江西齐云山长苞铁杉种群特征与种间联结研究
2023-12-27汪志培黄世贵朱祥龙吴治明
汪志培,黄世贵,朱祥龙,卢 建,吴治明*
江西齐云山长苞铁杉种群特征与种间联结研究
汪志培1,黄世贵2,朱祥龙2,卢建2,吴治明2*
(1. 江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045;2. 江西齐云山国家级自然保护区管理局,江西 赣州 341300)
【目的】探讨江西齐云山国家级自然保护区长苞铁杉()种群与种间联结特征,为保护区珍稀孑遗植物的监测与保护提供科学依据。【方法】根据前期踏查的结果,在江西齐云山国家级自然保护区内选择长苞铁杉集中分布区域,建立15个40 m×40 m的长苞铁杉群落固定样地,对长苞铁杉种群和种间联结进行分析。【结果】(1)长苞铁杉个体数量总体随着龄级的增加而减少,年龄结构呈“金字塔”型。(2)长苞铁杉在Ⅱ、Ⅶ和Ⅻ龄级存在3个死亡高峰期。(3)长苞铁杉群落优势树种总体呈显著正联结,其中长苞铁杉与杉木()呈显著正相关,而与大果马蹄荷()、圆萼折柄茶()呈显著负相关。【结论】齐云山保护区长苞铁杉种群呈增长趋势,但种群生存压力较大,应提前采取相应的干预措施,以预防未来种群出现衰退。长苞铁杉群落主要物种间总体呈显著正相关,在保护过程中需重点关注与长苞铁杉呈负相关的物种,可适当进行人为干预,以防止这些物种过度增长导致长苞铁杉自然更新出现困难。
齐云山保护区;长苞铁杉;孑遗植物;种群特征;种间联结
【研究意义】长苞铁杉()属松科铁杉属,为我国特有的珍稀孑遗树种,它是大果铁杉组(Sect. Hesperopeuce)分布于中国的唯一代表种类,其叶肉薄壁组织中有石细胞,花粉有气囊[1],形态极其特殊,有别于国产各种铁杉,这对研究我国裸子植物系统发育、古生态和古气候等具有重要价值[2-4]。而种群特征和种间联结是种群生态研究的核心问题,本研究通过对珍稀孑遗植物长苞铁杉种特征和种间联结进行研究,以期揭示长苞铁杉种群在群落中的年龄结构、生存状态、与其它种群的相互作用状态,从而为长苞铁杉的保护与资源合理利用提供参考。【前人研究进展】近年来,关于长苞铁杉的研究主要集中于倒木特征[5-7]、土壤理化性质[8-9]、种苗生长特征[10-11]、林窗特征[12-13]、种群分布格局[14-15]等方面。但目前为止,关于长苞铁杉种群特征与种间联结的相关研究仍相对较少,齐云山地区在该方面的研究仍处于空白阶段。【本研究切入点】江西齐云山国家级自然保护区拥有大面积的长苞铁杉群落,群落分布面积达608.5 hm2,数量27.67万株,这在江西乃至全国都十分罕见[16]。但关于齐云山长苞铁杉的相关研究极为匮乏,这对长苞铁杉种群来说还远未达到保护与监测所需的要求,因此,亟需对齐云山保护区内分布的长苞铁杉群落开展相关研究。【拟解决的关键问题】采用典型样方法,在齐云山保护区长苞铁杉集中分布的区域建立15个40 m×40 m长苞铁杉群落固定样地。以期探明以下问题:(1)齐云山保护区长苞铁杉种群年龄结构如何?(2)齐云山保护区长苞铁杉种群生存函数特征如何?(3)齐云山保护区长苞铁杉群落主要优势树种的种间关系如何?
