基于转录组测序分析溶解氧对四指马鲅幼鱼鳃组织的影响
2024-01-01林欣谢希尧区又君李加儿



摘要:【目的】基于转录组测序分析溶解氧对四指马鲅(Eleutheronema tetradactylum)幼鱼鳃组织的影响,探究四 指马鲅应对低氧胁迫的适应机制,为四指马鲅的抗逆性研究提供参考依据。【方法】以四指马鲅幼鱼为试验材料, 设3种溶解氧含量处理,常氧组(CG)6.33±0.15mg/L、中度低氧组(OC4)3.99±0.18mg/L和重度低氧组(OC2)2.05±0.15mg/L,处理29h,采集鳃组织进行转录组测序,利用DEGseq筛选不同比较组的差异表达基因(DEGs),使用 GOseq进行GO功能注释分析,利用KOBAS进行KEGG信号通路富集分析,采用STEM对DEGs进行表达趋势分析。 【结果】OC4 vs CG比较组有86个DEGs,其中28个上调表达,58个下调表达;OC2vs CG比较组有2018个DEGs,其中 740个上调表达,1278个下调表达;OC4 vs OC2比较组有171个DEGs,其中68个上调表达,103个下调表达。GO功能注释分析结果显示,DEGs主要涉及代谢过程、有机物生物合成、细胞氧化还原平衡、细胞内、染色质、核糖体结构成分和催化活性等功能条目。KEGG信号通路富集分析结果显示,DEGs主要富集在蛋白酶体、DNA复制、脂肪酸降解、磷 酸戊糖途径、氧化磷酸化、谷胱甘肽代谢、赖氨酸降解和核糖体生物发生等信号通路;其中pacl、lsm7、mcm6、mcm2和 mdm2等基因富集在蛋白酶体、剪接体、DNA复制和细胞周期等信号通路,与四指马鲅幼鱼鳃组织蛋白合成有关,随 着溶解氧含量下降,其表达量呈下降趋势;shmt2、hsd17b10、sucla2和glol等基因富集在磷酸戊糖途径信号通路,与四 指马鲅幼鱼鳃组织能量代谢有关。【结论】四指马鲅幼鱼鳃组织蛋白质合成能力和能量代谢活动均随着溶解氧含量下 降而下降,溶解氧含量下降显著影响细胞周期、DNA复制、脂肪酸降解和糖代谢等信号通路。
关键词:四指马鲅;幼鱼;鳃;溶解氧;转录组
中图分类号:S917.4
文献标志码:A
文章编号:2095-1191(2024)03-0660-10
Effects of dissolved oxygen on gill tissue of juvenile Eleutheronema tetradactylum based on transcriptome sequencingLIN Xin1.2, XIE Xi-yao1.2, OU You-jun1*, LI Jia-er1
(1South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences/Key Laboratory of South China Sea Fishery Resources Exploitation amp; Utilization, Ministry of Agriculture and Rural Affairs, Guangzhou, Guangdong 510300, China; 2College of Fisheries and Life Science,Dalian Ocean University, Dalian, Liaoning 116023, China)
Abstract: [Objective] Transcriptome sequencing-based analysis of the effects of dissolved oxygen on gill tissues of juvenile Eleutheronema tetradactylum was conducted to investigate the adaptive mechanisms of E. tetradactylum in response to hypoxia stress. To provide a reference basis for research on the resistance of E. tetradactylum. 【Method]JuvenileE. tetradactylum were used as experimental materials, and three dissolved oxygen content treatments were set up, normoxic group (CG) 6.3340.15 mg/L, moderately hypoxic group (OC4) 3.9940.18 mg/L, and severely hypoxic group (OC2) 2.05+0.15 mg/L. The treatments were carried out for 29 h, and gill tissues were collected for transcriptome sequencing. Differentially expressed genes (DEGs) from different comparison groups were screened using DEGseq, GOfunctional annotation analysis was performed using GOseq, and KEGG signaling pathway enrichment analysis was performed using KOBAS, trends of DEGs were analyzed by STEM. 【Result】The OC4 vs CG comparison group had 86 DEGs, of which 28 were up-regulated and 58 were down-regulated; the OC2 vs CG comparison group had 2018 DEGs, of which 740 were up-regulated and 1278 were down-regulated; and the OC4 vs OC2 comparison group had 171 DEGs, of which 68 were up-regulated and 103 were down-regulated. The results of GO functional annotation analysis showed that DEGs were mainly involved in functional terms such as metabolic processes, organic matter biosynthesis, cellular re- dox homeostasis, intracellular, chromatin, ribosomal structural components and catalytic activity. KEGG signaling path- way enrichment analysis showed that DEGs were mainly enriched in signaling pathways such as proteasome, DNA repli- cation, fatty acid degradation, pentose phosphate pathway, oxidative phosphorylation, glutathione metabolism, lysine degradation, and ribosome biogenesis; among them, genes such as pacl, lsm7, mcmó, mcm2 and mdm2 were enriched in signaling pathways such as proteasome, spliceosome, DNA replication and cell cycle, they were related to protein syn- thesis in gill tissues of juvenile E. tetradactylum, and the expression of which showed a decreasing trend with the decrease of dissolved oxygen content; shmt2, hsd17b10, sucla2 and glol genes were enriched in pentose phosphate pathway, which were related to energy metabolism in gill tissues of juvenile E. tetradactylum. 【Conclusion ]Both protein synthesis capacity and energy metabolism activity of gill tissues of juvenile E. tetradactylum decrease with decreasing dissolved oxygen content, decrease of dissolved oxygen content significantly affects signaling pathways such as cell cycle, DNA replication, fatty acid degradation and sugar metabolism.
