桉树、相思、湿地松生物量估算系数差异特征及其影响因素
2023-12-27刘晓华杜阿朋许宇星王志超黄润霞竹万宽
刘晓华,杜阿朋,许宇星,王志超,黄润霞,竹万宽*
桉树、相思、湿地松生物量估算系数差异特征及其影响因素
刘晓华1,杜阿朋2,许宇星2,王志超2,黄润霞2,竹万宽2*
(1. 广东湛江红树林国家级自然保护区管理局,广东 湛江 524088;2. 中国林业科学研究院速生树木研究所,广东 湛江 524022)
对已出版或发表的我国不同地点的桉树(主要包括尾巨桉、巨尾桉、尾叶桉和尾细桉等)、相思(主要包括马占相思、黑木相思、大叶相思、台湾相思和肯氏相思等)和湿地松生物量数据进行收集、整理和筛选,分析不同人工林生物量3个常见估算系数(生物量转换与扩展系数BCEF、生物量扩展系数BEF和根茎比)的差异特征以及与林分结构特征(林龄、胸径、树高、林分密度、林分蓄积量)、气候因子(年均气温、年均降水量)和地形因子(海拔)的关系。结果表明:桉树、相思、湿地松人工林的BCEF平均值分别为0.66 Mg·m−3、0.98 Mg·m−3、0.69 Mg·m−3,BCEF平均值分别为1.26、1.61、1.66,平均值分别为0.19、0.23、0.22。桉树林生物量估算系数的主要影响因子为树高和林分密度,相思林生物量估算系数主要受胸径和蓄积量影响。湿地松林生物量估算系数受林分结构特征影响不大,主要受到气候因子(年均温度、年均降水量)和地形因子(海拔)影响。因此,在估算区域尺度人工林生物量时应考虑林分结构特征及气候和地形因子引起的生物量估算系数差异。
桉树;相思;湿地松;生物量估算系数;影响因子
森林生态系统是陆地生态系统的主体,对陆地植物总固碳量和碳储量的贡献率分别达82.5%和33% ~ 46%,在调节全球碳平衡及减缓温室气体浓度上升中具有不可替代的作用[1-2]。森林生物量是评估森林碳收支过程的重要指标,提高其估算精度对于量化森林生态系统固碳效率、森林可持续发展及评估气候变化等至关重要[3-4]。政府间气候变化委员会(IPCC)发布的《土地利用、土地利用变化和林业的优良做法指南》和《2006 IPCC国家温室气体清单编制指南》均建议使用生物量估算参数法评估区域森林生物量[5]。常用的森林生物量估算系数包括生物量转换与扩展系数(BCEF)、生物量扩展系数(BEF)、根茎比()和基本木材密度(D)等。相关研究表明,生物量估算系数随森林类型、林龄、林分密度、立地条件等变化表现出很大差异[5-7]。因此,研究生物量估算系数与影响因子间的关系,对提高森林生物量估算精度意义重大。
桉树(spp.)、相思(spp.)、湿地松()是我国南方常见的造林树种,本研究利用收集整理的桉树、相思、湿地松林生物量数据,分析BCEF、BEF和估算系数的差异特征及主要影响因子,以期为提高区域尺度森林碳收支评估准确度提供科学依据。
1 研究方法
1.1 数据来源及筛选
收集整理CNKI中国知网数据库和万方数据库已发表或出版的144篇文献(其中桉树52篇,相思60篇,湿地松32篇),共543(桉树248条,相思182条,湿地松113条)条生物量实测数据,数据指标包括:研究区(试验地),经纬度(°),海拔(m),年均气温(℃),年均降水量(mm),林龄(年),平均胸径(cm),平均树高(m),林分密度(株·hm−2),林分蓄积量(m3·hm−2),树干、树皮、树枝、树叶、树根生物量(Mg·hm−2)和乔木地上部分、地下部分生物量(Mg·hm−2)等。数据筛选原则按照竹万宽等[5,8]文中所述。数据来源见表1。
