四川宁南钟家梁子遗址人骨的稳定同位素
2024-01-01朱语桐张国郑万泉张燕

摘要:本文通过对四川宁南县钟家梁子遗址人骨样品的C、N 稳定同位素分析,探索该遗址及金沙江中下游地区先民的食物结构和生业经济。结果显示,钟家梁子遗址先民的δ13C 值范围为-18.4‰~-10.0‰,δ15N 值范围为4.76‰~12.63‰,表明其食物结构中C3、C4 类食物均占有一定比例,且摄入了较多的动物性蛋白;不同性别、年龄及葬俗的群体间存在食谱差异。结合当地的自然环境及相关考古成果,推测钟家梁子遗址先民从事稻/ 粟混作农业,并通过采集野生植物进行补充;对动物资源的利用具有多样性,渔猎及家畜饲养均占有重要地位。通过对金沙江中下游地区相关研究的进一步梳理,发现该地区新石器时代晚期至青铜时代的作物结构一直以稻/ 粟混作为主,青铜时代开始种植小麦、大麦、大豆等作为补充,对植物资源的利用越发灵活和多样化。此外,该地区不同遗址的经济结构存在较大差别,可能受到了自然环境、地形阻隔、人群交流及当地文化传统等因素的影响。
关键词:金沙江;人骨;稳定同位素;饮食;生业经济
1 引言
金沙江是长江的上游,穿行于川、藏、滇三省区之间[1]。金沙江流域属于川滇青藏民族走廊的一部分,自古以来便是民族迁徙及文化交流的重要通道,同时又是星罗棋布的小区域文化的孕育之地[2]。该流域一直是民族文化最为丰富多彩的地区之一,这与其历史进程密切相关,而生业经济的发展为当地的文明化进程奠定了基础。了解该地区早期生业经济的特点及发展过程,对理解这一地区文化的面貌及形成机制有重要意义。目前金沙江中下游地区开展的田野发掘及研究工作较为集中,为探讨本区域生业经济提供了丰富的材料。当地山高谷深,人群的生业经济呈现出明显的立体分布特征,因而即便相距不远,先民的经济结构也会因环境差异而呈现出复杂的面貌。当地的生业面貌、先民对环境的适应策略、农作物的传播等问题,是学者们关注的重点。
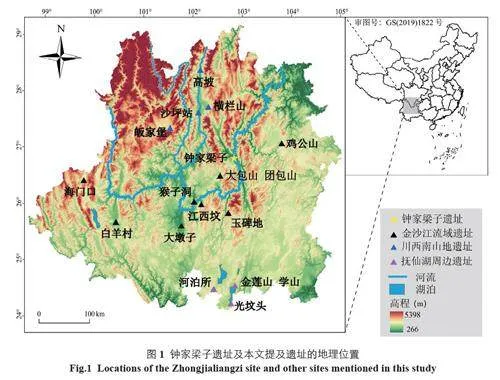
图1 展示了金沙江中下游地区新石器时代晚期至青铜时代进行过生业经济研究的主要遗址点分布。目前生业经济的研究大多来自植物考古学,许多遗址如宾川白羊村[3]、元谋大墩子[4]、东川玉碑地[5] 等,均发现稻、粟、黍等植物遗存共存的现象,为该地区的农业发展研究提供了直接的考古学证据。动物考古学研究较少,大多仅在发掘报告中列出了出土动物骨骼的种属,显示该地区先民饲养猪、狗等家畜,同时渔猎活动也占有较大比重[6];C、N 稳定同位素分析则限于会理猴子洞[7]、武定江西坟[8] 等少数几个遗址,均表现出C3、C4类食物混合的现象。由此可见,该地区农业结构应以稻与粟混作为主,但其生业经济的特点,不同群体或个体对稻、粟依赖程度的差异,随时间有无变化等问题都未得到很好地揭示。此外,目前的资料多集中在中游地区,下游地区先民的食谱及生业经济尚不清晰。
人骨的C、N 稳定同位素分析,为了解个体所食用的植物类型、动物蛋白质来源及其在饮食中的相对重要性提供了可能,是揭示先民食物结构和生活方式、探索农业起源与发展的主要方法之一[9,10]。钟家梁子遗址位于金沙江下游左岸的台地上,是近年来在金沙江干流两岸发掘面积较大的一处遗址,可作为研究金沙江中下游地区早期文化的重要切入点。本文以钟家梁子遗址出土人骨为研究对象,对其进行C、N 稳定同位素分析,试图揭示该遗址先民的食物结构、生业经济等信息,并结合周边遗址的研究成果,对金沙江中下游地区新石器时代晚期至青铜时代的经济结构进行探讨,以期对该地区先民的文化面貌、文明化进程等问题的深入阐释起到促进作用。
2 材料和方法
2.1 遗址概况
钟家梁子遗址(26°55′43.2″N, 102°52′49.58″E)位于四川省凉山彝族自治州宁南县华弹镇金江村,地处金沙江北岸的二级阶地上,面积约4 万平方米,分布在高差达70 m 的斜坡上(图1)。遗址共发现石棺墓27 座、土坑墓242 座、瓮棺18 座,出土陶器、石器、骨器、蚌器等各类随葬品463 件(组)[11](图2)。14C 测年约为2200 BC~1300 BC,属于新石器时代晚期至夏商时期 [12]。西南地区土壤酸性较强,人骨普遍保存不佳,但该遗址出土的大量保存状况较好的人骨遗存,为本研究提供了丰富的材料。
2.2 样品的选择
本文样品选自钟家梁子遗址墓葬出土的168 例人骨,部分个体同时选取了肋骨和肢骨进行同位素对比分析,以揭示个体生前不同阶段食物结构的变化情况(样品共计197 件,其出土单位、取样部位及考古学背景信息详见本文网络版附属材料)。