1 材料与方法
1.1 研究区概况
江西齐云山国家级自然保护区位于赣州市崇义县西北部(25°41′47.00″~25°54′21.00″N,113°54′37.00″~114°07′34.00″E),地处南岭山脉与罗霄山脉交汇的诸广山脉腹地,总面积17 105.0 hm2。齐云山自然保护区海拔最高为2 061.3 m,即齐云山顶峰,最低点为香炉坝,海拔仅为300.0 m。区内群山叠起,溪河纵横,属中亚热带山地湿润季风气候,四季分明、冬长夏短、春迟秋早,春夏冬3季湿润,秋季干燥,光照适宜[16]。该地区独特的地理条件,孕育了极其丰富的植被类型[17],有着江西“植物博物馆”的美誉[18]。其中长苞铁杉为齐云山最具特色的珍稀孑遗植物,其数量之多、面积之大在全国都属罕见。
1.2 样地设置与调查
2022年6—9月,经过前期的实地踏查了解植被状况,在保护区选择具有代表性的长苞铁杉集中分布地段建立15个40 m×40 m样地,总面积24 000 m2。具体样地布设参照方精云等[19]的植物群落调查方式,对样地内所有胸径(DBH)≥5.0 cm的木本植物进行每木检尺,并记录物种名称、胸径、树高、植株位置等数据。
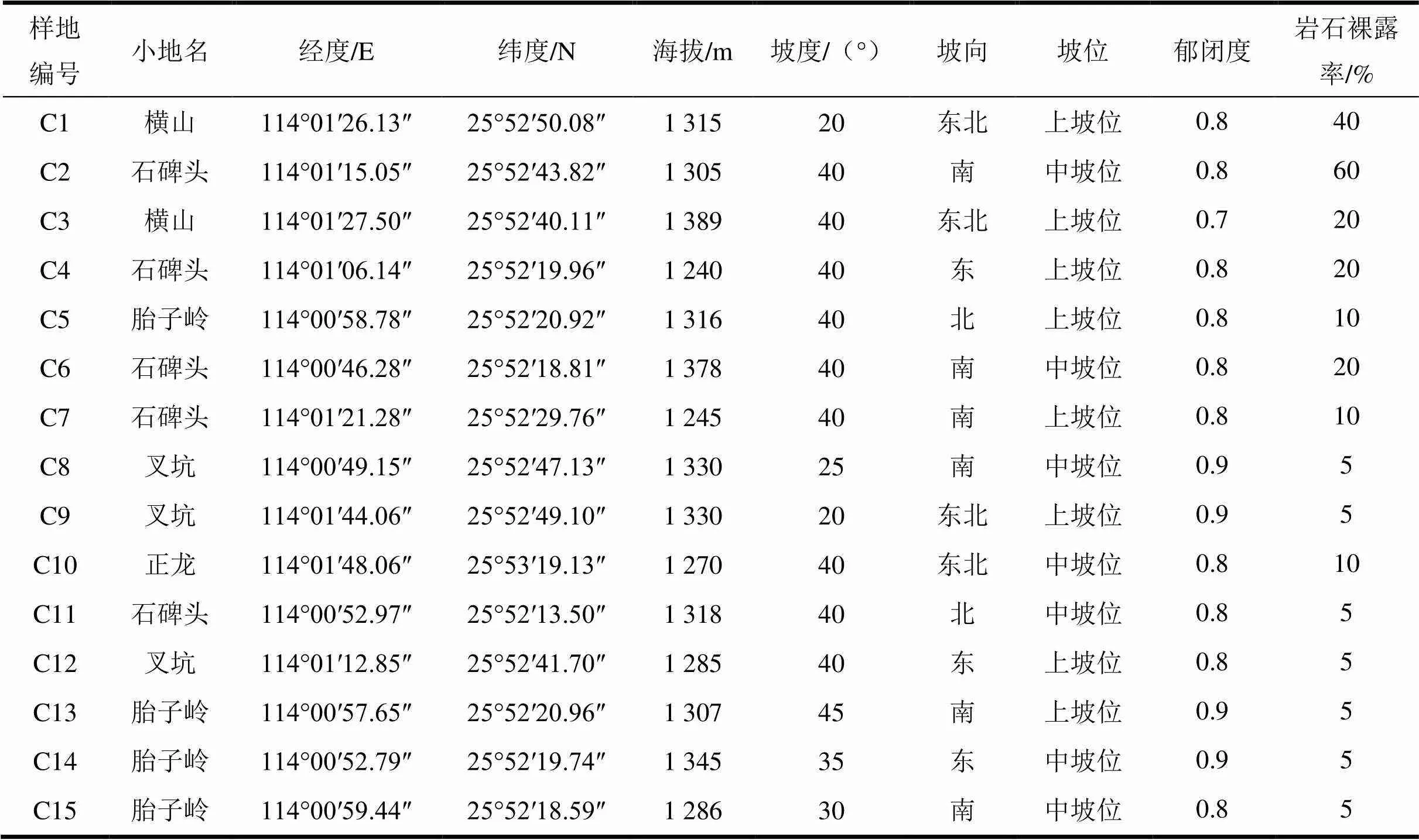
表1 长苞铁杉(Nothotsuga longibracteata)样地概况
1.3 数据处理与分析
1.3.1 年龄结构的划分 年龄结构采用时空替代法,第Ⅰ径级DBH为0.0 cm<DBH≤5.0 cm,第Ⅱ径级DBH为5.0 cm<DBH≤10.0 cm,之后每隔5.0 cm作一个径级划分,以此类推,其中Ⅰ径级对应Ⅰ龄级,Ⅱ径级对应Ⅱ龄级,并以此类推。长苞铁杉由于缺少第Ⅰ径级数据,故将长苞铁杉从第Ⅱ径级开始划分年龄结构,从Ⅱ-Ⅻ共划分11个龄级。统计在每一龄级的存活数,以龄级为纵坐标轴,株数为横坐标,绘制种群的年龄结构图。
1.3.2 静态生命表的编制 静态生命表是反映在特点时空下多个重叠的年龄动态,并不对种群整个生活史追踪,因而在编制过程中会出现死亡率为负值的情况,但这不符合编制静态生命表的假设[20]。为此,本研究采用匀滑技术对种群存活数A进行调整,得到修正后龄级内存活个体数a。
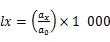
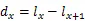
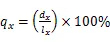
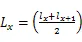
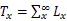
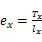

式中:a为匀滑后存活数,l为标准化存活个体数,L为平均存活数,T为总体个数,e为生命期望值,S为存活率。
