短期模拟增温对短尖薹草生长与繁殖的影响
2023-12-28李铭恩何池标李美瑶
李铭恩,何池标,李美瑶
(中南林业科技大学 生命科学与技术学院,湖南 长沙 410004)
随着人类活动日益加剧,全球气候持续变暖,未来20年的全球气温或将上升1.5~2.0℃[1,2];其中中国是全球受到气候变化影响最大的国家之一[3]。气候变暖对淡水湖泊生态系统造成巨大影响[4],其生态系统结构与功能将发生改变。湖泊湿地在气候调节、涵养水源、固碳增汇等方面有重大作用[5]。洞庭湖是我国第二大淡水湖泊,短尖薹草(Carexbrevicuspis)作为洞庭湖优势群落,是鱼类和鸟类的觅食场、产卵场及栖息地[6],对维持湖区生态系统功能具有重要作用。
模拟增温是目前研究植物应对气候变化的重要手段,对探究植被生长与环境因子之间的关联提供了重要依据[7-12]。在模拟增温条件下,黑褐薹草生理指标与温度变化显著性水平极高[13];模拟增温后,温度会对植物生长演替产生影响[14]。植物的分蘖数、株高、叶片数等生理指标以及相关繁殖指标均可作为研究其演替趋势的直观依据,同时生物量的变化也是植物适应环境的结果,可反映其生长状况[15,16]。
本文通过对洞庭湖短尖薹草进行模拟增温,研究其在模拟增温下的生理指标变化过程与繁殖情况,揭示增温对短尖薹草群落的影响,以期为洞庭湖将来的植物群落变迁以及生态系统结构变化提供科学依据,并可为亚热带湿地生态系统修复提供思路。
1 材料与方法
1.1 研究地点
本研究在湖南省长沙市中南林业科技大学西园温室进行,所在地点为亚热带气候区,终年湿润,夏季炎热,降水集中在4~7月,年均气温达17.4℃。短尖薹草与培养土壤均取自于洞庭湖。短尖薹草根茎粗短,秆高20~55 cm,三棱形且平滑坚硬,基部有深棕色分裂成纤维状的老叶鞘,叶宽5~10 mm,平张,叶顶端逐渐狭窄,苞片短叶状且具长鞘。由于洞庭湖每年6~10月会被周期性浸淹,短尖薹草生命周期有3个阶段:12月为第一生长季,翌年1~2月为低温休眠季,3~5月为第二生长季,花期在4~5月,共6个月。故本研究选择在1~6月进行。
1.2 试验设计
在西园温室内搭建试验平台,温室为玻璃材质,植物和土壤均从岳阳市洞庭湖湿地中原位取回,盛装仪器为截面积120 cm × 80 cm的长方形胶桶。试验设置24桶植物随机排放,并采用相同功率的悬挂红外加热辐射器实现土壤增温(图1),对照组不作处理。增温组和对照组各12组。