Key words: Eleutheronema tetradactylum; juvenile fish; gill; dissolved oxygen; transcriptome
Foundation items: “Blue Granary Science and Technology Innovation\"Project of National Key Research and Deve- lopment Program of China (2018YFD0900200); Guangdong Rural Revitalization Strategy Special Fund Seed Industry Re- vitalization Project (2022-SPY-00-013) ; Central Public-interest Scientific Institution Basal Research Fund, CAFS (2023 TD58)
0 引言
【研究意义】四指马鲅(Eleutheronema tetradac- tylum)隶属于鲻形目(Mugiliformes)马鲅亚目(Poly- nemoidei)马鲅科(Polynemidae)四指马鲅属(Eleu- theronema),俗称马友鱼(孙典荣和陈铮,2013)。主 要分布在印度、印度尼西亚、菲律宾、澳洲西部、日本 及中国沿海,在中国沿海主要分布于台湾、福建、广 东和海南等地区(潘炯华等,1991;庄平等,2006)。 四指马鲅脂肪含量高、肉质鲜美且口感细腻,是中国 沿海地区的名贵鱼类之一。近年来,四指马鲅人工 繁育技术取得突破并在广东、广西、福建、海南和浙 江等沿海地区推广养殖,已发展成为水产养殖的新 品种(区又君和李加儿,2022),具有极高的经济价值 和发展前景。但四指马鲅具有特殊生理结构,鳃盖 骨小,鳃丝面积小且疏离,氧气交换能力较差,一旦 离水很快死亡,因此水体中的溶解氧对其有重要影 响,研究四指马鲅鳃组织应对水体不同溶解氧含量 的响应机理,对揭示其应对低氧胁迫的调节机制有 重要意义。【前人研究进展】自然环境中的昼夜变化、 水温以及养殖密度等原因均会影响水体溶解氧含量(Wu,2002)。不同水域的水体溶解氧含量也存在差 别,河口水体溶解氧含量对底栖鱼类物种丰度和分 布的影响尤为明显(McClatchie et al.,2010)。当底层水体中溶解氧含量降至4mg/L以下时,会对底栖 物种产生应激胁迫,造成部分物种主动逃离低溶解 氧区域,底栖鱼类的丰度和多样性明显降低(Buch一 heister et al.,2013)。导致这种迁移的溶解氧含量存在物种特异性差异,由于不同物种对低溶解氧环 境的生理耐受性存在差异,部分鱼类可能仍留在低氧区域(Pihl et al.,1991,1992;Eby and Crowder, 2002;Craig,2012)。但在人工养殖中,低氧是养殖 水体的普遍问题,行动能力受限的养殖鱼类无法逃 离低氧区域,一直处于低氧胁迫状态下,严重时可能 会导致其死亡。因此,水体溶解氧是影响养殖鱼类 生存的关键环境因素之一,溶解氧对鱼类的生长发 育、新陈代谢和繁殖等生命过程具有重要影响(Vala- vanidis et al.,2006;吴志昊等,2011);低氧会对鱼类鳃组织造成氧化损伤,导致其生理功能受影响,能量 代谢和其他合成代谢发生紊乱,从而使鱼类出现 抗氧化应激状态(Sun et al.,2015);此外,水体中溶 解氧含量直接影响鱼卵孵化的成功率(之ak et al., 2023)。而目前对四指马鲅的研究主要集中在盐度 耐受性(谢木娇等,2016a)、胚胎发育(谢木娇等, 2016b)及消化道结构和功能细胞的发育等(谢木娇 等,2017)。【本研究切入点】对四指马鲅的相关研究 较多,但少有研究报道溶解氧对其鳃组织的影响,四 指马鲅鳃组织应对低溶解氧含量的调控机制尚不明 确。【拟解决的关键问题】以四指马鲅幼鱼为试验材料,设常氧组、中度低氧组和重度低氧组,通过转录 组测序分析四指马鲅鳃幼鱼在不同溶解氧含量下鳃 组织的基因差异表达趋势和代谢途径,分析溶解氧 对四指马鲅幼鱼鳃组织的影响,探究四指马鲅应对 低氧胁迫的适应机制,为四指马鲅的抗逆性研究提 供参考依据。
