基因序列进化树,结果(图6)显示,Pc-GA2oxl与丹参GA2ox7同属于一小支,相似度最高,达到99%,亲缘关系最近;与荔波连蕊茶、烟草、番茄、碧冬茄的同源性也达到50%以上。
2.3病毒诱导PcGA2oxl基因沉默
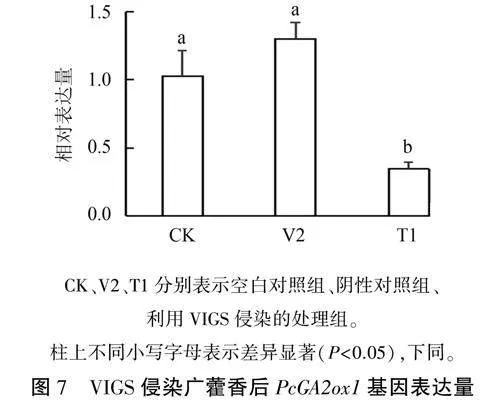
pTRV2载体经EcoRI和XhoI双酶切后与PcGA2oxl同源臂PCR切胶回收产物连接,获得pTRV2-PcGA2oxl重组质粒,热激法转化农杆菌GV3101,用VIGS法侵染广藿香。被侵染的广藿香培养两周后,取叶片,通过qRT-PCR分析Pc-GA2oxl表达量的变化。以未侵染的叶片为空白对照,注射pTRV2空载的叶片为阴性对照。结果(图7)显示,被侵染的广藿香叶片中PcGA2oxl表达量分别低于空白对照组和阴性对照组71%和83%,差异显著,沉默效果明显:空白对照组与阴性对照组之间没有显著差异,表明pTRV2空载不能影响广藿香叶片GA2oxl基因的表达水平,说明GA2oxl基因在广藿香中成功沉默。
2.4PcGA2oxl基因组织特异性表达分析
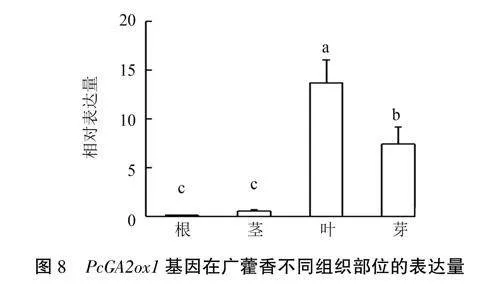
选取生长时间相同、长势一致的广藿香植株,对其根、茎、叶和芽中的PcGA2oxl基因表达量进行检测,结果(图8)显示,PcGA2oxl基因在各组织中均有表达,但表达水平存在差异,以叶中的表达量最高,根中的表达量最低。
3讨论
GAs的生物合成在高等植物生长发育过程中不可或缺,GAs代谢过程中有多种酶,这些酶会影响生物合成与分解之间的平衡,能维持特定发育阶段生物活性水平,实现植物生长发育正常进行。GA2氧化酶使GAs羟基化,导致活性赤霉素钝化为非活性形式,因此可以调节植物体内赤霉素含量,影响植物生长过程。GA2ox是由多基因家族编码的双加氧酶,可分为三类,I类和Ⅱ类都作用于C19-GAs,I类使其失活,Ⅱ类使其前体失活;Ⅲ类作用于C20-GAs,使其2-羟基化。根据生物信息学分析及构建系统发育树可知,广藿香PcGA2oxl基因与丹参的GA2ox7相似度最高,达到99%,亲缘关系最近;与荔波连蕊茶、烟草、番茄、碧冬茄也聚在一支,同源性达到50%以上。推测PcGA2oxl基因同上述植物的GA2ox基因相似,也属于I类,催化C19-GAs失活,从而使具有生物活性的GA1和GA4失活。
研究表明,GA2ox基因在植物生长发育过程中存在组织特异性表达差异。大麦HvGA2oxl和HvGA20x3在成熟种子的节间和胚乳中高表达,HvGA2ox6主要在胚中表达:丹参SmGA20xl在花中的表达较高,SmGA20x4在根中的表达较高,SmGA2ox7在叶中高表达,SmGA2ox10在茎中高表达:而荔波连蕊茶CIGA2oxl基因在嫩枝和二年生茎段中的表达量最高,在幼叶中的表达量最低;万寿菊GA2ox基因在茎及叶中的表达量较高.在种子中的表达量较低。表明不同物种GA2ox基因虽同源性较高,但功能存在差异。本研究结果表明,Pc-GA2oxl基因在广藿香的根、茎、叶、芽中均有表达,以叶中的表达量最高,芽中的次之,根中的表达量最低,且其表达在根与叶、茎与叶、根与芽、茎与芽间均存在显著性差异,具有一定的组织特异性。由此推测PcGA2oxl基因主要参与广藿香叶、芽的生长发育。
由于易用性好和产生表型所需的时间短,VIGS被广泛用于植物遗传学中的基因沉默。在广藿香中,靳巧春等利用VIGS技术沉默广藿香醇合酶PcPTS基因,对其功能进行了鉴定,并初步建立了广藿香的VIGS体系:杨豫章通过VIGS技术有效沉默了广藿香基因,一定程度上影响了广藿香中百秋李醇的生物合成:王小兵利用TRV病毒载体沉默广藿香内源PcPDS基因,使其表达量明显下降,且观察到叶片白化表型。本研究利用VIGS技术沉默广藿香PcGA2oxl基因,qRT-PCR结果显示,沉默两周后,PcGA2oxl基因表达量与阴性对照相比显著下降83%,与空白对照相比显著下降71%。表明PcGA2oxl基因在广藿香中被有效沉默。
已有研究表明,VIGS沉默技术用于GA20x基因上可促进茎节生长、新枝发育等,如:Joanna等利用VIGS技术沉默碧冬茄“梦幻蓝”的GA2ox基因,基因沉默植株会表现出茎伸长:Li等在紫薇垂枝性状机制研究时发现,VIGS诱导可降低GA信号传导水平,使紫薇长出新枝。但本研究仅采用烟草脆裂病毒载体TRV使用注射法瞬时侵染广藿香叶片,且调查时间短,尚未发现明显表型变化,后续将延长侵染时间,观察广藿香生长的变化,并通过含量测定和激素水平分析等进一步探寻VIGS沉默PcGA2oxl基因是否对广藿香生长发育产生影响。
4结论
本研究首次从广藿香中克隆出PcGA2oxl基因,全长为561bp,编码186个氨基酸,为亲水性蛋白,具有GA2ox基因家族的保守结构域,主要定位在细胞核。PcGA2oxl基因在广藿香中的表达存在组织特异性,在叶中的表达量最高:利用VIGS技术成功沉默了PcGA2oxl基因,初步验证了该基因的功能。本研究结果可为今后深入研究PcGA2oxl基因的功能、选育矮化广藿香等奠定基础。
