高光谱遥感在植物多样性研究中的应用进展与趋势
2024-01-01张艺伟郭焱培唐荣唐志尧
张艺伟,郭焱培,唐荣,唐志尧
北京大学 城市与环境学院 北京大学生态研究中心,北京 100871
1 引言
植物的生物多样性对群落稳定和生产力具有极为重要的作用(Hector 和Bagchi,2007;Schaub等,2020),对于整个生态系统而言,植物多样性保证了消费者的食物供应、多样的栖息地环境以及对植物物候和水循环的调节能力(Mi 等,2021)。生物多样性会受到多种生物和非生物因素的影响,并在空间分布上呈现出一定规律性,例如全球尺度下多样性随纬度梯度变化的格局,以及局域尺度下多样性随气候梯度变化的格局等(Gillman 等,2015)。近年来,由于人类活动和气候变化导致物种灭绝事件频发,全球生物多样性丧失加剧,成为危害自然环境和人类社会的重要因素之一。
了解不同空间尺度生物多样性的格局是生物多样性保护和管理的基础(Pressey 等,2003)。然而现有的物种分布记录主要源于野外调查数据,数据点的空间分布难以全面表征实际的分布。遥感技术能在极短时间内无接触、无破坏性地大面积采集地面信息,在生态学领域受到越来越深入和广泛的应用,如地物类型识别和植被分类(Hansen 等,2000),植被特征的定量估测及其空间格局制图(Hakkenberg,2017;Zheng 和Moskal,2009)等。遥感技术是目前获取大尺度空间数据,监测陆地生态系统变化的最有力手段(Darvishzadeh 等,2011),可以直接提供植被、地形、气候数据进行后期分析,保证了变量间的观测尺度平衡(Ustin和Gamon,2010)。目前各类多光谱遥感数据已经被广泛应用于植被监测,但因其覆盖的光谱区间较少,对植被的研究仅限于使用宽波段红光、近红外以及中红外波段的反射特征,对于植被长势和生理性状探测不足,难以满足生态学研究和应用的需求(Ollinger,2011)。高光谱成像技术的出现为更深入的遥感植被信息提取提供了有力支持(彭羽 等,2019)。
高光谱遥感HRS(Hyperspectral Remote Sensing)全称为高光谱分辨率成像光谱遥感,是将目标对象与电磁波相互作用的高分辨率光谱特征作为研究对象的光学遥感技术(Goetz等,1985)。根据研究需求的不同,选取目标像元的特定波段或特定反射光谱特征可辨别其地物大小、丰度、物质组成等信息(Farrand 和Harsanyi,1995)。最早的高光谱成像源于20 世纪80 年代的机载可见光/红外光谱仪AVIRIS(Advanced Visible Infrared Imaging Spectrometer),其光谱分辨率达到10 nm,共有224 个窄波波段,覆盖了400—2400 nm 的光谱区间,是高质量高光谱信息采集方式的开端(Goetz,2009)。近年来,越来越多的高光谱遥感卫星平台投入使用,以满足人们对植物多样性的空间制图和实时监测的需求(表1)。
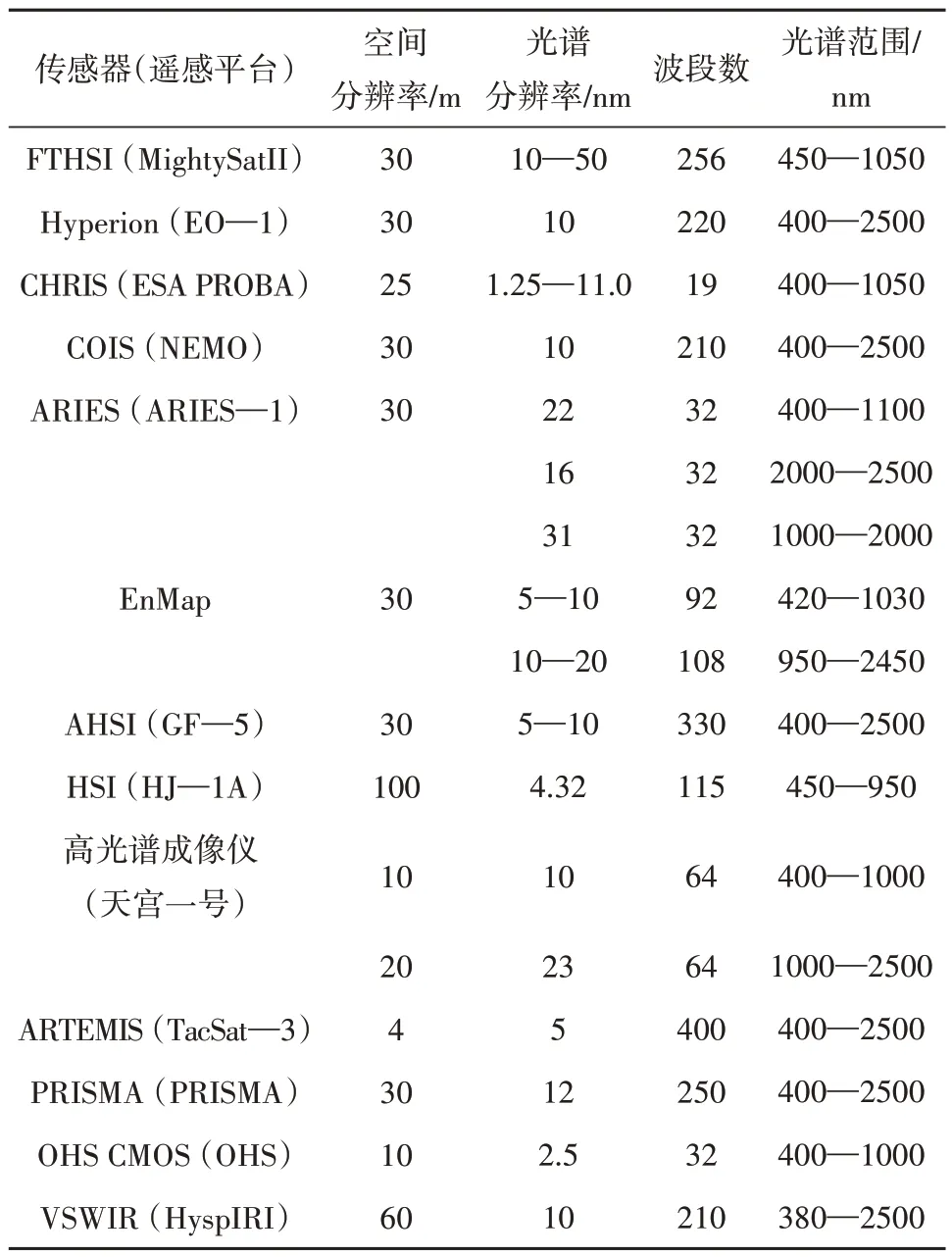
表1 部分在轨卫星高光谱传感器技术参数Table 1 An overview of operational hyperspectral sensors’ technical parameters
高光谱传感器具有精细光谱分光和图谱合一的技术优势,光谱分辨率的提高加强了对植物物种、功能和基因水平细微差异的探测和识别(Asner,1998;Goetz 等,1985)。结合实地调查,高光谱遥感可以在大空间尺度下探测并提取叶片生化特性、冠层结构特征等植物功能性状(Wang和Gamon,2019;Houborg 和Boegh,2008;Asner等,2015b),甚至可用于识别不同树种、估测部分基因水平特征(Fassnacht 等,2016;Féret 等,2011;Wessman 等,1988;Schlemmer 等,2013);高光谱遥感数据还被广泛地应用于植被监测(Thenkabail 等,2004;Asner 和Heidebrecht,2002;覃先林 等,2020)、植物功能性状制图(Skidmore等,2010;Chan 和Paelinckx,2008)、生物多样性估测(Gholizadeh 等,2019)等领域。