1.3 试验方法
1.3.1 温度测定
土壤温度与大气温度采用TMS-4土壤温湿度记录仪测定,将温度计插入土壤深度约10 cm处;同步测定气温和土温变化。
1.3.2 生长监测
为模拟野外状态,本次试验进行为期6个月的监测,2022年1~6月,间隔1个月进行一次生长观测,记录薹草叶片数、分蘖数以及株高等生理指标。2022年的3月11日,第一次发现短尖薹草开花,每72 h进行一次繁殖指标统计。通过建立异速生长方程推算植物生物量的变化。
1.4 数据处理
通过获取气温和土温的日均温数据,比较增温组与对照组的增温效果。观测统计2组的分蘖数、叶片数和株高,比较增温条件对其的影响;生物量则根据分蘖数、株高、叶片数进行异速生长方程构建并进行推算。数据处理与统计使用Excel和RStudio软件,作图采用Origin 2022 和Excel。
2 结果与分析
2.1 温度变化
土温与气温均随时间变化而上升,但增温组均高于对照组。与对照组相比,土壤平均温度升高了1.11℃,大气升高了0.63℃,模拟增温量接近第六次IPCC报告预测的21世纪全球温度将升高1.5℃[1],说明本次增温效果较明显(表1)。
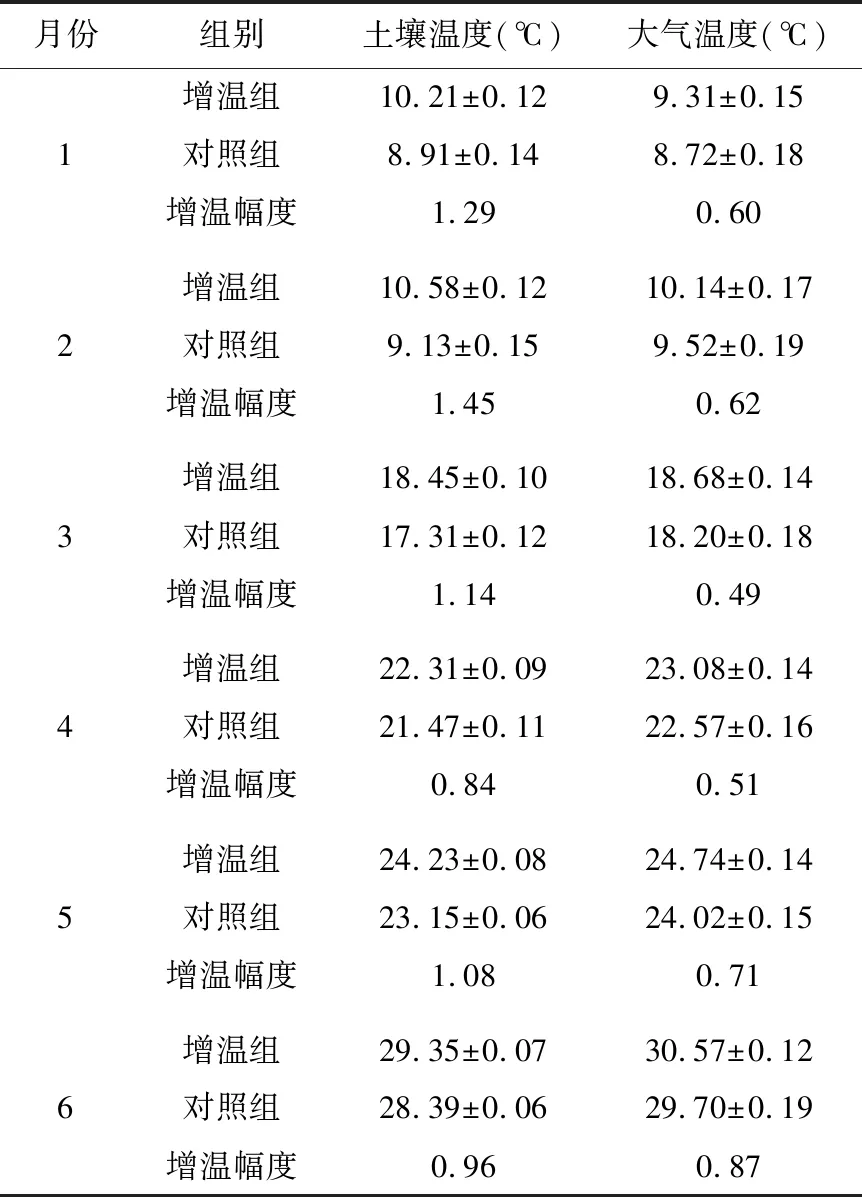
表1 实验平台增温组与非增温组的温度对比
2.2 生长性状变化
增温处理前3个月,植物株高呈持续生长趋势,但之后株高保持稳定不再上升(图2-a)。在增温开始前2个月,增温组的株高显著高于对照处理(P<0.001);但从3月开始,两组间的株高差异不再显著(P>0.05)。

图2 短尖薹草增温组与未增温组的生长性状对比
增温处理前3个月,植物分蘖数表现较为稳定,但自增温的4月之后,植物分蘖数随温度升高而显著增加(图2-b)。5月时未增温组的分蘖数已经超过增温组,并在6月呈现显著差异。
增温期间,植物叶片数呈现先增加、后降低的趋势(图2-c)。在增温的3-4月,叶片数达到峰值,而后开始降低。增温组和未增温组叶片数达到峰值的温度十分接近,但未增温组的平均叶片数峰值高于增温组。
2.3 异速生长方程构建
通过收获法测定植物的生物量,以Origin建立植物异速生长方程:
Z=1+ax2+by2+cx+dy
(1)
式中:Z为植株生物量,x为植株叶片数,y为植株株高,a,b,c,d为常数,本次计算中,a=0.00712,b=1.04542,c=-0.20677,d=0.02112。通过异速生长方程对生物量变化进行估算。
2.4 生物量变化
短尖薹草对增温的环境变化没有明显的生物量变化响应规律,但总生长周期内,前4个月增温组的生物量均高于对照组,而后期逐渐反之(图3),因此增温处理下薹草的生物量增加量低于对照处理。通过计算,增温使薹草在整个试验生长周期内生物量的增长量较对照组减少了6.72%。

图3 短尖薹草增温组与未增温组的生物量对比
2.5 有性繁殖
观察发现增温会使短尖薹草花期提前,增温组在3月11日开花株数已达到稳定值,而对照组在3月17日才达到稳定值。对增温植株进行统计,发现增温组在此周期开花率为3%,未增温组为6%,增温组雌穗平均数量(9.6个)低于未增温组雌穗(11.6个),增温组雄穗平均数量(3.3个)低于未增温组(5个),增温组开花率明显低于未增温组;同时,从雌雄蕊长度来看,增温组雌穗的平均长度显著低于未增温组,雄穗平均长度高于未增温组(图4)。
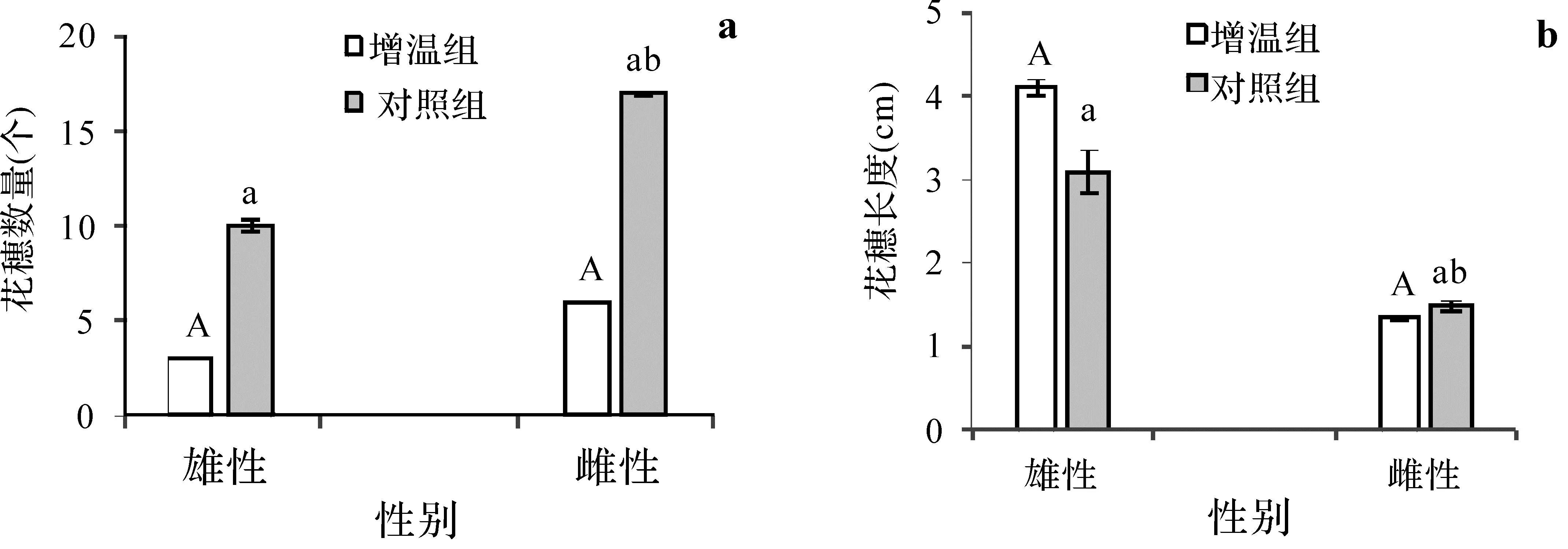
图4 短尖薹草雌雄植株的生殖与生长对比
3 讨论
3.1 增温对薹草生长的影响
此次模拟增温试验期间,增温组薹草的株高和叶片数在前几个月均持续增长,但从增温的5月开始降低且低于对照组,增温条件下的株高、叶片数均无显著差异,表明增温可短暂促进短尖薹草生长和分蘖数增加,但温度过高反而会对薹草生长产生生理抑制;同时,增温组和对照组分蘖数一直在增加,但二者并无显著差异,说明增温对薹草长期生长分蘖无显著影响。在植物与环境温度的相互作用中,温度一方面能改变植物的形态结构和生理生化特性;另一方面,植物对环境温度也有适应性和适宜范围。有研究表明,湖滨带湿地植物体现了生态因子对其功能性状的影响及其应对环境变化的适应机制[18]。温度作为最重要的生态因子之一,对植物的生长和发育起着至关重要的作用。