1.3.3 生存分析函数计算 生存分析函数包括种群生存率函数()、累计死亡率函数()、死亡密度函数()和危险率函数()可以全面分析种群数量变化[21],计算公式如下:
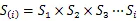
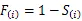

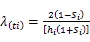
1.3.4 总体联结性计算 采用Schluter[22]的方差比率(VR)检验方法对植物群落总体联结性进行检验,并检验相关系数的显著性:



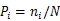

式中,为物种总数,为样方总数,T为样方中的物种数,为物种的样方数,为样方中的平均物种数。在独立性检验下,VR=1,当VR>1时,群落总体呈正联结,当VR<1时,群落总体呈负联结,由于物种间正负联结可以相互抵消,使用统计量进一步检验VR与1的偏离程度,若相关性不显著,则落入20.95()<<20.05()临界值区间内,若相关性显著,则在临界值区间外。
1.3.5 种对间联结性计算 利用卡方检验(c2)、联结系数(AC)、Jaccard指数(JI)、Ochiai指数(OI)和Dice指数(DI)计算群落内优势种的种间联结[23-24]:
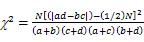
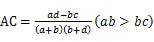
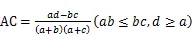

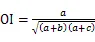
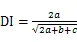
式中为样方数,通过建立种对间2×2列联表,统计的值。其中为A、B两物种在同一样方的样方数,为B物种存在而A物种不存在的样方数,为A物种存在而B物种不存在的样方数,为A、B两物种都不存在的样方数。当>时,种对间为正联结;当<时,种对间为负联结。当2<3.841时,种对间不存在联结性;当3.841<2<6.635时,种对间存在联结性;当2>6.635时,种对间为显著联结。
1.3.6 数据统计与分析 通过Excel 2016对样地数据进行预处理,采用Matlab R2021a软件对静态生命表、生存分析函数和联结性进行计算分析,并用Excel 2016和Origin 2021软件进行制图。对植株统计分析时都包括分枝在内。
2 结果与分析
2.1 长苞铁杉种群特征
2.1.1 静态生命表分析 随龄级的增加,长苞铁杉种群匀滑后存活数(a)和标准化存活个体数(l)逐渐减小,死亡率(q)反应了该种群随龄级增加的动态特征情况。死亡率在Ⅱ-Ⅶ龄级波动较大,其中Ⅱ和Ⅲ龄级的死亡率最高,说明长苞铁杉种群在幼龄期的环境生存压力较大。生命期望(e)能反应x龄级内植株个体的平均生存能力,长苞铁杉种群生命期望最高的2个龄级为Ⅴ和Ⅷ龄级,从Ⅷ-Ⅻ龄级生命期望值逐渐降低,Ⅻ龄级生命期望值最低(表2)。