2003 年首次出现了利用高光谱数据进行植物多样性指标提取的研究(Held等,2003),其后的20年,基于高光谱的植物多样性研究受到越来越多的关注,为定量化地进行植物多样性监测提供了新的助力。
目前,高光谱遥感已经广泛应用于大尺度的生境制图、植物功能和谱系多样性反演等多层次的植物多样性研究(Wang和Gamon,2019)。高光谱遥感手段大大减少了人力、物力的消耗,保证了多样性监测的时效性,满足了对大尺度植物群落多样性格局动态监测的要求。
根据多样性度量要求的不同,高光谱遥感对多样性的反演方法可以分为基于光谱特征的直接估测和基于定量反演的间接估测。前者主要通过建立光谱异质性与生物多样性的直接关系实现多样性估测,并已经应用于物种、功能性状和系统发育的多样性估测(Thenkabail 等,2018);后者主要通过高光谱数据的精细光谱特征定量提取植物性状或植被参数,进一步关联生物多样性指标,例如定量反演特定冠层性状并结合相应的多样性指标实现功能多样性的估测(Durán 等,2019),通过生境制图间接表示物种分布格局(Turner 等,2003)等。
2 基于光谱变异假说的植物多样性直接估测
不同植物群落间的物种多样性可以通过其结构和生化特性体现出来,并产生可区分的光谱(吴炳方 等,2020)。高光谱遥感数据能够以上百个连续光谱波段对观测目标实现同时成像,其光谱分辨率和响应波段范围相比于多光谱遥感数据都有了极大的提升,因此可以更精细地呈现和量测物种多样性差异导致的光谱差异,并在很大程度上表征传统遥感手段难以描述的物种多样性指标。
2.1 基本原理
高光谱特征直接应用于物种多样性反演的理论基础是光谱变异假说SVH(Spectral Variation Hypothesis)(Palmer 等,2002),这一假说认为生物多样性与光谱反射率差异间存在一定的联系。冠层生化性状的差异主要源于群落内多样的物种组成(Townsend 等,2007),而且这种差异能够被高光谱信号响应,因此直接建立光谱反射率差异与群落生物多样性差异的联系在理论上是可行的(Wang 等,2018a)。不同植物的化学、解剖、形态、分类特征的差异共同影响着光谱特征,因此光谱特征的差异可以在一定程度上直接表征植物多样性的高低(Lausch 等,2016)。Asner 和Martin(2009)比较了热带森林树种的多个冠层生化性状组合及其光谱特征,发现当引入足够多的生化性状时,性状值组合可以与物种唯一关联;在此基础上,物种识别的问题转化为性状定量反演。SVH 假说通过直接建立光谱反射率曲线特征与植物群落多样性间的关系,来实现不同植物群落物种多样性相对大小的估算和比较(图1)。
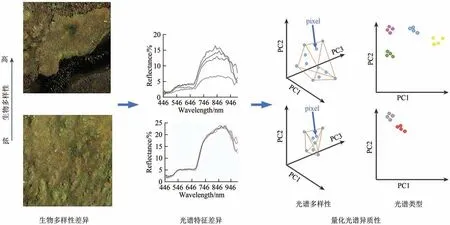
图1 基于光谱特征的多样性提取原理Fig.1 Schematic illustration of the extraction of biodiversity from spectral traits