1材料与方法
1.1试验材料
四指马鲅幼鱼由中国水产科学研究院南海水产 研究所中山科技成果转化基地提供,平均全长4.5± 0.5cm,平均体质量1.5±0.5g。在玻璃鱼缸(60cm× 50cm×45cm)中不间断充氧暂养7d,养殖用水经曝 气处理,暂养期间每天换水1次,换水量1/2。暂养期间每天8:00、18:00和22:00进行饱食投喂,观察 并记录幼鱼进食情况。暂养水温27°℃,pH6.8+0.1, 盐度4%o,试验前禁食1d。
1.2试验设计和样品采集
设3种溶解氧含量处理,常氧组6.33±0.15mg/L、中度低氧组3.99±0.18mg/L和重度低氧组2.05±0.15mg/L,每组设3个平行,每平行10尾鱼,试验周 期为29h。试验期间,中度低氧组和重度低氧组通 过调节充气阀进气量,向水体充入氮气,控制溶解氧 含量。试验水温27℃,试验期间停止饲喂。试验结 束后对四指马鲅幼鱼进行活体解剖,收集鳃组织,置 于液氮中保持备用。常氧组鳃组织编号为CG,中度低氧组鳃组织编号为OC4,重度低氧组鳃组织编号为OC2。
1.3RNA抽提
在液氮中研磨四指马鲅幼鱼鳃组织样品,采用 总RNA提取试剂盒TransZol Up Plus RNA Kit(广州 迈宝生物科技有限公司)提取总RNA,用1.5%琼脂 糖凝胶电泳和超微量核酸蛋白测定仪检测其完整性 及浓度,检测合格后以提取的总RNA为模板,按照 反转录试剂盒EasyScript First-Strand cDNA Synthe- sis SuperMix(北京全式金生物技术股份有限公司)说明反转录合成cDNA。
1.4转录组测序
使用带有Oligo(dT)的磁珠富集mRNA,加入干扰剂,合成纯化双链cDNA,对纯化后的cDNA进行末端修复、加poly(A)尾并连接测序接头,选择片段大小,进行PCR扩增,构建cDNA文库。使用Agilent 2100生物分析仪[安捷伦科技(中国)有限公司]检测 构建的文库质量,利用Illumina HiSeq 2500/4000高 通量测序平台进行测序。
1.5测序数据质量控制
对原始序列(Raw reads)进行过滤,去除带测序接头的Reads,去除N含量超过10%的Reads,去除低质量(Q≤20)碱基含量超过50%的Reads,获得高质量的有效序列(Clean reads)。
1.6基因表达量分析
将Clean reads与Unigenes进行比较,使用RSEM 计算比对上每个Unigene的Reads数量,结合Unigenes 的有效长度及总比对上的Reads数量,采用FPKM对 Unigenes的表达量进行标准化处理。
1.7差异表达基因筛选与分析
利用DEGseq筛选3个比较组OC4 vs CG、OC2vs CG和OC4 vs OC2的差异表达基因(DEGs),筛选 标准为Qlt;0.05且|log2Fold Change|gt;1。采用K-means 法、SOM法和H-cluster法,以DEGs的相对表达水平值log2ratios进行聚类。使用GOseq对DEGs进行GO功能注释分析,当Qlt;0.05时,视为显著富集。利用KOBAS进行KEGG信号通路富集分析,以Qlt;0.01为标准。采用STEM对3个比较组的共同DEGs进行 表达分析。
1.8实时荧光定量PCR验证
在各比较组的共同DEGs中随机选取10个(5个上调,5个下调)进行实时荧光定量PCR验证,以 tubulin为内参基因,引物序列如表1所示。采用 2-AAa法计算基因相对表达量。将实时荧光定量PCR 验证结果与转录组测序数据进行比较分析。
2结果与分析
2.1转录组测序数据质量评估结果
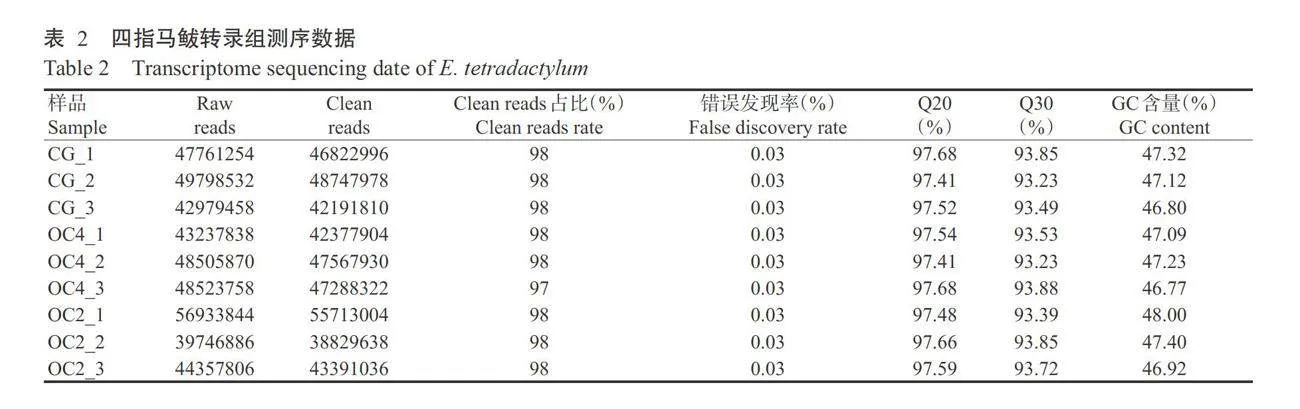
3种溶解氧含量处理四指马鲅鳃组织样品共获得412930618条Clean reads,各样品Q30均在93.23%以上,Q20均在97.41%以上,GC含量均在46.77%以 上(表2)。
2.2DEGs筛选结果
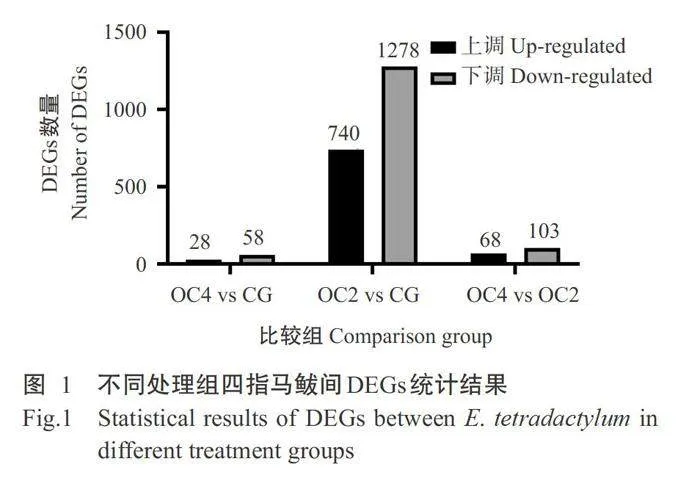