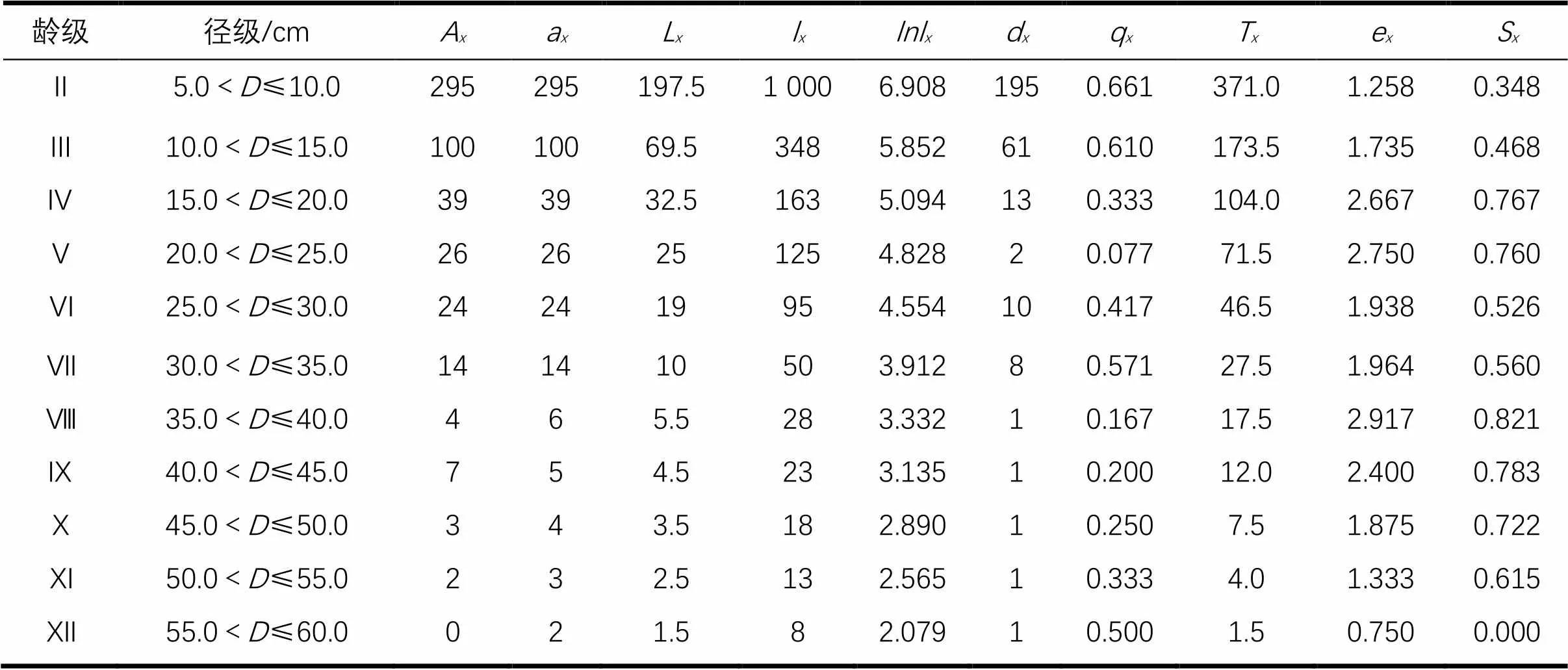
表2 长苞铁杉静态生命
:胸径(cm);:存活数;a:匀滑后存活数;L:平均存活数;l:标准化存活个体数;lnl:标准化存活个体数的自然对数;d:死亡数;q:死亡率;T:总体个数;e:生命期望值;S:存活率。
2.1.2 年龄结构特征 长苞铁杉种群共分为11个龄级,其中在第Ⅱ龄级的个体数量最多,占总个体数的57.39%,第Ⅺ龄级的个体数最少,个体数量总体随着龄级的增加而减少,年龄结构呈“金字塔”型(图1),可见齐云山长苞铁杉种群总体更新状况较为良好。

图1 长苞铁杉(Nothotsuga longibracteata)种群年龄结构
2.1.3 生存函数曲线分析 长苞铁杉的存活率随龄级的增大而降低,累计死亡率随龄级的增大而增大。在Ⅱ-Ⅳ龄级,长苞铁杉种群存活率和累计死亡率变化较大,第Ⅴ-Ⅻ龄级种群存活率和死亡率逐渐趋于稳定。第Ⅱ龄级的存活率为0.339,第Ⅳ龄级迅速降至0.088,下降幅度较大。第Ⅱ龄级的累计死亡率为0.661,到第Ⅳ龄级增加到了0.912。第Ⅴ-Ⅻ龄级长苞铁杉的存活率和累计死亡率的增减趋于平缓,大量个体由于达到一定年龄而自然死亡,种群出现衰退特征(图2),长苞铁杉种群在该阶段进入生理衰老阶段。

图2 长苞铁杉种群生存率和累计死亡率曲线
长苞铁杉种群的死亡密度在Ⅱ-Ⅷ龄级存在波动,尤其在Ⅱ-Ⅲ龄级波动较大,之后平稳降低。危险率曲线在整个年龄阶段波动都较大,表现出前期、中期和后期3个死亡高峰(图3)。可见长苞铁杉种群具有前期快速衰减、中期竞争压力大、后期到达生理年龄正常死亡的特点,后期树种到达生理年龄而正常死亡。
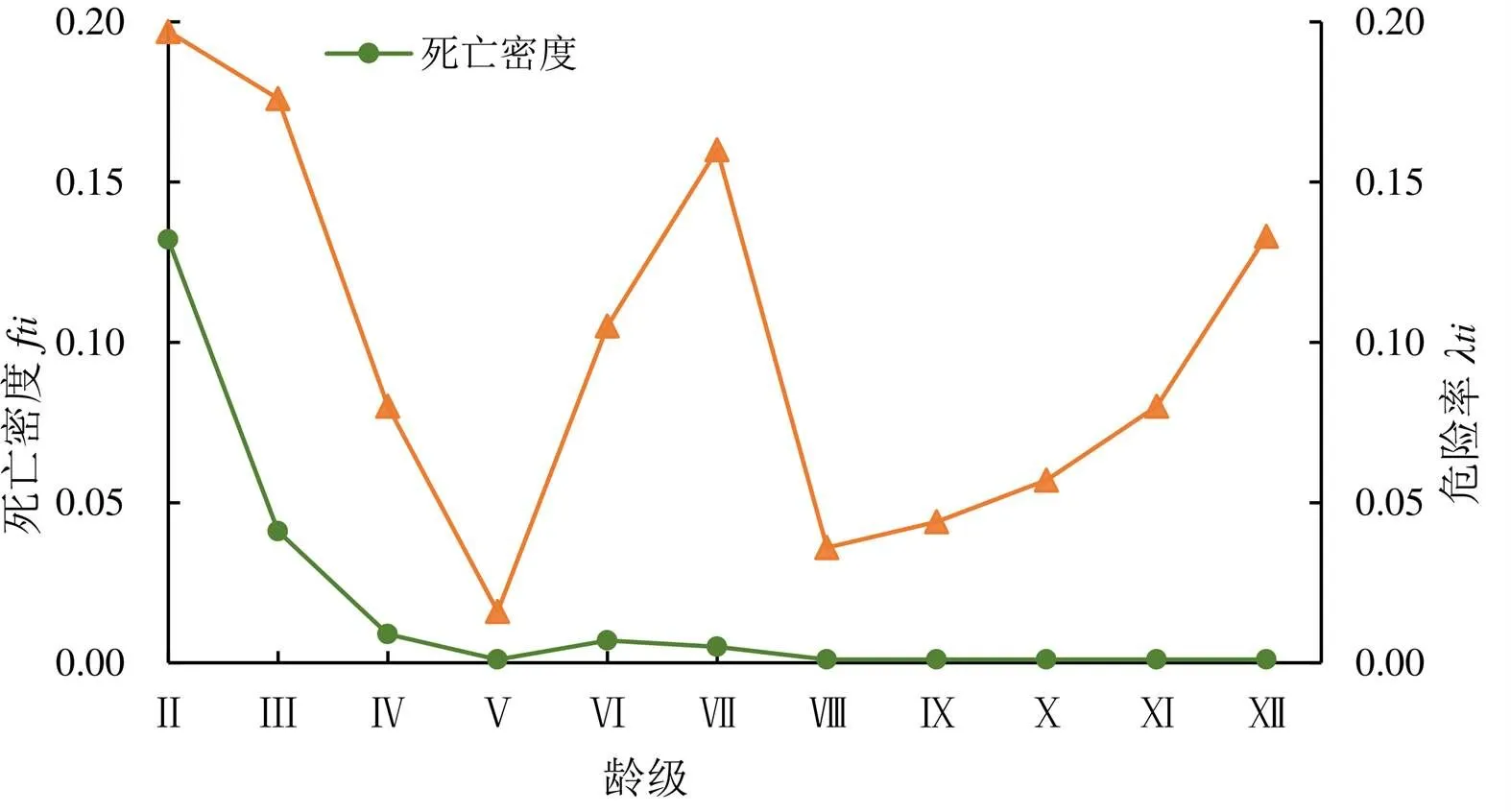
图3 长苞铁杉种群死亡密度和危险率曲线
2.2 长苞铁杉与主要树种种间关系
2.2.1 总体相关性 根据15×15的物种-样地二元数据矩阵计算长苞铁杉群落重要值前15树种的总体相关性,得总体相关性方差比率VR=1.19>1,表明15个主要物种之间总体表现为净的正相关。统计量=286.89,20.95(15)=278.21,20.05(15)=206.06,不在20.95(15)<<20.05(15)临界值区间内,表明长苞铁杉群落主要物种间呈显著正联结。

表3 长苞铁杉群落总体联结性
2.2.2 主要种对间的联结性 在105个种对中,68对不相关,呈显著相关的有37对,占总对数的35.24%,其中19对呈显著正相关,18对呈显著负相关(图4),正负联结比为1.06,表明种对间正联结相对占据优势地位,但没有占据绝对优势,存在较为激烈的种间竞争,物种更替较为频繁,群落物种组成和结构较不稳定。其中长苞铁杉-杉木种对之间呈显著正相关,长苞铁杉-大果马蹄荷、长苞铁杉-圆萼折柄茶两个种对之间呈显著负相关。