高光谱信号对生化性状的差异响应是生化性状定量反演的基础,对光谱信号异质性的直接量化方式可以归纳为两类。一是应用聚类思想,如“光谱类型”(optical types)或“光谱物种”(optical species)方法,该类方法通过对光谱特征进行聚类,得到植物群落内具有相似生化和结构特征的有限物种的光谱信息团聚体,每个团聚体即称为“光谱类型”或“光谱物种”(Féret 和Asner,2014;Asner 和Martin,2009;衣海燕 等,2020;Zhao 等,2018)。二是基于光谱空间来量化光谱异质性或光谱多样性(spectral diversity)(Asner 和Martin,2009),即假定光谱空间是一个多维空间,由筛选出的包含主要信息且相互正交的波段来定义各坐标轴,影像在多维空间形成的包络体大小作为光谱多样性大小(Schweiger 等,2018)。度量不同植物群落影像光谱在光谱空间占据的范围和填充模式,可用于比较其群落物种多样性的相对高低(Rocchini等,2010)。
SVH 假说在物种丰富度较高的热带和亚热带森林地区的实验中,直接估测都表现出令人满意的精度(Asner 和Martin,2009;Zhao 等,2016)。因此该方法在大空间尺度下植物多样性的识别和比较中具有较大优势。
2.2 主要方法
光谱变异假说将植物多样性转化为光谱特征的异质性问题,即通过度量不同植物群落的光谱异质性程度来比较多样性高低。基于这一假说,研究者提出了不同的方法来量化光谱异质性(表2),如应用全谱段信息评估光谱多样性的变异系数法CV(Coefficient of Variations),光谱角法SAM(Spectral Angle Mapper)和信息离散度法SID(Spectral Information Divergence),以及需要进行波段降维处理的包络体法CHV(Convex Hull Volume)和体面积法CHA(Convex Hull Area)。这几种方法都是以群落样方为基本单位,通过比较各个像元光谱与平均光谱的差异程度来度量样方内光谱多样性。变异系数法计算了样方内光谱特征参数的变异系数或所有像元各波段的变异系数均值作为光谱多样性指标,变异系数越高表明样方内像元光谱多样性越高,是目前较为常用的光谱多样性计量指标(Lucas 和Carter,2008)。光谱角法是将像元所有波段的光谱响应转化为同等维度光谱空间的光谱向量,通过计算样方内向量间夹角(光谱角)作为光谱差异的指标(Kruse 等,1993),光谱角数值越大,代表群落的光谱异质性越高,一定程度上说明该群落生物多样性较高。信息离散度法同样针对光谱向量进行比较,将向量间离散度大小作为光谱差异大小(Chang,2000)。

表2 常用光谱多样性指标Table 2 Common spectral diversity indices
包络体法首先选取包含主要波段信息的前3个主成分,在3个主成分构建的光谱空间内计算样方像元占据的包络体体积,包络体体积越大表明样方光谱异质性越高(Dahlin,2016)。体面积法同样借鉴了包络体的思想,该方法定义了二维坐标空间,以每个波段平均反射率和像元反射率作为两轴,第N个点表示第N个波段平均反射率以及样方内与平均反射率差异最大的像元反射率对应的坐标位置,N个点形成的包络面的面积越大表明该样方光谱异质性越高,面积接近零时,表明样方各像元反射率几乎等于各波段反射率均值,在二维空间内近似形成一条直线(Gholizadeh 等,2018)。近年来,随着机器学习和深度学习的广泛应用,相关算法也被应用到植物群落多样性与光谱异质性直接关系的构建中,例如随机森林算法、支持向量机、高斯过程回归、神经网络模型等。
不同层次的植物群落多样性研究证实,光谱多样性能够直观地表征群落内的植物多样性(John等,2008;Wang 等,2016)(表3)。Gholizadeh等(2019)发现机载高光谱影像提取的不同草地群落光谱多样性与实地调查的α 多样性显著相关,且在不同群落类型中、不同空间尺度及分辨率下都有较好表现。在不同植物群落进行的实验证明,光谱多样性能对物种丰富度、Rao’s Q 指数等多样性指标实现较好的反演(Khare 等,2019;Polley等,2019;Gholizadeh 等,2019),因此在某些难以调查的区域,可以直接用光谱多样性表征其物种多样性。