2.3 样品预处理
骨胶原的提取主要依据Richards 和Hedge 的方法[13]。首先用打磨器清除骨样表面的污染物,取大约500~1000 mg 的骨样,将其浸泡在0.5 mol/L 的盐酸溶液中,放置于5℃环境下,每2~3 天更换盐酸溶液,直至骨样松软且溶液无明显气泡,用去离子水洗至中性,放入带有0.001 mol/L 盐酸溶液的密封试管中,于70℃明胶化48 h,趁热过滤,将骨胶原溶液冷冻干燥后得到骨胶原,然后称重并计算骨胶原产率。
2.4 样品测试
骨胶原的C、N 元素含量及稳定同位素比值测定在中国农业科学院农业环境与可持续发展研究所测试中心进行,所用仪器为稳定同位素质谱分析仪(Isoprime 100 coupled withElementar Vario)。测试C、N 元素含量所用标准物质为磺胺(Sulfanilamide)。C、N 稳定同位素比值分别以IAEA-600、IAEA-CH-6 标定碳钢瓶气(以VPDB 为基准)和IAEA-600、IAEA-N-2 标定氮钢瓶气(以AIR 为基准),每测试10 个样品插入1 个实验室自制胶原蛋白标样(δ13C 值为-14.7‰±0.1‰,δ15N 值为7.0‰±0.1‰)。分析精度均低于±0.2‰,测试结果以δ13C(相对于VPDB),δ15N(相对于AIR)表示(详见本文网络版附属材料)。
3 结果与讨论
3.1 污染样品的辨别
在长期埋藏过程中,骨骼会受到温湿度、微生物等因素的影响,导致其结构和化学组成发生改变。样品没有受到污染是进行骨骼稳定同位素分析的前提。
由数据计算可知, 钟家梁子遗址所有骨骼样品(n=197) 的骨胶原提取率在0.01%~46.43% 之间,均值约2.85%±3.9%,而现代骨胶原提取率约为20.0%,表明该遗址骨骼在埋藏过程中存在一定程度的降解[14]。
判断骨胶原是否被污染最重要的指标是骨胶原的C、N 含量及C/N 摩尔比值。若骨胶原的C、N 含量分别位于15.3%~47.0% 和5.5%~17.3%,C/N 摩尔比值介于2.9~3.6 之间,可视为基本未被污染[15,16]。钟家梁子遗址骨胶原样品(n=197) 的C 含量在1.74%~43.53%之间,均值约25.55%±13.3%(n=197);N 含量在0.17%~15.27% 之间,均值约8.80%±5.0%;C/N 摩尔比值在1.5~11.8 之间,表明部分样品受到了污染。经筛选,共有139 例样品的C/N 摩尔比值落于2.9~3.6 范围内,可用作下文的稳定同位素分析。其余58 例样品的C/N摩尔比值落于此有效值范围外(详见本文网络版附属材料),下文研究时予以剔除。
3.2 先民食谱分析
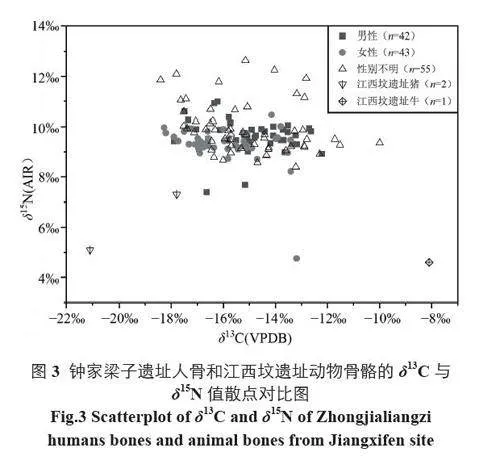
钟家梁子遗址未能获取到动物骨骼样品来进行稳定同位素分析。为评估该遗址先民对动物蛋白的消耗情况,本文将相近的云南江西坟遗址出土动物骨骼的稳定同位素数据纳入比较[8],以钟家梁子遗址未污染人类骨骼样品及江西坟遗址猪和牛的δ13C 和δ15N 值作散点图,探查数据的分布情况(图3)。
根据光合作用途径的不同,可将植物分为C3、C4 和CAM (crassulacean acid metabolism)途径,其中前两类与人类生活更为密切[17]。不同途径的植物,其δ13C 值也各不相同。C3 类植物主要为稻、麦、豆类等,δ13C 均值为-26.5‰;C4 类植物主要包括粟、黍、玉米等,δ13C 均值为-12.5‰[18]。当植物被动物所食,动物骨骼中骨胶原的δ13C 值将发生约5.0‰ 的富集[19]。因此,分别以C3 和C4 类植物为食的动物和人,其骨胶原中的δ13C 值大致为-21.5‰和-7.5‰。由此,可通过人和动物骨胶原的δ13C 值来了解其所食用的食物类型。
由图3 可见,钟家梁子遗址先民(n=139) 的δ13C 值范围为-18.4‰~-10.0‰,均值为-15.3‰±1.6‰(具体数据详见网络版附属材料)。总体而言,先民的δ13C 值较为分散,意味着其内部可能存在食物结构的差异。绝大多数样品的δ13C 值介于-18.0‰~-12.0‰,表明大部分先民的食物结构中C3 和C4 类食物均占有一定比例;也有少量个体的δ13C 值小于-18.0‰ 或大于-12.0‰,表明其主要食用C3 类食物或C4 类食物。但由于该遗址δ13C 值分布范围整体较广,这些墓葬的数值相比于其他墓葬并未表现出明显的特殊性。考虑到与钟家梁子遗址年代相近且同处金沙江中下游的元谋大墩子[4]、武定江西坟[8]、威宁鸡公山[20]等遗址均出土了稻、粟、黍等炭化谷物遗存,推测钟家梁子遗址先民的C3 类食物与水稻及C3 类野生植物有关,C4 类食物应主要源自粟、黍或间接来自以C4 植物为食的动物。