在此之前,学者们研究增温对植物的影响有不一样的观点,一些学者认为升温对芦苇湿地细根的总量和生长速率提高显著[19],大气温度控制对杉木幼苗物候和生长有显著影响[20],增温对北美车前(Plantagovirginica)的株高有显著促进作用[21],这说明增温有助于地上部分的生长;而有些学者却认为增温对植物生长的影响并不显著[21],还有学者认为增温不仅使矮嵩草草甸的各种群生长期延长,而且各物候的始期提前、末期推迟[22];对长江源区高山嵩草的研究发现,在区域气候变暖背景下,植物物候表现为返青期提前,开花期和枯黄期推迟,生长季长度延长[23];有人则认为返青期推迟,枯黄期被提前,生长季长度会被缩短[24]。
3.2 增温对薹草生物量的影响
本次研究表明,模拟增温对短尖薹草的根系生物量具有促进作用,与其他学者的研究结果一致[25]。植物生物量是评价生态系统的重要生理指标之一,植物分布也反映其适应环境的生长策略,温度对生物量的影响体现在多个方面,且不同植物的生物量在气候变化条件下也会有不同的变化[26,27]。温度和降水等环境条件变化可直接或间接影响植物的生产力,以致影响植物生物量的积累与分配[28];同时,物候期和生物量的积累也有一定关系,气候变暖直接影响杉木幼树的光合速率,改变植物物候,延长植物的生长季[29];通过增温处理后发现,剑叶金鸡菊(Coreopsislanceolata)的地上生物量显著增加,总生物量也增加,但增温的影响差异不显著,短期增温促进了剑叶金鸡菊的生长及生物量的积累,但增温导致土壤水分降低限制了其根系生长,使生物量更多分配给地上部分,总体有利于植物生长[30];青藏高原腹地典型高寒草甸和沼泽草甸在两种增温梯度条件下,模拟增温使沼泽草甸生物量明显地向深层土壤中转移[31]。
3.3 增温对薹草有性繁殖的影响
植物开花不仅是其生长繁殖的重要过程,还是对全球气候变暖监测最灵敏的生物学指标之一[32]。通过模拟增温的手段观察温度对于短尖薹草花期的影响,对研究洞庭湖未来气候变暖背景下植物群落的演替变化具有重要意义。本研究发现,未增温条件下的薹草无论是雄穗还是雌穗其数量均大于增温组,并且增温组的开花数量稳定期比未增温组要提前近一周,因此推测增温条件下薹草的花期将会提前,观察还发现增温组的开花率低于对照组,由此可见在未来气候变暖的情况下,薹草的花期提前且繁殖能力降低。此结论与以往的研究结果相似[33];但也有研究认为增温前后对花期并无较大影响[34,35]。因此,在模拟增温对薹草花期物候研究方面还需进行长期监测,以求得更加准确科学的结论。