筛选不同溶解氧含量处理组间的DEGs,结果如图1所示,OC4vs CG比较组有86个DEGs,其中28个上调表达,58个下调表达;OC2vsCG比较组有2018个DEGs,其中740个上调表达,1278个下调表达;OC4 vs OC2比较组有171个DEGs,其中68个上 调表达,103个下调表达。说明不同溶解氧含量影响 四指马鲅幼鱼鳃组织基因表达。
2.3DEGs的GO功能注释分析结果
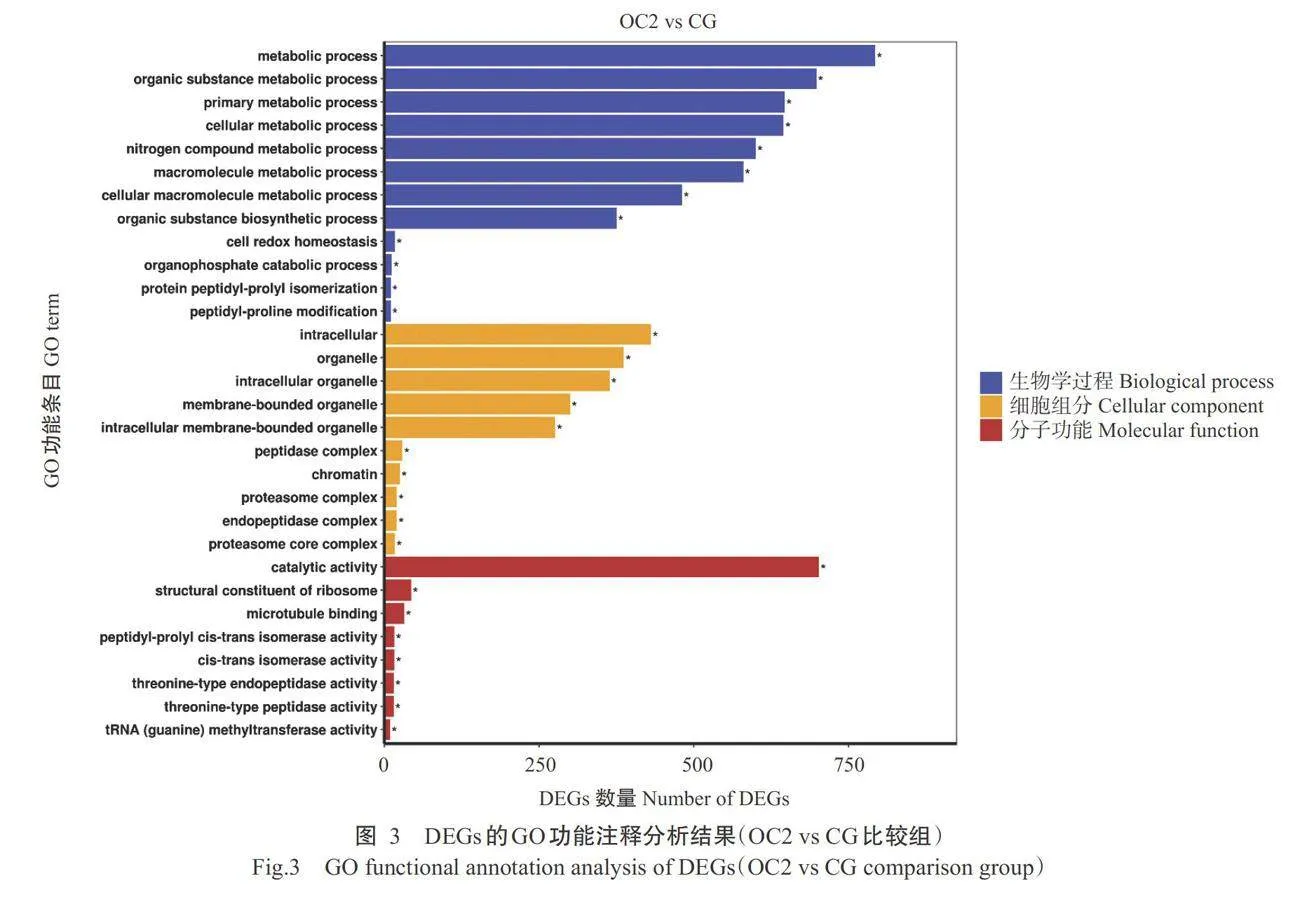
对DEGs进行GO功能注释分析,OC4 vs CG比较组的分析结果如图2所示,生物学过程中,DEGs 在脂质生物合成过程(Lipid biosynthetic process)功能条目显著富集(Plt;0.05,下同);细胞组分中,在胞 外区(Extracellular region)功能条目显著富集;分子 功能中,在氧化还原酶活性(Oxidoreductase activity) 功能条目显著富集。OC2vs CG比较组的分析结果如图3所示,生物学过程中,DEGs在有机物生物合成过程(Organic substance biosynthetic process)、代谢 过程(Metabolic process)和细胞氧化还原平衡(Cell redox homeostasis)等功能条目显著富集;细胞组分 中,在细胞内(Intracellular)、细胞器(Organelle)和染 色质(Chromatin)等功能条目显著富集;分子功能 中,在催化活性(Catalytic activity)、核糖体结构成分 (Structural constituent of ribosome)和tRNA(鸟嘌呤) 甲基转移酶活性[tRNA(guanine)methyltransferase activity]等功能条目显著富集。
2.4DEGs的KEGG信号通路富集分析结果
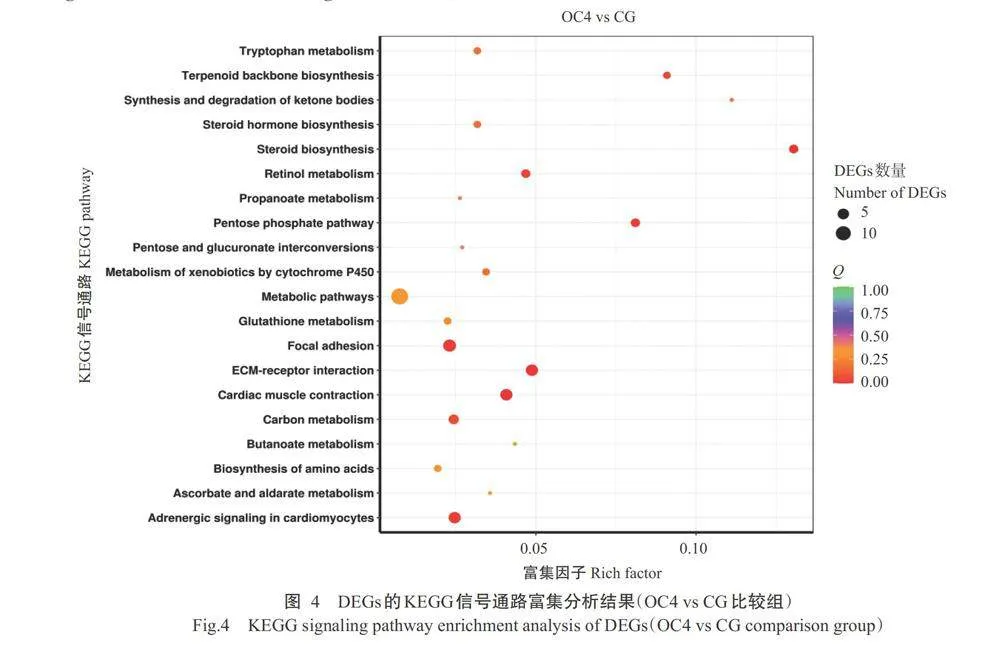
对DEGs进行KEGG信号通路富集分析。OC4vs CG比较组分析结果如图4所示,DEGs极显著(Plt;0.01,下同)富集的信号通路为ECM-受体相互作用(ECM-receptor interaction)、心肌收缩(Cardiac mu- scle contraction)、类固醇生物合成(Steroid biosynthe- sis)、磷酸戊糖途径(Pentose phosphate pathway)、黏 着(Focal adhesion)、心肌细胞中的肾上腺素能信号 传导(Adrenergic signaling in cardiomyocytes)、视黄醇代谢(Retinol metabolism)和萜类骨架生物合成 (Terpenoid backbone biosynthesis)。OC2 vs CG比较组分析结果如图5所示,DEGs极显著富集的信号通 路为代谢途径(Metabolic pathways)、剪接体(Spli- ceosome)、细胞周期(Cell cycle)、氧化磷酸化(Oxida- tive phosphorylation)、真核生物中的核糖体生物发生 (Ribosome biogenesis in eukaryotes)、DNA复制(DNA replication)、三羧酸循环[Citrate cycle(TCA cycle)]、 赖氨酸降解(Lysine degradation)、错配修复(Mis- match repair)、蛋白酶体(Proteasome)、核苷酸切除 修复(Nucleotide excision repair)、萜类骨架生物合成 (Terpenoid backbone biosynthesis)、剪接体(Spliceo- some)、N-聚糖生物合成(N-Glycan biosynthesis)、黄 体酮介导的卵母细胞成熟(Progesterone-mediated oocyte maturation)、谷胱甘肽代谢(Glutathione meta- bolism)、氧化磷酸化(Oxidative phosphorylation)和脂 肪酸降解(Fatty acid degradation)。