表3 两类高光谱反演植物多样性的应用实例Table 3 Examples of two methods of plant diversity retrieval through hyperspectral remote sensing technology
近年来,功能和谱系多样性理论的发展使得生物多样性的研究与监测向着多维度的方向迈进,生物多样性的遥感估测也随着技术的进步而拓展到不同的维度(Kong等,2019)。高光谱技术在植物遗传和系统发育多样性方面同样具备应用潜力。精细的光谱特征可以在一定程度上捕捉植物系统发育差异,进而实现对遗传多样性的探测。例如,Cavender-Bares 等(2016)分析了栎属多个树种的叶片光谱与谱系信号,发现光谱相似性与谱系相似性显著相关,全光谱信息能够识别和区分同一树种因种内变异而产生的不同种群。
3 基于定量反演的植物多样性间接估测
高光谱技术实现了植被遥感研究由定性到定量的突破,全谱段的光谱反射率信息以及“红边”位移监测能够敏感探测冠层养分、植被长势和环境胁迫等信息(Dawson 和Curran,1998)。植被高光谱定量遥感为植物功能性状研究提供了助力,通过定量反演的冠层性状计算群落功能多样性是重要的多样性间接估测手段。大尺度的植物性状和功能多样性格局的获取将有助于深入了解植被群落构建机制、生物多样性与生态系统功能的关系等生态学热点问题(Cadotte 等,2011)。除了在功能多样性方面的估测,高光谱数据在地物分类方面具有更高的识别能力,因此能够实现更精细的生境制图,借此推断物种分布的空间格局。连续光谱响应信息结合多种植被指数可以实现对植被表观和遗传信息的追踪和遥感探测。
3.1 基于性状反演的功能多样性间接估测
3.1.1 植物功能性状定量反演
植物功能性状是植物对环境适应策略的综合表达,特定环境条件下群落内各物种功能性状的趋向性体现了群落对外界环境的响应(Chave 等,2009;Wright 等,2004)。对植物功能性状数据的传统采集方式以实地调查为主(郭庆华 等,2018)。然而这一方法耗时费力,在较大空间范围内难以统一数据规范,并且具有破坏性和时滞性,因此难以大面积应用(Schneider 等,2017)。遥感技术获取大尺度空间数据的能力在一定程度上弥补了实地调查的缺陷,并将叶片和冠层层次的植被性状提升到了群落空间层次(Durán 等,2019)。冠层结构参数、叶片含水量指标、干物质含量等重要的植物功能性状,会在较宽光谱范围产生响应特征(Ollinger,2011),例如叶片含水量在近红外和短波红外具有明显吸收特征,叶绿素a 和b 在蓝波段和红波段形成明显吸收特征。高光谱遥感技术具备更宽波长范围的连续光谱覆盖,大大提升了遥感对多种植物功能性状的探测能力(表4),并且对空间数据的同步获取保证了数据精确性和标准统一性(Schweiger等,2017)。
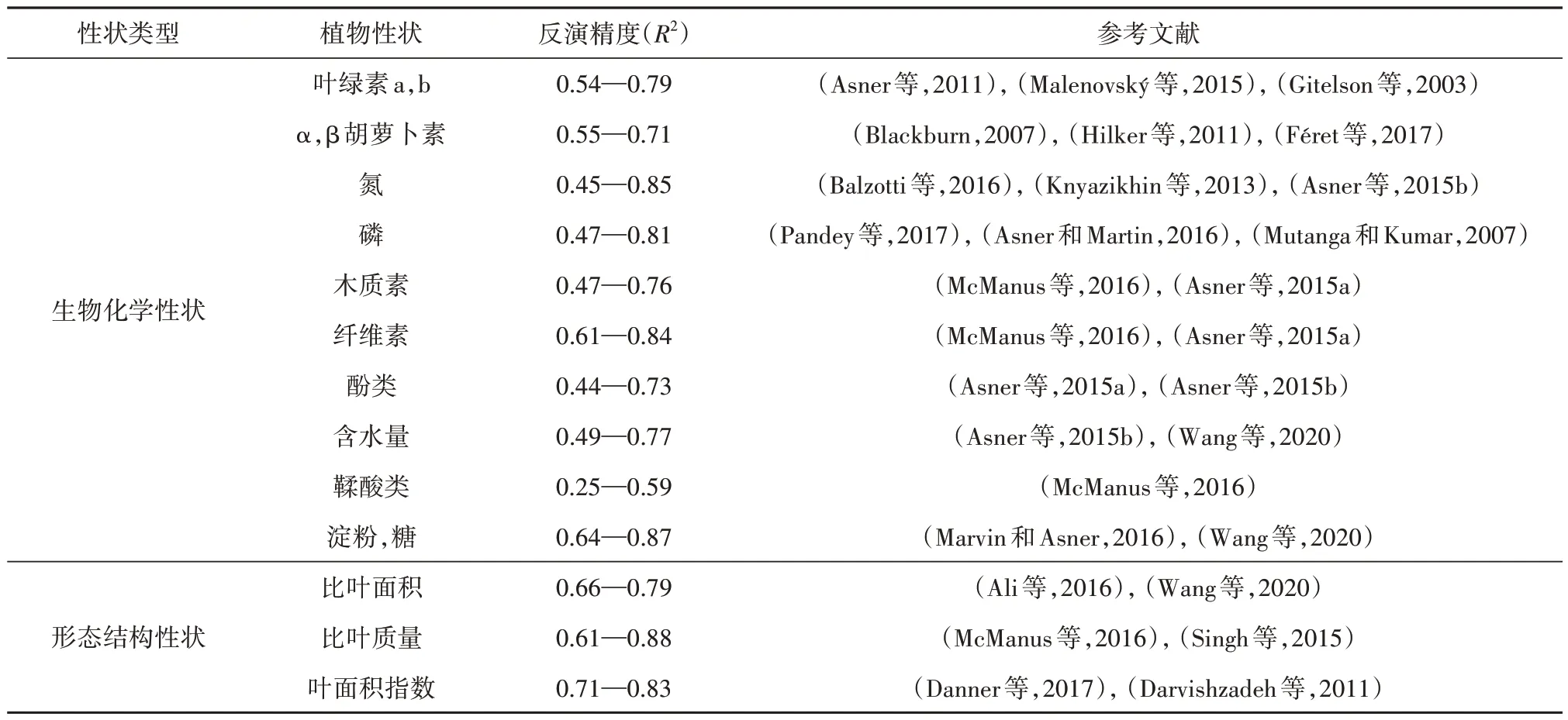
表4 基于高光谱遥感可探测的植物功能性状Table 4 Detectable plant functional traits based on hyperspectral remote sensing
基于遥感信息定量化反演植物功能性状的常用模型方法包括经验统计模型和物理模型(Skidmore 等,2010)(表3)。其中经验统计模型在区域尺度上应用更为广泛,并且对不同的性状都表现出了较好的预测能力(Singh等,2015)。如Wang 等(2019)通过建立实测样地数据和敏感光谱之间的偏最小二乘回归模型PLSR(Partial Least Squares Regression)和高斯过程模型GPR(Gaussian Processes Regression),提取了Cedar Creek 生态系统保护区草地样地的15 个性状。机器学习方法如随机森林模型(Zald 等,2016)、神经网络模型(Skidmore 等,2010)、支持向量机(Chen 和Hay,2011)等经验统计模型在对高维数据的处理中具有极大的优势。同时,这类算法需要较大的实测样本量以保证精度,模型反演质量的普适性也需要更多来自不同植被类型的数据校验(Gholizadeh等,2018)。对于较为均质的植被,统计模型估测结果的不确定性可以控制在较低范围内,能够满足生态学研究的数据精度需求(Verrelst 等,2016),因此得到了较为广泛的应用。