N 元素在沿营养级上升时存在着同位素的富集现象,每上升一级,δ15N 约增加3.0‰~5.0‰[21]。故根据骨骼中的δ15N 值,可辨别生物所处营养级的位置。一般认为,植食类动物的δ15N 值约为3.0‰~7.0‰,杂食类动物约为7.0‰~9.0‰,肉食类动物则大于9.0‰[22]。由图3 可知,钟家梁子遗址先民(n=139) 的δ15N 值范围为4.76‰~12.63‰,均值为9.6‰±0.9‰(具体数据详见网络版附属材料)。与δ13C 值相比,先民的δ15N 值差异相对较小。先民与江西坟遗址牛的δ15N 均值相差5.0‰,与猪的δ15N 均值相差3.4‰,表明多数先民摄入了大量的动物蛋白。但由于江西坟遗址动物样品量较少,这一结论有待后续研究的检验。值得注意的是, 钟家梁子遗址M18 的δ15N值(4.76‰) 低于7.0‰,表明该个体以植物性食物为主,但具体原因还需结合考古资料进一步分析;M182、M252、M257 三座墓葬的δ15N 值大于12.0‰,说明他们的营养级较高。研究表明,由于婴儿直接摄取母亲的母乳,故其营养级相较于母亲得以提升,δ15N值较其母亲富集约2‰~3‰[23,24]。由于上述三座墓葬的墓主均为婴幼儿,故其较高的δ15N 值可能与母乳喂养有关。
3.3 先民食谱的内部差异
3.3.1 不同性别年龄先民的食谱差异
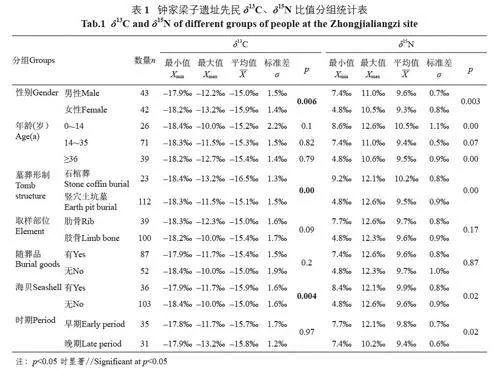
为了进一步比较不同性别、年龄人群食物结构的差异,本文分别统计了男性、女性、未成年人、青壮年及中老年人群的δ13C、δ15N 值(表1)。由于部分数据不满足正态分布,故本文采用Kruskal-Wallis 方差分析对不同群体间的差异进行比较。从性别上看,钟家梁子遗址男女性的δ13C 和δ15N 值均存在显著差异(δ13C: p=0.006lt;0.05; δ15N: p=0.003lt;0.05);男性相比于女性摄入更多的C4 类食物和动物性蛋白。
年龄方面,本文将0~14 岁划分为未成年组,其余再以35 岁为界,分别划分为青壮年和中老年组[25]。如表1 所示,三个年龄组的δ13C 值没有显著差异(p=0.97gt;0.05),表明不同年龄人群食用的植物类型并无明显不同。然而,未成年组δ15N 值整体上显著高于青壮年组(p=0.00lt;0.05) 和中老年组(p=0.00lt;0.05),而青壮年组与中老年组间不存在显著差异(p=0.07gt;0.05)。由此可知,钟家梁子遗址未成年人较成年人摄入了更多的动物性蛋白。
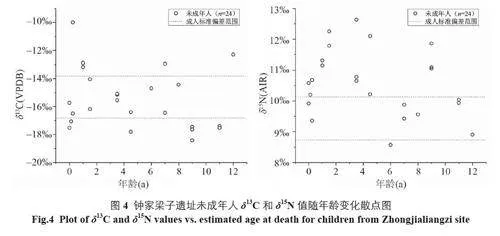
若将未成年人的δ13C 和δ15N 值按年龄变化进行排列,可进一步推测其断奶过程。研究表明,纯母乳喂养儿童的δ13C 值较母亲升高约1‰[26]。由图4 可见,部分0 岁左右死亡的婴儿可能未接受良好的母乳喂养,其数值与成人无异,暂时不予考虑;1.5 岁儿童的δ13C 值已位于成人范围内,说明一些儿童至少在1.5 岁前便有了辅食摄入。此外,5 岁之前儿童的δ15N 值明显高于成人,5 岁之后回落到成人范围,表明该遗址儿童在5 岁之前已完成断奶,考虑到骨骼更新速率的影响[27,28],实际断奶时间应该更早。部分个体在9 岁仍有较高的δ15N 值,说明其可能仍摄入了一定量的母乳或食用了较多的肉食。但由于样品代表性及骨骼更新滞后性等问题,上述结论有待后续研究验证。相比于未成年人,青壮年和中老年的δ15N 值相近,表明钟家梁子遗址先民成年后食物结构较为稳定。
3.3.2 个体不同生长时段的食谱差异