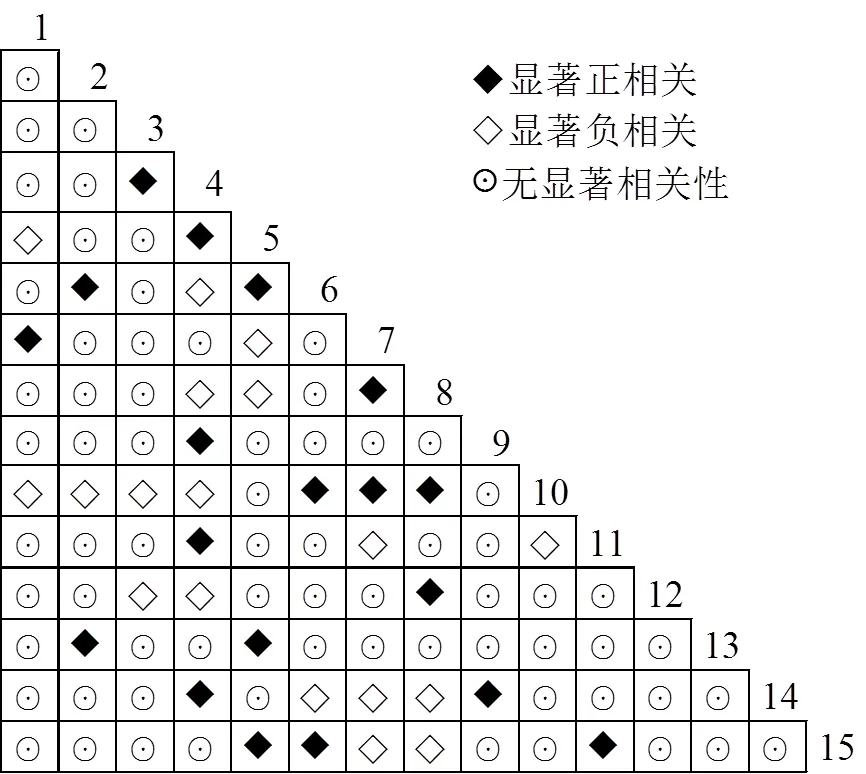
1、甜槠(Castanopsis eyrei),2、银木荷(Schima argentea),3、长苞铁杉(Nothotsuga longibracteata),4、杉木(Cunninghamia lanceolata),5、红楠(Machilus thunbergii),6、深山含笑(Michelia maudiae),7、五列木(Pentaphylax euryoides),8、福建柏(Fokienia hodginsii),9、马尾松(Pinus massoniana),10、大果马蹄荷(Exbucklandia tonkinensis),11、鹿角杜鹃(Rhododendron latoucheae),12、圆萼折柄茶(Stewartia crassifolia),13、雷公鹅耳枥(Carpinus viminea),14、石灰花楸(Sorbus folgneri),15、马银花(Rhododendron ovatum),下同。

a:联结系数AC;b:Jaccard指数(JI);c:Dice指数(DI);d:Ochiai指数(OI)。
2检验只能定性判断物种之间是否存在显著相关,不能确定种对之间相关强度的大小,还需要通过AC联结系数、JI指数、DI指数和OI指数对相关测度进行补充说明,二者是对种间相关显著性和相关强度在两个方面的不同解释。AC、JI、DI和OI的最大值分别是0.222 8、0.550 7、0.710 3和0.007 1,分别为红楠-深山含笑、甜槠-长苞铁杉、甜槠-长苞铁杉和杉木-石灰花楸。在2检验中表现为显著正相关的19个种对中,其对应的AC、JI、DI和OI值也都相对比较大,如红楠-深山含笑、五列木-福建柏等种对(图5)。
3 讨论与结论
3.1 长苞铁杉种群呈增长趋势
年龄结构可以反应出种群的发展与演变趋势[25]。本研究显示,长苞铁杉种群的年龄结构呈“金字塔”型,这说明长苞铁杉种群在齐云山的更新状况良好。这与李红等[26]对黄桑坪长苞铁杉的研究结果不一致,种群年龄结构表现为衰退型。其原因为齐云山在未成立保护区前的饥荒年代存在人为烧山的情况,而长苞铁杉具有适应火成演替相类似的机制,种群在经过林火干扰时幼苗的成活率更高[27],更有利于种群的更新,因此种群表现为增长型。
3.2 长苞铁杉种群生存压力较大
本研究发现,长苞铁杉在Ⅱ、Ⅶ和Ⅻ龄级存在3个死亡高峰期,种群表现出前期快速衰减、中期竞争压力大、后期到达生理年龄正常死亡的特点。其中,导致长苞铁杉种群在Ⅵ-Ⅷ的中龄级存在较高的死亡风险的原因,可能是由于长苞铁杉为阳性树种,喜温暖湿润气候,幼树比中龄树耐荫蔽,但中龄以上的长苞铁杉需光量较大,种群生长受环境制约较大[28-29],导致部分中龄植株在种内或种间竞争压力过大而被淘汰。Ⅶ龄级后,长苞铁杉种群累计死亡率变化趋势较为平缓,表明此时种群的竞争压力减小并已成功稳定占据一定的空间和环境资源,导致其死亡的主要因素转为自身生理衰老。