物理模型方法基于辐射传输理论实现。辐射传输模型是对给定环境条件下植被冠层的不同组分对电磁波的吸收和散射的模拟,通过将辐射传输模型逆运用来实现性状反演(图2),因此其稳健性(Robustness)和全局性更强(Wang 等,2018b)。常用的辐射传输模型包括提取叶片性状的PROSPECT 模 型(Jacquemoud 和Baret,1990)和提取冠层性状的SAIL 模型(Féret 等,2017;Jacquemoud 等,2009)等。由于模型的参数化受到植被条件、土壤背景等因素的影响,其反演精度也有局限(Yebra和Chuvieco,2009),并且模型反演精度单纯由输入参数决定,缺少相关生态条件的限制,某些异常参数会使得结果偏差极大(Wang 等,2019),导致反演精度不够理想。例如,在不同土壤背景和光照条件下,极高和极低叶面积指数LAI(Leaf Area Index)可能会呈现相同光谱反射曲线特征,但是当限定干旱条件时,高LAI 显然是不现实的结果,因此可以通过设定LAI阈值来提升模型的精确度(Jurdao等,2013)。
近几年随着近地面无人机遥感技术的迅速发展,高光谱与激光雷达的协同应用也成为森林遥感关注的焦点。激光雷达获取的三维点云数据能够提取树木结构性状,结合高光谱技术获取的冠层生理性状,可以全面刻画森林群落的功能性状分布规律。相比于像素级别的植物性状反演,基于个体的性状数据可以更强地关联生态系统功能,并且能够很好地反映森林群落的生态过程(Zheng等,2021;Chadwick和Gregory,2016;Martin等,2018)。
3.1.2 功能多样性度量
不同的生态过程、干扰和胁迫会影响或改变植物的系统发育和表观遗传过程,使得植物在性状上产生不同的响应,例如,植物会通过改变生物化学性状和(或)结构特性来适应外界干扰等,从而形成了群落内植物功能性状在时间和空间上的变化。研究显示,功能多样性作为对群落内植物功能性状差异的表征,与生态系统功能直接相关(Cadotte 等,2011),因此越来越多的研究开始关注功能性状及其多样性在生物多样性与生态系统功能BEF(Biodiversity and Ecosystem Functioning)关系中的作用(Díaz等,2007)。
功能多样性的测定和表示方式众多,前文提到的“光谱类型”法基于SVH 直接构建了光谱特征与功能类型的联系(Ustin 和Gamon,2010),而间接估测法则采用不同的功能多样性指标计算遥感反演的植被性状的多样性水平。其中常用的指标包括反映功能性状丰富程度的FD 指数(Petchey 和Gaston,2002)、功能丰富度指数FRic(functional richness)(Villéger 等,2008;Mason 等,2005)和TOP指数(trait onion peeling)(Fontana等,2016),反映功能性状均匀程度的功能均匀度FEve(functional evenness)以及反映功能性状分化差异的功能分散度FDiv(functional divergence)(Villéger等,2008)、功能离散度Fdis(functional dispersion)(Laliberté 和Legendre,2010)和Rao’s Q 指数(Rao’s quadratic entropy)(Botta-Dukát,2005)等。以上多样性指标的计算也因性状反演基本单位的不同而有所差别,可分为像素级别和个体级别的功能多样性。如以FDiv 为例,计算基于像元或个体的群落功能性状离散程度:
式中,S代表群落内像元(或个体)数目,dGi代表第i个像元(或个体)到所有像元(或个体)包络体的重心距离,代表所有像元(或个体)到重心的平均距离。
近年来功能性状的研究更加强调种内性状变异的重要性(Cadotte 等,2015),相关的功能多样性评估方法包括Frim 指数(Schleuter 等,2010)和性状概率密度TPD(Trait Probability Density )(Carmona 等,2016),这两种方法考虑了种内变异情况,但同时对数据反演结果也提出了质和量方面更高的要求,因此基于高光谱数据的反演应用还相对较少。
3.2 基于光谱指数的多样性间接估测
高光谱数据的光谱特征参量和光谱指数是经验模型常用的光谱信息指标。植被光谱反射率在可见光和近红外波段之间急剧上升,形成所谓的“红边”特征,对红边特征参量化来追踪植物冠层性状变化是植被遥感重要的研究内容。整合多个宽波段信息的植被指数突出了特定地物的光谱信号,在一定程度上可以反演植被冠层特征(Gutman,1991),例如应用较为广泛的归一化差值植被指数NDVI(Normalized Difference Vegetation Index)、增强植被指数EVI(Enhanced Vegetation Index)。针对特定研究需要也发展了多样的植被指数,例如优化的土壤调整植被指数OSAVI(Optimized Soil-Adjusted Vegetation Index)(Rondeaux等,1996)、增强型土壤调节植被指数MSAVI(Modified Soil-Adjusted Vegetation Index)(Qi 等,1994)、调节型叶绿素吸收比率指数MCARI2(Modified Chlorophyll Absorption Ratio Index)(Haboudane 等,2004)等。利用全波段光谱信息的全谱段植被指数法VIUPD(Vegetation Index based on the Universal Pattern Decomposition method),能够反映更细微的植被信息(She等,2014)。
大量研究证实,借助光谱指数可以与植物遗传、功能和物种水平多样性参数建立一定联系,实现多样性的间接估测。例如,可借助NDVI 实现对草地(Wang等,2016)、灌丛和森林(Fairbanks和McGwire,2004)群落物种丰富度的估测。在遗传多样性水平,Blonder 等(2020)通过随机森林算法对美洲山杨(PopulustremuloidesMichx.)叶片和冠层光谱计算得到的NDVI 进行建模,成功实现了对染色体不同倍性的识别。
4 基于高光谱数据提取植物多样性的应用进展
利用高光谱进行植物多样性的研究受到了越来越多的关注。在Web of Science(www.webofknowledge.com)所有数据库,结合主题“hyperspectral”或“imaging spectroscopy”以及“plant biodiversity”或“plant diversity”进行检索发现,近20 年来,相关出版物条目(左侧Y 轴),以及相关出版物在当年关键词搜索得到出版条目总数中所占比例(右侧Y 轴)呈现逐年增长的趋势(图3),体现出高光谱遥感技术在植物多样性研究中巨大的应用潜力。多源数据融合是当今大尺度植物多样性研究的发展趋势:以无人机遥感为代表的近地面遥感技术的推广,解决了卫星和航空遥感数据时空分辨率粗糙的问题;高光谱遥感技术与激光雷达技术的协同应用,保证了植被生化性状和结构性状的同步获取;叶绿素荧光遥感技术的不断发展,建立起了遥感信息与植被冠层光合作用的直接联系,能够从物种—生产力假说(species-productivity hypothesis)角度验证植物多样性格局;计算机可视化和深度学习算法为海量数据的处理和分析提供了技术支持;通量监测技术为遥感数据验证提供了精确的冠层尺度信息(Inoue等,2008)。高光谱技术与这些新兴技术的协同应用能够为不同尺度和维度下植物多样性问题的研究提供有力技术支持,例如入侵物种识别、植物群落构建机制、大尺度物种分布格局以及大尺度BEF 理论的验证等生态学热点问题。