其中pacl基因位于蛋白酶体信号通路,lsm7、 acaa2、snrpa、phf5a、snrpc和u2afl等核糖体小亚基 蛋白基因富集于剪接体信号通路,此外ndufaf3基因也位于剪接体信号通路,mcm6、mcm2、rfc4和nop58等基因富集于DNA复制信号通路,mdm2、rpa2和pcna等基因富集于等细胞周期信号通路,shmt2、mgst1、dut、hsd17b10、sucla2和glo1等基因富集于磷酸戊糖途径信号通路,gstf7、gstll和gstf8等基因富 集于谷胱甘肽代谢信号通路,nans基因位于氧化磷 酸化信号通路。
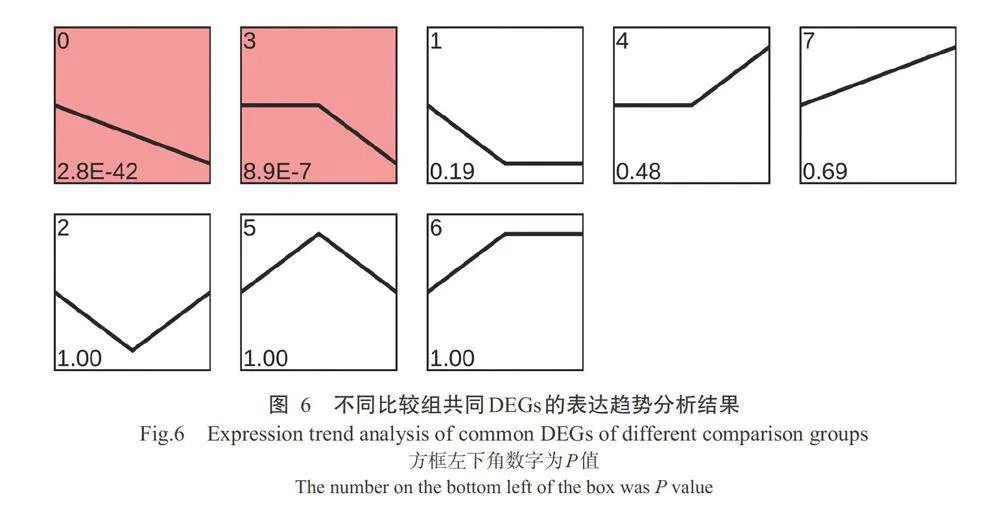
2.5DEGs表达趋势分析
对3个比较组OC4vs CG、OC2vs CG和OC4vs OC2的255个共同DEGs进行表达趋势分析,共生 成8种基因表达趋势模式(图6),其中有2种极显著 的基因表达趋势模式;Profile 0模式表示溶解氧含量 为3.99士0.18和2.05+0.15mg/L时,基因表达量持续 下降;Profile3模式表示基因表达量在溶解氧含量为 3.99±0.18mg/L时无明显变化,但在溶解氧含量为2.05±0.15mg/L时下降。其中Profile 0模式中含52个DEGs,包括位于蛋白酶体信号通路的pacl基因, 位于细胞周期信号通路的mdm2、rpa2和pcna基因,位于磷酸戊糖途径信号通路的shmt2、mgstl和dut 基因,位于氧化磷酸化信号通路的nans基因,位于 剪接体信号通路的ndufaf3基因;Profile 3模式中含 16个DEGs,包括Ism7、acaa2和snrpa等核糖体小亚基蛋白基因,mcm6、mcm2和rfc4等DNA复制过程 相关基因,gsyf7、gstll和gsyf8等谷胱甘肽代谢相关 基因。
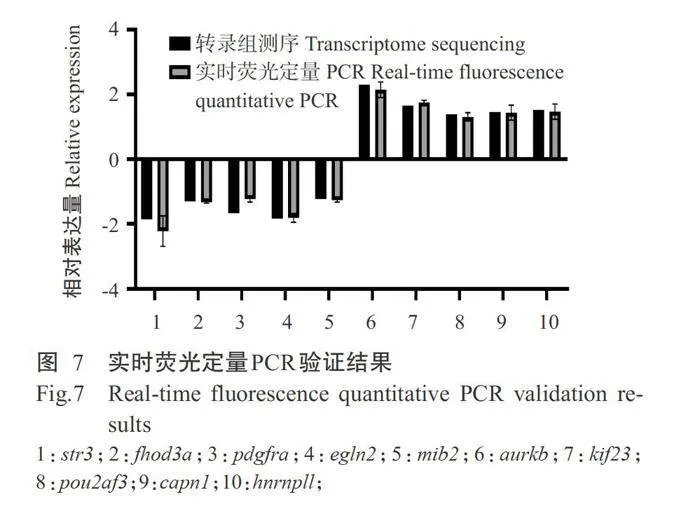
2.6实时荧光定量PCR验证结果
实时荧光定量PCR验证结果如图7所示,选取的10个DEGs表达趋势与转录组测序结果一致,表 明转录组测序结果可靠。
3讨论
3.1低氧对四指马鲅幼鱼鳃组织基因表达的影响
鳃组织是鱼类呼吸、排泄和调节渗透压等功能的重要器官,由于其直接与水体接触,极易受水环境 变化的影响。本研究对不同溶解氧含量处理的四指 马鲅鳃组织进行转录组测序并筛选DEGs,结果表明3个比较组OC4 vs CG、OC2 vs CG和OC4 vs OC2的 下调表达基因均多于上调,与低氧胁迫下团头鲂(陈 柏湘,2016)的转录组研究结果一致,说明四指马鲅 幼鱼鳃组织应对溶解氧含量下降,主要是通过抑制 基因表达进行响应。本研究对所有DEGs进行GO 功能注释分析,结果显示注释到生物学进程、细胞组 分和分子功能三大类中,主要涉及代谢过程、有机物 生物合成、细胞氧化还原平衡、细胞内、染色质、核糖 体结构成分和催化活性等功能条目,表明四指马鲅 幼鱼在不同溶解氧条件下,其鳃组织的代谢与细胞 活性均增强。KEGG信号通路富集分析结果显示,DEGs富集在蛋白酶体、DNA复制、脂肪酸降解、氧 化磷酸化、谷胱甘肽代谢、赖氨酸降解和核糖体生 物发生等信号通路。本研究对3个比较组的共同 DEGs进行表达趋势分析,表达趋势分析通过将表达 模式相似的基因进行归类,挖掘出试验变化过程中 最具代表性的基因集和相应表达趋势特性,以揭示 生物样本在变化过程中的特殊规律(郭睿等,2018)。
3.2低氧对四指马鲅幼鱼鳃组织蛋白合成相关 基因的影响
本研究中,随着溶解氧含量下降,四指马鲅幼鱼 鳃组织的pacl基因表达量下调,其位于蛋白酶体信 号通路;Rousseau和Bertolotti(2018)发现蛋白酶体 可高效降解细胞中的各种蛋白质,并调节细胞周期、 转录、信号传导和运输等多种生命活动,蛋白酶体异 常或缺失与机体损伤有关;上述结果表明四指马鲅 通过抑制pacl表达抵御由水环境溶解氧含量下降而引起机体损伤。Ism7、acaa2、snrpa、phf5a、snrpc 和u2af1等多个核糖体蛋白基因富集在剪接体通路 上;Xie等(2018)发现在rRNA加工中,核糖体蛋白起关键作用,帮助rRNA折叠形成功能性立体结构, 并使核糖体的最终空间构象保持稳定;Yamashita等 (2006)研究发现,U2AF1参与催化线粒体β氧化途 径最后一步,而线粒体β氧化途径是将脂肪酸分解 成乙酰辅酶A的有氧过程;上述结果表明四指马鲅 通过增加对脂肪的利用率及改变核糖体蛋白调节翻 译来应对溶解氧含量下降。