先民食物结构的稳定性也可从不同部位骨骼的稳定同位素比值中得到证实。人体不同部位骨骼的代谢速率存在一定差异,如长骨骨胶原中的化学成分一般需要10 年以上的时间才能完全更新[27],而肋骨只需2~5 年[28]。因此,肢骨和肋骨的稳定同位素值可分别代表个体生前10 年和2~5 年的饮食状况。由表1 可见,取样于肋骨和肢骨的个体,其δ13C值(p=0.09gt;0.05) 和δ15N 值(p=0.17gt;0.05) 并无显著差异。此外,绝大部分取样于同一个体的肢骨和肋骨,其δ13C、δ15N 值也无明显变化,说明该遗址先民在生前10~2 年这段时间整体的食物结构较为稳定。然而,钟家梁子遗址M254 肢骨的δ13C 值明显低于肋骨,δ15N值明显高于肋骨;M256 肢骨的δ13C 和δ15N 均明显低于肋骨,表明两个个体的食物来源发生了较大改变。这可能与个体生前饮食习惯的变化有关,两座墓葬中出土的不属于本地的海贝表明墓主也存在迁徙活动的可能,但还有待于锶同位素等研究的验证。
3.3.3 先民食谱的时间变化
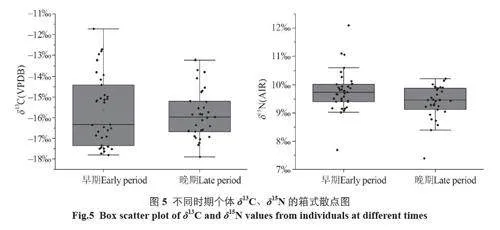
根据测年结果及墓葬出土的随葬品信息,钟家梁子遗址可大体分为早、晚两期[12]。由表1 可知,早期与晚期先民的δ13C 值无显著差异(p=0.97gt;0.05),但早期δ13C 值的分布范围明显大于晚期(图5),表明早期先民的食物来源更加多样化,晚期则更为固定。δ15N 值方面,晚期较早期有所降低(p=0.02lt;0.05),表明晚期先民的食物中动物性蛋白减少。可见先民的食物结构随时间略有变化,可能与农业的进一步发展有关。
3.3.4 不同葬俗先民的食谱差异
埋葬习俗可以在一定程度上反映当时的社会经济状况、社会习俗、个体的身份认同等,如随葬品的数量、质量及种类可能与死者的社会地位和社会身份有关[29]。而食物除营养价值外也常被赋予一定的社会内涵,以传达食用者的身份信息。本文拟对不同墓葬形制、随葬品群体的δ13C和δ15N值分别进行统计和对比,考察不同葬俗群体在食物结构上的差异。
由于瓮棺葬数量极少,且墓主均为婴儿,本文主要比较石棺葬及长方形竖穴土坑墓两个群体的食物结构。由表1 可知,长方形竖穴土坑墓墓主的δ13C 值(p=0lt;0.05) 和δ15N值(p=0lt;0.05) 均与石棺葬群体有显著差异,石棺葬墓主饮食结构中的C3 类食物比例较高,且食用较多动物性蛋白。石棺葬群体数量较少,且大多具有较大的墓葬面积及更多的随葬品,再加上其食物中含有较多的动物性蛋白,由此推测,石棺葬群体可能拥有较多的财富或较高的社会地位,也可能具有某种特殊的社会身份。
有随葬品和无随葬品两个群体的δ13C、δ15N 比值如表1 所示,两个群体的δ13C 值(p=0.2gt;0.05)、δ15N 值(p=0.87gt;0.05) 均无统计学上的显著差异,表明其食物结构并无较大区别。值得注意的是,钟家梁子遗址中部分墓葬出土了数量不等的海贝,而四川西南山地并不出产海贝,故推测海贝的出现可能与沿海地区的文化交流有关。由表1 可知,出土海贝与未出土海贝两群体的δ13C 值(p=0.004lt;0.05) 和δ15N 值(p=0.02lt;0.05) 均存在显著性差异,推测有海贝随葬的群体摄入更多C3 类植物和动物性蛋白。
3.4 先民的生业经济
自然环境对人们的生活方式有至关重要的影响。钟家梁子遗址所处的宁南县位于金沙江下游地区,当地光热充足、降水不少但蒸发量较大[30]。宁南县的年均温为19.3℃,年降水量948 mm,年日照时间2257 小时,当地地形起伏较大,立体气候明显[31]。根据钟家梁子遗址的先民骨骼δ13C 值可知,该遗址先民的食物结构为C3、C4 类食物兼有,该区域其他年代相近的遗址也发现了大量稻、粟、黍等炭化植物遗存,故推测钟家梁子先民可能从事稻/ 粟混作农业。这与当地的自然环境和地形条件密切相关。钟家梁子遗址位于高差达70 米的山坡上,坡下河流可以提供充足的水源,河谷地区适合种植水稻,而距河流较远的山坡光热充足且较为干燥,适合粟、黍等旱田作物的种植。此外,白羊村[3]、海门口[32]、玉碑地[5] 等临近遗址出土了甜瓜、桃、葡萄及一些块茎类植物。由此推测,除了农业生产,采集可能也是钟家梁子遗址先民获取植物性食物资源的方式之一。
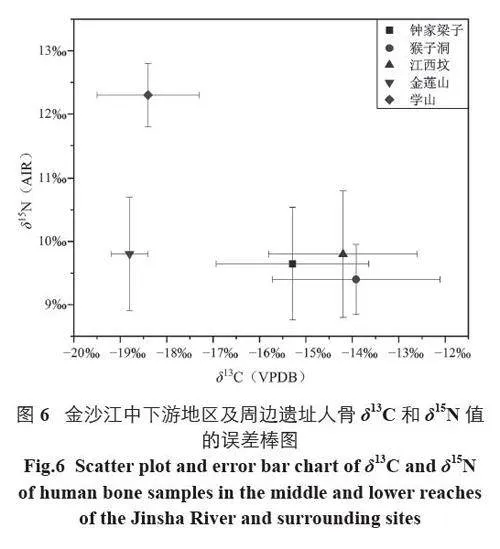
从δ15N 值可以看出,钟家梁子遗址先民摄入了较多的动物蛋白。虽然遗址内动物骨骼的出土情况暂不清楚,但可通过出土的生产工具和附近遗址的相关信息来探讨先民获取肉食资源的方式。通过与会理猴子洞、武定江西坟等遗址的稳定同位素值比较可知,该地区遗址的δ15N 均值都处于9.0‰~10.0‰,表明金沙江中下游地区先民的动物蛋白来源较为稳定,不同遗址的动物资源利用情况可能具有相似性(图6)。元谋大墩子[6]、永仁菜园子和磨盘地遗址[33]出土了猪、狗等家养动物和大量野生动物骨骼,表明新石器时代晚期饲养家畜是金沙江中下游地区先民获取肉食资源的重要方式,故钟家梁子遗址先民也极有可能饲养猪、狗等家畜。从周边遗址出土的大量野生动物骨骼推测,渔猎可能是钟家梁子遗址先民重要的生业部门,这从遗址出土的螺壳及石镞等狩猎工具也能得到证实。