3.3 长苞铁杉与其他物种存在显著相关性
种间联结是群落中不同物种在空间分布的联结性,这种联结客观有效地阐明了物种间相互依存和相互制约的复杂关系[30-31]。长苞铁杉群落重要值前15的优势种2检验的105个种对中,不显著相关的有68对,呈显著相关的种对有37对,其中19对呈显著正相关,18对呈显著负相关,说明长苞铁杉群落未形成较为稳定的种间关系,群落处于演替的前中期阶段。
相关研究表明,在生长环境及竞争一致的条件下,具有相同或相似生态习性的物种往往共同出现在同一群落中,其种间联结性一般表现为显著的正相关,种对常表现为互利、互补的关系[32-33];而不同生态习性的物种具有不同的生态位,其种间联结性一般表现显著负相关,种对常表现为排斥、竞争的关系[34-35]。本研究中长苞铁杉的种间相关性也验证了这一规律。例如:联结系数显示长苞铁杉与杉木呈显著正相关,原因是二者都为喜光针叶树种,针叶树的林下透光性较好[36],有利于相互之间幼苗的生长,所以表现为互利、互补的关系;而联结系数显示长苞铁杉与大果马蹄荷之间呈显著负相关,原因是二者生长需光性不同,长苞铁杉为喜光阳性树种[37],大果马蹄荷为耐荫树种[38],并且枝叶浓密林下光照度极低,这可能会造成长苞铁杉种子萌发和幼苗生长没有合适的环境条件[39],因此表现出排斥、竞争的关系。
虽然目前群落处于演替的前中期,长苞铁杉种群更新状况良好,短时间内种群仍会呈增长趋势。但长苞铁杉属于阳性树种,随着演替的进行,大果马蹄荷和圆萼折柄茶等呈显著负相关耐荫树种会进一步压缩长苞铁杉的生存空间,再加上种群中龄期出现死亡率高的特点,这会使得种苗难以得到足够数量的更新,并且种子传播存在范围小(多在15.0~50.0 m),幼苗分布呈现一定的聚集性的情况[40]。以上情况表明,虽然目前长苞铁杉在齐云山更新状况良好,但未来种群生存形式不容乐观,应当采取相应的干预性保护措施控制群落中与长苞铁杉种群呈负相关的树种数量,为长苞铁杉幼苗创造阳光充足的生长环境,以促种群的自然更新,预防未来种群出现衰退的情况。
综上所述,齐云山长苞铁杉种群呈增长趋势,短期内种群数量仍会增加,但种群生存压力较大,从长远来看生存形式不容乐观,应当提前采取相应的干预性保护措施,以预防未来种群出现衰退。长苞铁杉群落处于演替的前中期,在长苞铁杉就地保护过程中,需关注与长苞铁杉呈负相关的物种,特别是与长苞铁杉呈显著负相关的大果马蹄荷、圆萼折柄茶等物种。迁地保护或生境修复时,可寻找与长苞铁杉正相关性较强的物种,如杉木等,与之形成功能搭配,为长苞铁杉创造良好的生存环境。
[1] 中国科学院中国植物志委员会. 中国植物志(第七卷)[M]. 北京: 科学出版社, 1978.