mcm6、mcm2、rfc4和 nop58等富集于DNA复制通路;大量研究表明, MCM复合物是真核细胞中DNA复制起始和伸长所必需的复制解旋酶,在复制过程中解开模板DNA的分子机器,并围绕其构建复制体(Rzechorzek et al., 2020;Jenkyn-Bedford et al.,2021;Jones et al.,2021)。 此外,mdm2、rpa2和pcna富集于等细胞周期通路; Elkholi等(2019)研究表明,MDM2为E3泛素蛋白连接酶,介导p53/TP53的泛素化,导致其被蛋白酶体降解,MDM2通过结合其转录激活结构域来抑制p53/TP53和p73/TP73介导的细胞周期停滞和凋亡, p53基因是重要的抑癌基因,在细胞凋亡、细胞循环 和细胞增殖中发挥重要调控作用;贾若南等(2022) 对低氧胁迫和常氧条件下的斑马鱼鳃组织进行转录 组测序,发现部分DEGs与MDM2蛋白和p53基因 有关,本研究结果与之相似。本研究中随着溶解氧 含量下降,四指马鲅蛋白质合成相关基因的表达呈 下降趋势,表明四指马鲅的蛋白合成能力随着溶解 氧含量的下降而降低,可能是因为低氧影响四指马 鲅鳃组织中蛋白质翻译和mRNA合成,导致机体的 蛋白代谢水平降低,以减少氧气的消耗,从而降低了 四指马鲅幼鱼的低氧损伤。
3.3低氧对四指马鲅幼鱼鳃组织能量代谢相关 基因的影响
鱼类在低氧条件下主要通过降低代谢率和能量 需求,来维持较长时间的存活(Hochachka et al.,1996)。本研究发现,Profile 0和Profile 3基因表达趋势模式中的大量基因富集在磷酸戊糖途径通路,包括shmt2、mgst1、dut、hsd17b10、sucla2和glo1等。
GLO1催化由甲基乙二醛和谷胱甘肽形成的半硫醇 转化为S-乳酰谷胱甘肽(Chiba et al.,2012);SHMT2 催化丝氨酸裂解为甘氨酸,同时产生5,10-亚甲基四 氢叶酸,其参与线粒体翻译;hsdl7b10编码的线粒体 脱氢酶参与脂肪酸、支链氨基酸和类固醇代谢途径; sucla2编码琥珀酰辅酶A合成酶亚基,琥珀酰辅酶A 合成酶在三羧酸循环中起作用。此外,本研究还发 现了与氧化磷酸化及呼吸代谢酶相关的基因nans, 其与N-乙酰神经氨酸的生成有关(Lawrence et al., 2000)。上述基因在脂肪酸降解和三羧酸循环中发 挥重要作用,表明四指马鲅通过调节内部能量代谢 应对溶解氧含量下降;Li等(2017)研究表明,低氧影 响水生动物鳃组织中的能量代谢且其响应模式因物 种不同存在差异,本研究结果与之相符。
3.4低氧对四指马鲅幼鱼鳃组织抗氧化酶相关 基因的影响
低氧会抑制机体细胞内的能量产生,引起鱼类 体内活性氧大量累积,导致氧化应激和抗氧化系统 失调。本研究在不同溶解氧含量处理四指马鲅鳃组织中筛选出的DEGs包括gstf7、gstl1、gstf8和ndu- faf3,均为抗氧化酶相关基因。gstf7、gst11和gstf8均 属于谷胱甘肽转移酶基因家族,谷胱甘肽转移酶参 与还原型谷胱甘肽与大量外源性和内源性疏水亲电 试剂的结合,谷胱甘肽可帮助维持机体免疫系统 的正常机能并具有抗氧化性(Saetan et al.,2020); NDUFAF3是构成线粒体泛醌氧化还原酶复合物的 关键装配因子。上述基因在抗氧化损伤中发挥重要 作用,表明四指马鲅通过基因调控抵御机体受到的 氧化损伤。
4结论
四指马鲅幼鱼鳃组织蛋白质合成能力和能量代 谢活动均随着溶解氧含量下降而下降,溶解氧含量 下降显著影响细胞周期、DNA复制、脂肪酸降解和 糖代谢等相关通路。
参考文献:
陈柏湘.2016.团头鲂(Megalobrama amblycephala)低氧相关 分子标记的开发及应用[D].武汉:华中农业大学.[Chen B X. 2016. Identification and application of hypoxiaassociated molecular markers in Megalobrama amblycephala[D]. Wuhan:Huazhong Agricultural University.]