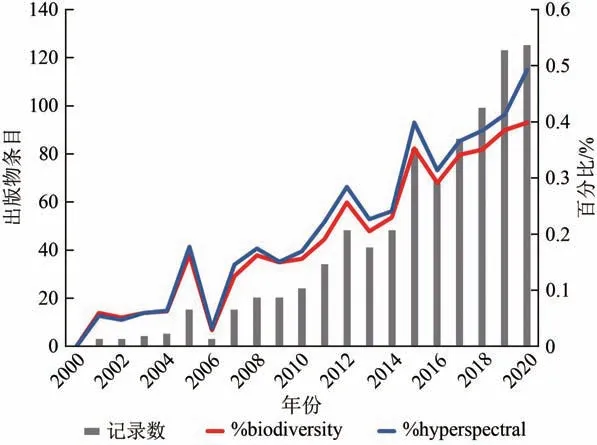
图3 以“高光谱”及“植物多样性”为主题的年出版物数量,及其所占比例Fig.3 The yearly numbers of publications with the topic of‘hyperspectral’ and‘ plant biodiversity’and percentages of the total numbers of publications
研究表明,目前全球物种入侵形势严峻。入侵物种的快速扩散和传播,通过种间竞争占据资源并改变了原有生境条件,极大威胁了本地物种的生存(Ricciardi,2007)。高光谱遥感数据丰富的光谱信息,能够通过探测植物生化性状(Khanna 等,2011)、物候特征(Andrew 和Ustin,2008)等方面的差异实现对入侵种的监测,例如Asner等(2008)基于机载高光谱图像检测不同树种色素含量、营养水平和结构特征,实现了对夏威夷森林入侵种和本地种的区分。海量的光谱信息使得对同一植物功能群内不同物种的识别成为可能,从而有助于揭示物种入侵的具体过程及其对当地环境的影响。
在群落生态学研究中,高分辨率的近地面高光谱技术可以将多样性格局监测从物种分类维度推广至物种、功能甚至基因多样性维度(Schaepman 等,2009),因此在生物多样性格局的研究中具有独到的技术优势。如Durán 等(2019)基于高光谱数据间接估测了热带森林群落功能丰富度和离散度指标,分析了随海拔梯度功能多样性指标的变化和不同空间尺度下群落构建机制的差异。高光谱遥感结合激光雷达技术能够在结构复杂的森林群落同步采集植物群落结构、功能性状和地形信息(郭庆华 等,2016a;Hakkenberg,2017),多源遥感技术协同反演是未来全球视野下植物多样性研究的重要发展方向(李玉美 等,2021;Jetz 等,2016),为生态学研究提供了丰富的数据资源(图4),满足了从个体到宏观尺度的研究。
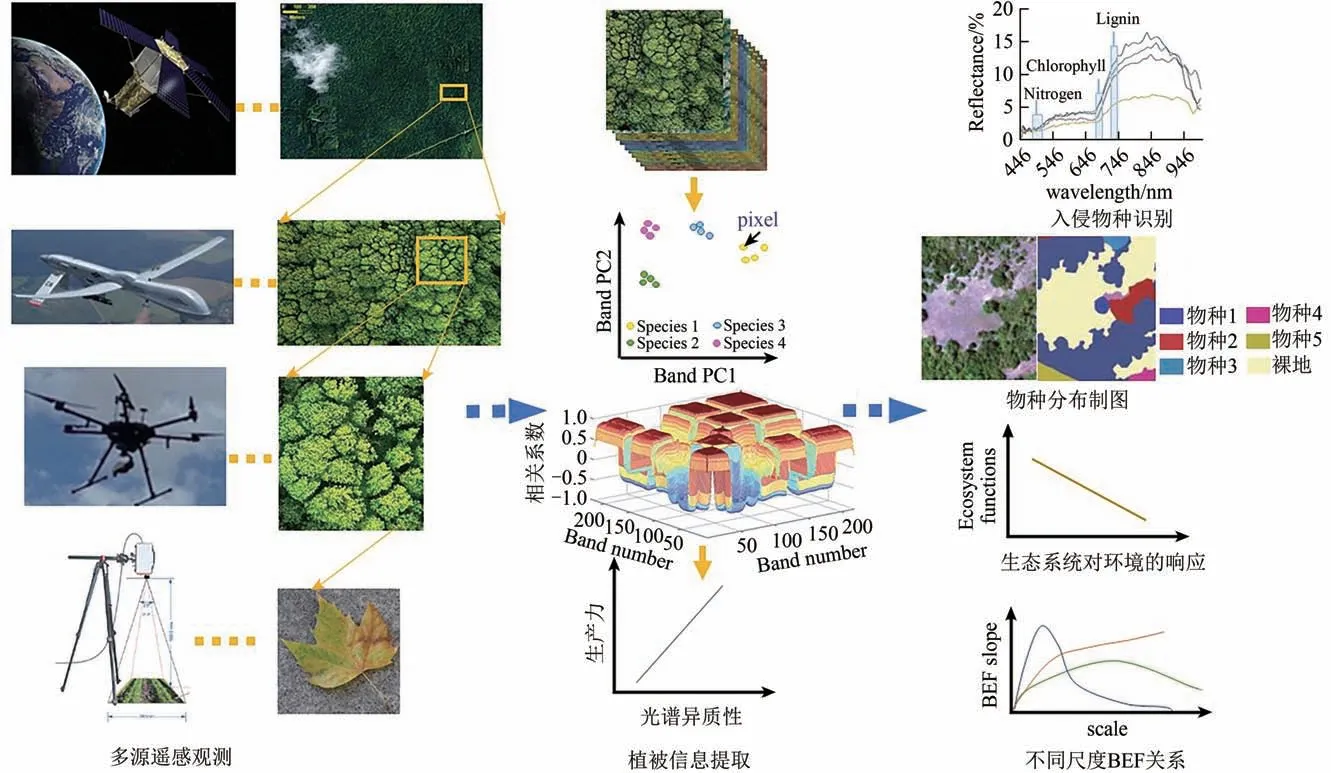
图4 遥感高光谱生物多样性提取的应用(部分改编整理自Thenkabail等,2018)Fig.4 Application of biodiversity retrieval based on hyperspectral remote sensing data(partly adapted from Thenkabail et al.,2018)
不同植被类型的样地实验揭示了生物多样性对生态系统功能的促进作用(Tilman 等,2006),但同时这种促进关系随着空间尺度和环境条件发生变化(Fraser 等,2015;Thompson 等,2018),因此基于群落调查的空间大尺度BEF 关系研究也相继展开(Liang 等,2016;Guo 等,2019;Stein等,2014)。然而其中多样性格局的获取多依赖于实地调查或清查数据的整合,采样标准难以统一,因此可能造成难以消除的系统偏差(Jetz 等,2016)。目前卫星遥感数据可以通过提取土地覆盖差异来间接表征大尺度的生境多样性或物种组成差异(Wilson 等,2012;Oehri等,2020),而高光谱遥感技术可以直接满足对功能性状反演和物种识别的需要,在BEF 研究中表现出极大的潜力,如基于高光谱可见光波段对草地群落反演的功能丰富度结果表现出与生态系统生产力间较好的联系(Zhao等,2021)。