[2] 中国科学院植物研究所. 中国珍稀濒危植物[M]. 上海: 上海教育出版社, 1989.
[3] 傅立国. 中国植物红皮书: 稀有濒危植物[M]. 北京: 科学出版社, 1991.
[4] 林金星, 胡玉熹, 王献溥, 等. 中国特有植物长苞铁衫的生物学特性及其保护[J]. 生物多样性, 1995, 3(3): 147-152.
[5] 穆振北, 潘辉, 温鑫鸿, 等. 腐烂等级和径级对天宝岩长苞铁杉林木质残体理化性质的影响[J]. 生态学杂志, 2022, 41(2): 246-255.
[6] 肖石红, 何东进, 刘进山, 等. 天宝岩长苞铁杉倒木微生物生物量和可溶性有机碳氮的变化[J]. 生态与农村环境学报, 2017, 33(11): 983-991.
[7] 何东进, 游惠明, 肖石红, 等. 天宝岩长苞铁杉林倒木接触处土壤酶活性变化及其环境效应[J]. 生态学报, 2017, 37(1): 118-126.
[8] 黄雄俊, 刘君成, 温鑫鸿, 等. 福建天宝岩不同长苞铁杉林土壤微生物生物量碳含量的比较及其与土壤含水量的关系[J]. 植物资源与环境学报, 2022, 31(5): 1-8.
[9] 游惠明, 何东进, 蔡昌棠, 等. 天宝岩长苞铁杉林倒木对土壤肥力质量的影响评价[J]. 应用与环境生物学报, 2013, 19(1): 168-174.
[10] 游秀华, 白静, 刘君成, 等. 倒木基质对长苞铁杉种苗生长及生理特性的影响[J]. 中国农学通报, 2020, 36(31): 20-27.
[11] 朱小龙, 冯大兰. 长苞铁杉天然更新研究Ⅱ.不同群落的幼苗建立及其环境影响[J]. 福建林学院学报, 2011, 31(4): 315-319.
[12] 李苏闽, 何东进, 覃德华, 等. 福建天宝岩国家级自然保护区长苞铁杉林林窗的物种构成和边缘效应分析[J]. 植物资源与环境学报, 2015, 24(4): 89-96.
[13] 李苏闽, 游巍斌, 肖石红, 等. 天宝岩长苞铁杉林林窗的微环境特征[J]. 森林与环境学报, 2015, 35(4): 343-350.
[14] 谭雪, 张林, 张爱平, 等. 孑遗植物长苞铁杉()分布格局对未来气候变化的响应[J]. 生态学报, 2018, 38(24): 8934-8945.
[15] 严绍裕. 天宝岩自然保护区长苞铁杉不同群落结构与空间格局比较[J]. 西南林业大学学报, 2013, 33(4): 44-48.
[16] 刘小明, 郭英荣, 刘仁林. 江西齐云山自然保护区综合科学考察集[M]. 北京: 中国林业出版社, 2010.
[17] 宋述望, 罗会潭, 黄萌, 等. 江西齐云山自然保护区植被类型与森林资源特征[J]. 南方林业科学, 2021, 49(5): 34-38.
[18] 黄声亮, 陈齐通, 龚超, 等. 江西齐云山植物群落多样性及蓄积量对海拔梯度的响应[J]. 生物灾害科学, 2023, 46(1): 80-85.
[19] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533-548.
[20] 江洪. 云杉种群生态学[M]. 北京: 中国林业出版社, 1992.
[21] 杨凤翔, 王顺庆, 徐海根, 等. 生存分析理论及其在研究生命表中的应用[J]. 生态学报, 1991, 11(2): 153-158.
[22] SCHLUTER D. A variance test for detecting species associations, with some example applications[J]. Ecology, 1984, 65(3): 998-1005.
[23] 江常春, 梁希妮, 田磊, 等. 浙江省常绿阔叶林优势树种种间联结分析[J]. 浙江师范大学学报(自然科学版), 2022, 45(4): 437-445.
[24] 张零念, 朱贵青, 杨宽, 等. 滇中云南杨梅灌丛主要木本植物生态位与种间联结[J]. 植物生态学报, 2022, 46(11): 1400-1410.
[25] 焦德志, 王昱深, 杨允菲. 扎龙湿地不同生境芦苇种群根茎构件的年龄结构[J]. 生态学报, 2020, 40(7): 2186-2193.