郭睿,解彦玲,熊翠玲,尹伟轩,郑燕珍,付中民,陈大福.
2018.意大利蜜蜂4、5和6日龄幼虫肠道发育过程中差异表达基因的趋势分析[J].上海交通大学学报(农业科学版),36(4):14-21.[Guo R,Xie YL,Xiong CL,Yin W X,Zheng Y Z,Fu Z M, Chen D F. 2018. Trend analysis for sifferentially expressed genes in the developmental process of 4-,5- and 6-day-0ld larval guts of Apis mellifera ligustica [J]. Journal of Shanghai Jiaotong University (Agricultural Science) ,36(4) : 14-21.] doi: 10.3969/J.ISSN.1671-9964.
2018.04.003.
贾若南,林枫,许强华.2022.低氧胁迫下斑马鱼鳃中核糖体 蛋白基因家族的表达分析[J].上海海洋大学学报,31
(2) : 318-327. [Jia R N, Lin F, Xu Q H. 2022. Differential expression analysis of the ribosomal protein gene family in zebrafish gills under hypoxia stress [J]. Journal of Shanghai Ocean University,31(2):318-327.] doi: 10.12024/jsou.
20210403367.
区又君,李加儿.2022.四指马鲅养殖与生物学[M].北京:海洋出版社:1-4.[Ou YJ,LiJE.2022.Culture and biology of four-fingered threadfin[M]. Beijing:China Ocean Press:1-4.]
潘炯华,钟麟,郑慈英,伍汉霖,刘家照.1991.广东淡水鱼类 志[M].广州:广东科技出版社:355-356.[PanJH,Zhong L, Zheng C Y, Wu H L, Liu J Z. 1991. The freshwater fishes of Guangdong Province[M. Guangzhou: Guangdong Science amp; Technology Press:355-356.]
孙典荣,陈铮.2013.南海鱼类检索(上册)[M].北京:海洋出
版社:380-382.[Sun D R,Chen Z.2013.South China Sea fish search (Volume 1) [M]. Beijing: China Ocean Press:380-382.]
吴志昊,尤锋,王英芳,文爱韵,马得友,徐永立,张培军.
2011.低氧和高氧对大菱鲆幼鱼红细胞核异常及氧化抗 氧化平衡的影响[J].上海海洋大学学报,20(6):808- 813. [Wu Z H,You F,Wang Y F,Wen A Y,Ma D Y,Xu Y
L, Zhang P J. 2011. The effects of hypoxia and hyperoxia on nucleus anomaly, SOD, CAT activities and MDA content in juvenile turbot Scophthalmus maximus [J]. Journal of Shanghai Ocean University,20(6) :808-813.]
谢木娇,区又君,李加儿,温久福,周慧,王鹏飞,刘奇奇.
2017.四指马鲅(Eleutheronema tetradactylum)消化系统 胚后发育组织学观察[J].渔业科学进展,38(2):50-58.
[Xie M J,Ou Y J, Li J E, Wen J F,Zhou H,Wang P F,Liu Q Q. 2017. Histological observation of the post-embryonic development of digestive tract of Eleutheronema tetradactylum[J]. Progress in Fishery Sciences,38(2):50-58.] doi:
10.11758/yykxjz.20151125001.
谢木娇,区又君,温久福,李加儿,王鹏飞,周慧,刘奇奇.
2016a.四指马鲅(Eleutheronema tetradactylum)受精卵和 仔鱼对不同盐度的耐受性[J].生态学杂志,35(5):1263- 1267. [Xie M J, Ou Y J,Wen J F, Li J E, Wang P F,Zhou H, Liu Q Q. 2016a. Salinity tolerance of fertilized eggs and larva of Eleutheronema tetradactylum [J]. Chinese Journal
of Ecology,35(5):1263-1267.] doi: 10.13292/j.1000-4890.201605.007.
谢木娇,区又君,温久福,李加儿,周慧,王鹏飞,刘奇奇.
2016b.四指马鲅胚胎发育观察[J].应用海洋学学报,35
(3):405-411.[Xie M J,Ou Y J,Wen J F,Li J E,Zhou H, Wang P F, Liu Q Q. 2016b. Observation on embryonic development of Eleutheronema tetradactylum [J]. Journal of Applied Oceanography,35 (3) : 405-411.] doi: 10.3969/ J.ISSN.2095-4972.2016.03.012.
庄平,王幼槐,李圣法,邓思明,李长松,倪勇.2006.长江口鱼 类[M].上海:上海科学技术出版社:194-195.[Zhuang P, Wang Y H, Li S F, Deng S M, Li C S, Ni Y. 2006. Fishes of the Yangtze Estuary[M]. Shanghai: Shanghai Scientific amp; Technical Publishers: 194-195.]
Buchheister A, Bonzek C F, Gartland J, Latour R J. 2013. Patterns and drivers of the demersal fish community of Chesapeake Bay [J」. Marine Ecology Progress Series, 481: 161- 180. doi: 10.3354/meps10253.
Chiba T, Ohwada J, Sakamoto H, Kobayashi T, Fukami T A, Irie M, Miura T, Ohara K, Koyano H. 2012. Design and evaluation of azaindole-substituted N-hydroxypyridones as
glyoxalase I inhibitors[J]. Bioorganic amp; Medicinal Chemistry Letters,22(24):7486-7489. doi:10.1016/j.bmcl.2012.10.