3.5 金沙江中下游地区新石器时代晚期至青铜时代先民的生业经济
钟家梁子遗址出土人骨的稳定同位素数据分析,为重建金沙江中下游地区的经济状况提供了重要参考。本文梳理该区域新石器时代晚期至青铜时代先民生业经济的相关研究,结合钟家梁子遗址人骨的稳定同位素数据,深入探索该地区先民的生业经济。
目前,这一地区最早的农作物遗存出土于白羊村遗址,该遗址出土的稻、粟、黍等农作物遗存占所有可识别遗存的80% 以上,表明距今4500 年左右,稻/ 粟混作农业已在当地先民的生业经济中发挥重要作用[3]。此后各遗址也大多呈现出稻/ 粟混作的模式,只是大墩子[4]、猴子洞[7,34]、江西坟[8] 等遗址粟作农业比重较高;鸡公山[35]、玉碑地[5] 等遗址则是以稻为主,粟作农业占比较低。麦作农业在该地区最早出现于距今约3400 年的海门口二期,并逐渐成为该遗址最重要的农作物[32];春秋战国时期的大包山、大团山遗址[36]也出土了小麦和大麦遗存,但由于样品量有限,无法讨论其在农业中所占比重。此外,玉碑地遗址出土了少量小麦、大豆等遗存,可能作为一种选择性粮食作物[5]。
由此可知,各类农作物的比重存在遗址间差异。从浮选和稳定同位素分析结果来看,猴子洞、江西坟和大墩子遗址的粟作比重较高,钟家梁子、白羊村等遗址次之,鸡公山等遗址最低。遗址间经济结构的差异可能是由金沙江沿岸地貌特征多样性导致的水热条件差异造成的,钟家梁子、猴子洞等遗址可能受到干热河谷气候的影响较为干燥,适合粟、黍等作物的生长;而鸡公山遗址海拔偏高,相对湿润,适合水稻的种植。遗址间经济结构的差异体现了当地先民对局部小环境灵活的适应策略。值得注意的是,同样海拔较高的海门口遗址麦作农业发达,但鸡公山遗址并未发现麦作遗存,说明除水热条件外,地形阻隔、先民的文化传统等因素也会对经济结构造成影响。
虽然该地区作物结构存在遗址间差异,但也有一些共同点。与安宁河流域的横栏山[37]、高坡[38]、沙坪站[39] 等遗址呈现出的以稻为主、粟黍为辅的结构,以及盐源盆地皈家堡遗址[40] 粟作为主的结构不同,金沙江中下游地区的大部分遗址均表现出稻/ 粟混作的现象,且稻和粟在大部分遗址中均占有重要地位。此外,在以澄江学山[41]、晋宁河泊所[42]、江川光坟头[43] 等为代表的滇文化遗址中,已形成稻、粟、麦混作的农业格局。相比之下,金沙江中下游地区的麦作农业不算发达,除海门口遗址小麦数量较多外,多数遗址并未发现小麦遗存,至春秋战国时期的玉碑地、大包山、团包山等遗址,出土的小麦数量也较少。通过稳定同位素数值的比较也能看出,相比于金莲山[44] 遗址和学山[45] 遗址,金沙江中下游地区的钟家梁子、会理猴子洞、武定江西坟的δ13C 值偏高(图6),说明金沙江中下游地区的先民食用了较多的C4 类食物,这可能与金沙江沿岸较为干燥的气候有关;而金莲山等遗址所在的抚仙湖周围气候湿润,水资源充足,适合水稻、小麦等C3 类植物的种植。此外,该地区复杂的地形也可能对麦类作物的传播起到了阻碍作用 [8]。但由于样品量较少,这一地区麦作农业的发展状况仍不清晰,有待后续研究数据的积累。
综上,金沙江中下游地区的农作物比重存在遗址间差异,但整体以稻/ 粟混作为主;至青铜时代,该区域逐渐出现小麦、大麦、大豆等遗存,当地先民对植物资源的利用更加灵活和多样化。该地区自古以来便是各民族文化交流和人群迁徙的重要通道,其多样化的经济结构正是文化交流和复杂多样的地貌与生态条件共同作用的结果[46],同时也为当地多元文化的进一步发展奠定了经济基础。
4 结论
本文通过对钟家梁子遗址出土人骨的C、N 稳定同位素分析,揭示了该遗址先民的食物结构及生业经济模式。此外,在既有相关研究的基础上,进一步探讨了金沙江中下游地区新石器时代晚期至青铜时代先民的生业经济。研究结果如下:
1)钟家梁子遗址先民的食物结构表现为C3、C4 类食物兼有,且食谱中包含大量的动物性蛋白,不同性别、葬俗人群的食物结构存在差异。但鉴于从该遗址考古资料中未见明显的等级差异,故推测先民的食谱可能更多地受到人群组成、生活习惯等因素的影响,也可能与其在社会中扮演的角色有关[47]。