[26] 李红, 杨宁. 湖南省绥宁县黄桑坪自然保护区珍稀濒危植物长苞铁杉自然种群年龄结构及生态对策[J]. 湖南生态科学学报, 2014, 1(3): 12-16.
[27] 王建林. 林火干扰对长苞铁杉更新的影响[J]. 华东森林经理, 2011, 25(2): 18-21.
[28] 邱迎君, 李作洲, 黄宏文. 濒危植物长苞铁杉的年龄结构与空间格局的研究[J]. 武汉植物学研究, 2008, 26(5): 495-500.
[29] 吴承祯, 洪伟, 吴继林, 等. 珍稀濒危植物长苞铁杉优势度增长规律的研究[J]. 林业科学, 2004, 40(2): 189-192.
[30] 王国庆, 杜广明, 聂莹莹, 等. 围栏封育对呼伦贝尔草甸草原群落种间联结的影响[J]. 中国草地学报, 2017, 39(4): 72-80.
[31] 黄石德, 聂森, 肖祥希, 等. 武夷山米槠群落优势种群生态位与种间联结[J]. 植物科学学报, 2023, 41(3): 291-300.
[32] 农友, 郑路, 贾宏炎, 等. 广西大青山南亚热带森林植物群落的种间联结性[J]. 广西植物, 2016, 36(7): 848-858.
[33]薛卫星, 李春辉, 艾训儒, 等. 鹅掌楸天然林优势树种生态位与种间联结性[J]. 森林与环境学报, 2023, 43(1): 26-34.
[34] 徐满厚, 刘敏, 翟大彤, 等. 植物种间联结研究内容与方法评述[J]. 生态学报, 2016, 36(24): 8224-8233.
[35] 刘益鹏, 叶兴状, 叶利奇, 等. 观光木群落优势树种生态位和种间联结[J]. 应用生态学报, 2022, 33(10): 2670-2678.
[36] 王俊男, 于洋洋, 林柳兴, 等. 杉木林林内光环境对楠木生长的影响[J]. 森林与环境学报, 2018, 38(4): 425-430.
[37] 邱迎君, 易官美, 宁祖林, 等. 濒危植物长苞铁杉的地理分布和资源现状及致危因素分析[J]. 植物资源与环境学报, 2011, 20(1): 53-59.
[38] 夏江林, 彭珍宝, 范水平, 等. 36个稀有树种的耐性分析[J]. 植物资源与环境学报, 2005, 14(3): 47-52.
[39] 罗敏贤, 林碧华, 陈绪辉, 等. 福建龟山伞花木所在群落乔灌层优势种的种间联结性[J]. 植物资源与环境学报, 2022, 31(6): 63-72.
[40] 钱莲文, 郭建宏, 吴承祯, 等. 长苞铁杉林林隙物种更新动态的初步研究[J]. 江西农业大学学报, 2005, 27(5): 719-722.
Population Characteristics and Interspecific Association ofin Qiyunshan Mountain of Jiangxi Province
WANG Zhipei1, HUANG Shigui2, ZHU Xianglong2, LU Jian2, WU Zhiming2*
(1. Jiangxi Provincial Key Laboratory for Bamboo Germplasm Resources and Utilization, Jiangxi Agricultural University, Nanchang 330045, China; 2. Administration of Qiyunshan National Nature Reserve, Ganzhou, Jiangxi 341300, China)
This study was conducted to explore the population characteristics and interspecific association ofin Qiyunshan National Nature Reserve of Jiangxi Province, so as to provide scientific basis for the monitoring and protection of rare and relict plants there.According to the results of preliminary investigation, concentrated distribution areas ofwas selected in Qiyunshan National Nature Reserve of Jiangxi Province, and 15 fixed plots of 40 m×40 mcommunity were established to analyze the population and interspecific association of.(1) The number ofindividuals decreased with the increase of age class, and the age structure was a kind of ‘pyramid’ type; (2) There were three death peaks ofin II, VII and XII age classes; (3) The dominant tree species ofcommunity were significantly positively correlated.was significantly positively correlated with, and significantly negatively correlated withand.Thepopulation in the protection area of Qiyunshan Mountain showed an increasing trend, but the population survival pressure is large. Corresponding intervention measures should be taken in advance to prevent the future population decline. There is a significant positive correlation between the main species ofcommunity. In the process of protection, it is necessary to pay attention to the species negatively correlated with, and appropriate human intervention can be carried out to prevent the excessive growth of these species from causing difficulties in the natural regeneration of.
Qiyunshan National Nature Reserve;;relict plant; population characteristics; interspecific association
10.3969/j.issn.2095-3704.2023.04.75
S719.17
A
2095-3704(2023)04-0500-10
2023-06-12
2023-08-25
中央财政林业国家级自然保护区补助资金项目(2020)
汪志培(1996—),男,硕士,主要从事生物多样性保护研究,863103096@qq.com;*通信作者:吴治明,高级工程师,qyszrbhq@163.com。
汪志培, 黄世贵,朱祥龙, 等. 江西齐云山长苞铁杉种群特征与种间联结研究[J]. 生物灾害科学, 2023, 46(4): 500-509.