045. Craig J K. 2012. Aggregation on the edge: Effects of hypoxia avoidance on the spatial distribution of brown shrimp and demersal fishes in the Northern Gulf of Mexico[J]. Marine Ecology Progress Series, 445: 75-95. doi: 10.3354/meps
09437. Eby L A, Crowder L B. 2002. Hypoxia-based habitat compression in the Neuse River Estuary: Context-dependent shifts in behavioral avoidance thresholds [J]. Canadian Journal of Fisheries and Aquatic Sciences , 59 (6) : 952-965. doi :10.1139/f02-067.
Elkholi R, Abraham-Enachescu I, Trotta A P, Rubio-Patiño C, Mohammed J N, Luna-Vargas M P A, Gelles J D, Kami- netsky J R, Serasinghe M N, Zou C, Ali S, McStay G P, Pfleger C M, Chipuk J E. 2019. MDM2 integrates cellular respiration and apoptotic signaling through NDUFS1 and
the mitochondrial network[J]. Molecular Cell, 74(3):452-
465. doi: 10.1016/j.molcel.2019.02.012.
Hochachka P W,Buck L T,Doll C J,Land S C. 1996. Unifying
theory of hypoxia tolerance: Molecular/metabolic defense and rescue mechanisms for surviving oxygen lack[J]. Proceedings of the National Academy of Sciences of the United States of America,93(18):9493-9498. doi: 10.1073/
pnas.93.18.9493.
Jenkyn-Bedford M,Jones M L, Baris Y, Labib K P M, Cannone G, Yeeles J T P, Deegan T D. 2021. A conserved mechanism for regulating replisome disassembly in eukaryotes
[J]. Nature, 600(7890): 743-747. doi: 10.1038/s41586-021-04145-3.
Jones M L, Baris Y, Taylor M R G, Yeeles J T P. 2021. Structure of a human replisome shows the organisation and interactions of a DNA replication machine [J]. The EMBO Journal,40(23):e108819. doi:10.15252/embj.2021108819.
Lawrence S M, Huddleston K A, Pitts L R, Nguyen N, Lee Y C, Vann W F, Coleman T A, Betenbaugh M J. 2000. Cloning and expression of the human N-acetylneuraminic acid phosphate synthase gene with 2-keto-3-deoxy-d-glycero- dgalacto-nononic acid biosynthetic ability[J]. Journal of Biological Chemistry,275(23):17869-17877. doi: 10.1074/jbc.
M000217200.
Li H L, Lin H R, Xia J H. 2017. Differential gene expression profiles and alternative isoform regulations in gill of Nile tilapia in response to acute hypoxia [J]. Marine Biotechno-
logy,19(6):551-562. doi:10.1007/s10126-017-9774-4.
McClatchie S, Goericke R, Cosgrove R, Auad G, Vetter R.
2010. Oxygen in the Southern California Bight: Multidecadal trends and implications for demersal fisheries[J].
Geophysical Research Letters , 37 (19 ) : L19602. doi : 10.1029/2010GL044497.
Pihl L, Baden S P, Diaz R J. 1991. Effects of periodic hypoxia on distribution of demersal fish and crustaceans[J]. Marine
Biology,108:349-360. doi:10.1007/BF01313644.
Pihl L, Baden S P, Diaz R J, Schaffner L C. 1992. Hypoxiainduced structural changes in the diet of bottom-feeding fish and Crustacea[J]. Marine Biology, 112: 349-361. doi:
10.1007/BF00356279.
Rousseau A, Bertolotti A. 2018. Regulation of proteasome assembly and activity in health and disease[J]. Nature Reviews Molecular Cell Biology, 19 (11) , 697-712. doi: 10.
1038/s41580-018-0040-z.
Rzechorzek N J, Hardwick S W, Jatikusumo V A, Chirgadze D Y, Pellegrini L. 2020. CryoEM structures of human CMGATPyS-DNA and CMG-AND-1 complexes[J]. Nucleic Acids Research, 48 (12) : 6980-6995. doi : 10.1093/nar/gkaa429.
Saetan W,Tian C X, Yu J W, Lin X H,He F X, Huang Y,Shi H J, Zhang Y L, Li G L. 2020. Comparative transcriptome
analysis of gill tissue in response to hypoxia in silver sillago (Sillago sihama) [J]. Animals, 10 (4) : 628. doi: 10.3390/ani10040628.
Sun S M, Xuan F J, Fu H T, Zhu J, Ge X P, Gu Z M. 2015. Transciptomic and histological analysis of hepatopancreas, muscle and gill tissues of oriental river prawn (Macrobrachium nipponense) in response to chronic hypoxia [J].
BMC Genomics, 16 (1) : 491. doi: 10.1186/s12864-015-1701-3.
Valavanidis A, Vlahogianni T, Dassenakis M, Scoullos M.
2006. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants [J].
Ecotoxicology and Environmental Safety, 64(2) : 178-189.
doi: 10.1016/j.ecoenv.2005.03.013.
Wu R S S. 2002. Hypoxia:From molecular responses to ecosystem responses [J]. Marine Pollution Bulletin, 45 (1-12) :
35-45. doi:10.1016/S0025-326X(02)00061-9.
Xie X, Guo P, Yu H, Wang Y, Chen G. 2018. Ribosomal pro-
teins: Insight into molecular roles and functions in hepatocellular carcinoma [J]. Oncogene,37 (3) : 277-285. doi: 10.1038/onc.2017.343.
Yamashita H, Itsuki A, Kimoto M, Hiemori M, Tsuji H. 2006. Acetate generation in rat liver mitochondria; acetyl-CoA hydrolase activity is demonstrated by 3-ketoacyl-CoA thiolase [J]. Biochimica et Biophysica Acta (BBA)- Molecular and Cell Biology of Lipids,1761(1):17-23. doi:10.1016/j.
bbalip.2006.01.001.
Zák J, Anil A N, Dyková I. 2023. Dissolved oxygen saturation is crucial for gas bladder inflation in turquoise killifish (Nothobranchius furzeri) [J]. Environmental Biology of
Fishes,106(4):673-683. doi:10.1007/s10641-023-01405-1.
(责任编辑刘可丹)