2)稻/ 粟混作农业在该遗址先民的生业经济中发挥重要作用,可能采集野生植物作为补充。先民对动物资源的利用具有多样性,家畜饲养及渔猎活动都是重要的生业方式。
3)从周边遗址的相关研究来看,金沙江中下游地区自新石器时代晚期至青铜时代,其农业结构一直以稻/ 粟混作为主。至青铜时代,先民开始种植少量小麦、大麦、大豆等作物作为补充,对植物资源的利用越发灵活和多样化。不同遗址的作物比重不尽相同,体现了金沙江沿岸地形多样性对自然环境的影响,以及当地先民对局部小环境灵活的适应策略。此外,当地的文化传统及人群交流、地形阻隔等可能也是影响经济结构的重要因素。
致谢:两位匿名审稿专家与编辑老师审稿时提出的宝贵意见对于本文的完善起到了重要作用,在此谨致谢意。感谢四川省文物考古研究院尹芊雪子、赖雨青、张又丹、张灵、蒋蕊等在取样以及论文写作过程中的帮助。
附记:本文网络版附有如下相关材料:附表1— 钟家梁子人骨样品的相关信息与测试结果,附表2— 人骨样品考古学背景信息,敬请查阅。
参考文献
[1] 周志清.论金沙江中游新石器文化圈[J].中华文化论坛,2023, 1: 131-141+190
[2] 刘弘.对金沙江考古的几点思考[J].中华文化论坛,2002, 4: 52-57
[3] Martelloa RD, Min R, Stevens C, et al. Early agriculture at the crossroads of China and Southeast Asia: Archaeobotanical evidence and radiocarbon dates from Baiyangcun, Yunnan[J]. Journal of Archaeological science: Reports, 2018, 20: 711-721
[4] 金和天,刘旭,闵锐,等.云南元谋大墩子遗址浮选结果及分析[J].江汉考古,2014, 3: 109-114
[5] 杨薇,蒋志龙,陈雪香.云南东川玉碑地遗址(2013 年度)植物遗存浮选结果及初步分析[J].中国农史,2020, 39(1): 3-11
[6] 云南省博物馆.元谋大墩子新石器时代遗址[J].考古学报,1977, 1: 43-72+169-178+73+179-181
[7] 王奕舒.四川会理县猴子洞遗址出土人骨的稳定同位素研究[D].硕士研究生论文,西安:西北大学,2020, 12-49
[8] Lu MX, Li XR, Wei WY, et al. Environmental influences on human subsistence strategies in Southwest China during the Bronze Age: A case study at the Jiangxifen site in Yunnan[J]. Frontiers in Earth Science, 2021, 9
[9] Yadira Chinique de Armas, Ulises M. González Herrera, et al. Multiproxy paleodietary reconstruction using stable isotopes and starch analysis: The case of the archaeological site of Playa del Mango, Granma, Cuba[J]. Journal of Archaeological Science:Reports, 2022, 46: 103671
[10] Richards MP. A brief review of the archaeological evidence for Palaeolithic and Neolithic subsistence[J]. European journal of clinical nutrition, 2002, 56(12): 1270-1278
[11] 郑万泉.四川凉山州钟家梁子遗址新发现两百余座商周时期墓葬[N].中国文物报,2021-12-3(008)
[12] 四川省文物考古研究院,凉山彝族自治州博物馆,宁南县文物管理所.四川宁南县钟家梁子遗址先秦时期墓葬发掘简报[J].四川文物,2023, 2: 4-28+2