5 高光谱植物多样性应用的局限性
尽管高光谱遥感在植物多样性研究中得到了广泛应用,但它仍存在一定的局限性。遥感手段虽然满足了不同空间尺度研究的要求,但是在调查郁闭度较高的森林和灌丛群落时,高光谱遥感技术只能获取冠层上部的光谱反射信息,对下层植被光谱信息的获取则较为困难。此外,由于大空间尺度冠层高光谱数据的空间分辨率相对粗糙,像元尺寸难以满足个体识别的要求,因此在反演草地甚至森林群落的性状和多样性时,需要借助地面定量光谱辐射计采集的叶片光谱作为参考。因此叶片尺度和冠层尺度的数据转换也是目前植物多样性反演面临的一大挑战。
相比于传统的多光谱遥感数据,高光谱数据的光谱分辨率大幅度提升,数据的处理更为复杂。以EO-1 卫星搭载的Hyperion 高光谱传感器为例,对于同一区域获取的遥感影像,其数据量相比Landsat TM 影像增加了37 倍(Thenkabail 等,2018),数据处理的工作量因此呈几何级数增长,分类的Hughes 现象不可避免。而对于特定植被而言,具有区分性的光吸收特征通常与离散分布在全光谱范围的几个波段相关,例如,在400—700 nm可见光范围内,叶片色素呈现强吸收特征;而在近红外和短波红外区间,叶片结构特性如细胞壁厚度、胞间隙、表层蜡质等对电磁辐射有更强的吸收效应(Schweiger 等,2018)。此外随着研究对象的不同,高敏感波段的组合也有所差异。因此利用高光谱数据进行植被探测尤其是对特定性状进行模拟估算时,需要在保证有效信息尽量保留的同时减少信息冗余。如何针对研究目标和植被类型,选取有效波段建立起光谱—性状的联系是后续数据处理中值得考虑的问题。
在分析方法上,由光谱特征直接关联植物多样性的方法减少了对大量实测数据的依赖,因此在监测大尺度植物多样性空间变化时具有极大的应用潜力,但这一方法难以直接从机理上建立光谱多样性与生物多样性的关联,使得植物群落光谱多样性的生态学意义有待进一步阐释(Cavender-Bares 等,2020b)。其次,反演大尺度植物群落多样性格局对高光谱数据具有较高的要求,而目前的高光谱卫星遥感仍处于发展的初级阶段。一方面,覆盖全球的高光谱遥感卫星系统还不够完善(童庆禧 等,2016),卫星的重访周期长使得高光谱数据获取的频率较低,难以满足实时获取数据和动态监测的要求;另一方面,现有的航天高光谱数据的空间分辨率较低,在多样性较高的区域,粗糙的空间分辨率会导致大量混合像元的出现,因此难以精确描述植物的多样性(Fassnacht等,2016)。
近年来以无人机为代表的近地面遥感监测平台不断发展,在一定程度上弥补了从航天遥感到样地调查之间数据尺度的空缺(郭庆华 等,2016b)。但是实际使用中,无人机平台的稳定性会受到气候和地形条件、驾驶员操控能力以及飞行政策等各种因素制约,并且无人机高光谱数据采集和后期解译主要依赖于人工设置,这也限制了数据应用的时效性和广泛性(纪景纯 等,2019)。在多源高光谱遥感数据的综合应用方面,如何实现不同区域遥感数据的背景归一化,以及如何结合运用不同空间尺度的机载数据与星载数据也是未来研究的重点(林珲和张鸿生,2021;Jetz等,2016)。
除此之外,传感器灵敏度、暗电流以及杂散光会导致相干噪声或随机噪声的产生,且高光谱数据的信噪比是否足以支持重要光谱特征的识别仍存在不确定性(Cavender-Bares 等,2020a)。当进行大尺度植物性状制图时,需要整合不同遥感平台的高光谱数据,如何校准和统一具有不同中心波段、不同光线条件下的成像数据也是今后数据处理工作中需要改进的方向之一。
目前成像光谱学的多样性研究仍处于起步阶段,相信随着多领域技术和理论的融合,人们将能够更好地实现不同尺度植被监测,进而更好地探究不同自然环境中生物多样性的动态变化规律以及对生态系统功能和服务的作用机制,以此制定合理的措施指导全球生物多样性保护(Fei 等,2018)。