[13] Richards MP, Hedges R. Stable isotope evidence for similarities in the types of marine foods used by Late Mesolithic humans at sites along the Atlantic coast of Europe[J]. Journal of Archaeological Science, 1999, 26(6): 717-722
[14] Ambrose SH, Butler BM, Hanson DB, et al. Stable isotopic analysis human diet in the Marianas Archipelago, Western Pacific[J].America Journal Physical Anthropology, 1997, 104(3): 343-361
[15] Ambrose SH. Preparation and characterization of bone and tooth collagen for isotopic analysis[J]. Journal of Archaeological Science, 1990, 17(4): 431-451
[16] DeNiro MJ. Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction [J]. Nature, 1985, 317: 806-809
[17] 张雪莲.碳十三和氮十五分析与古代人类食物结构研究及其新进展[J].考古,2006, 7: 50-56
[18] Hedges R. On bone collagen—apatite-carbonate isotopic relationships[J]. International Journal of Osteoarchaeology, 2003, 13: 66-79
[19] DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals[J]. Geochimica et Cosmochimica Acta,1978, 42(5): 495-506
[20] 孙华.四川盆地史前谷物种类的演变:主要来自考古学文化交互作用方面的信息[J].中华文化论坛,2009, S2: 147-154
[21] Hedges R, Reynard LM. Nitrogen isotopes and the trophic level of humans in archaeology[J]. Journal of Archaeological Science,2007, 34(8): 1240-1251
[22] 董豫,胡耀武,张全超,等.辽宁北票喇嘛洞遗址出土人骨稳定同位素分析[J].人类学学报,2007, 1: 77-84
[23] Fogel ML, Tuross N, Owsley DW. Nitrogen isotope tracers of human lactation in modern and archaeological populations[J].Carnegie Institution of Washington Yearbook 88, 1989, 111-117
[24] Fuller BT, Fuller JL, Harris DA, et al. Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios[J]. American Journal of Physical Anthropology, 2006, 129(2): 279-293
[25] 朱泓.体质人类学[M].北京:高等教育出版社,2004, 122
[26] Fuller BT, Harris DA, David A, et al. Isotopic investigation of pregnancy and breastfeeding in modern and archaeological populations /[J]. University of Oxford[2024-09-23]. doi:10.5137/1019-5149.JTN.4095-11.1.
[27] Hedges R, Clement JG, Thomas CDL, et al. Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements[J]. American Journal of Physical Anthropology, 2007, 133(2): 808-816
[28] Cox G, Sealy CJ. Investigating identity and life histories: Isotopic analysis and historical documentation of slave skeletons found on the Cape Town foreshore, South Africa[J]. International Journal of Historical Archaeology, 1997, 1(3): 207-224
[29] 刘莉.中国新石器时代:迈向早期国家之路[M].译者:陈星灿,乔玉,马萧林,等.北京:文物出版社,2007
[30] 赵琳,郎南军,郑科,等.云南干热河谷生态环境特性研究[J].林业调查规划,2006, 3: 114-117
[31] 罗二虎.文化与生态、社会、族群:川滇青藏民族走廊石棺葬研究[M].北京:科学出版社,2012, 335
[32] 薛轶宁.云南剑川海门口遗址植物遗存初步研究[D].硕士研究生论文,北京:北京大学,2010
[33] 云南省文物考古研究所,中国社会科学院考古研究所云南工作队,成都市文物考古研究所.云南永仁菜园子、磨盘地遗址2001 年发掘报告[J].考古学报,2003, 2: 263-296+325-328
[34] 王冰言.藏彝走廊早期旱作农业的南传:基于会理猴子洞及大坪子遗址植物遗存分析[D].硕士研究生论文,济南:山东大学,2021
[35] 张兴香,闫雪芹,张合荣,等.贵州威宁中水遗址群人类迁徙与饮食特征初探:基于牙釉质多种同位素分析[J].四川文物,2022, 5: 95-105
[36] 闫雪,姜铭,左志强,等.2015 年会理、会东县试掘遗址出土植物遗存分析报告[A].见:成都文物考古研究院.成都考古发现(2014)[C].北京:科学出版社,2016, 135-146
[37] 姜铭,闫雪,刘祥宇,等.西昌市横栏山遗址2014 年浮选结果及初步研究[A].见:成都文物考古研究院.成都考古发现(2014)[C].北京:科学出版社,2016, 115-134
[38] 姜铭,耿平,刘灵鹤,等.冕宁县高坡遗址2011 年度浮选结果鉴定简报及初步分析[A].见:成都文物考古研究院.成都考古发现(2011)[C].北京:科学出版社,2013, 331-337
[39] 闫雪,姜铭,左志强,等.2014 年西昌市沙坪站遗址出土植物遗存分析报告[A].见:成都文物考古研究院.成都考古发现(2014)[C].北京:科学出版社,2016, 155-162
[40] 闫雪,姜铭,刘祥宇,等.2015 年盐源县皈家堡遗址、道座庙遗址出土植物遗存分析报告[A].见:成都文物考古研究院.成都考古发现(2014)[C].北京:科学出版社,2016, 147-154
[41] 王祁,蒋志龙,杨薇,等.云南澄江学山遗址植物遗存浮选结果及初步研究[J].中国农史,2019, 38(2): 3-11
[42] 杨薇.云南河泊所和玉碑地遗址植物遗存分析[D].硕士研究生论文,济南:山东大学,2016, 31-41
[43] 李小瑞,刘旭.云南江川光坟头遗址植物遗存浮选结果及分析[J].农业考古,2016, 3: 20-27
[44] 张全超.云南澄江县金莲山墓地出土人骨稳定同位素的初步分析[J].考古,2011, 1: 30-33
[45] Wang TT, Wei D, Jiang ZL, et al. Microfossil analysis of dental calculus and isotopic measurements reveal the complexity of human-plant dietary relationships in Late Bronze Age Yunnan[J]. Archaeological and Anthropological Sciences, 2022, 14(5)
[46] 邓振华.金沙江流域农业的形成与早期发展[J].中华文化论坛,2023, 1: 142-153+190
[47] 王洋,南普恒,晓毅,等.相近社会等级先民的食物结构差异:以山西聂店遗址为例[J].人类学学报,2014, 33(1): 82-89