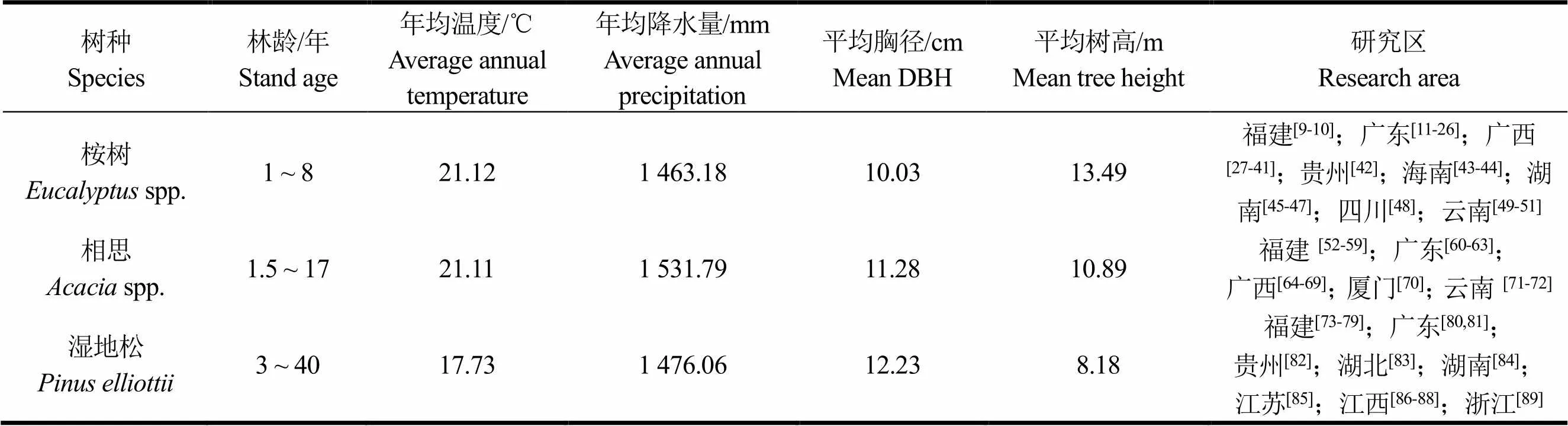
表1 数据来源
1.2 生物量估算公式
IPCC提倡的森林生物量计算基于生物量和蓄积量的转换系数,计算公式[90]为:
BCEF=/(1)
BEF=/(2)
=/(3)
式中为地上部分生物量(Mg·hm−2),为树干生物量(Mg·hm−2),为地下部分生物量(Mg·hm−2),为蓄积量(m3·hm−2)。
1.3 数据分析
采用单因素方差分析法分析桉树、相思、湿地松生物量估算系数的差异特征,Pearson相关性分析法分析生物量估算系数与影响因子的关系特征,回归分析建立生物量估算系数与影响因子的关系模型。
2 结果与分析
2.1 生物量估算系数差异特征
由图1可知,相思人工林生物量估算系数BCEF平均值(0.98±0.09 Mg·m−3)显著高于桉树(0.66±0.01 Mg·m−3)和湿地松(0.69±0.15 Mg·m−3),BEF表现为相思(1.61±0.06)和湿地松(1.66±0.11)显著高于桉树(1.25±0.02),表现为三个树种之间无显著差异。
2.2 生物量估算系数与影响因子的相关性分析
由图2可知,桉树生物量估算系数BCEF与树高和蓄积量呈极显著负相关,BEF与树高、林龄、胸径、蓄积量呈极显著负相关,仅与蓄积量呈极显著负相关;相思生物量估算系数BCEF与胸径、树高、蓄积量呈极显著负相关,BEF与胸径、树高、蓄积量、林龄呈极显著负相关,仅与蓄积量、树高呈极显著负相关;湿地松生物量估算系数BCEF、BEF、与林分结构特征因子均不存在显著相关性。

图中不同小写字母表示差异显著(P<0.05)。Different lowercase letters on the bars indicate significant differences among treatments (P<0.05).

Age 林龄;DBH 平均胸径;H 平均树高;D 林分密度;V 林分蓄积量。Age: stand age; DBH: mean diameter at breast height; H: mean tree height; D: stand density; V: stand volume.
由图3可知,桉树生物量估算系数BCEF、BEF与海拔、降水量及气温的相关性均不显著,与海拔呈极显著正相关;相思生物量估算系数BCEF、与气温分别呈显著、极显著负相关;湿地松生物量估算系数BCEF与气温呈显著正相关,BEF与海拔、气温呈显著正、负相关,仅与降水量呈显著正相关。
由图4可知,桉树生物量估算系数BCEF、BEF与树高的关系分别符合线性和二次多项式模型,BCEF随树高增大而减小,说明随着桉树的树高生长,地上部分中的树干所占生物量比重逐渐增大。桉树BEF随树高增大先出现减小趋势,在树高达到16.7 m后出现增大趋势,说明在桉树的生长前期,树干生物量积累速率较树枝、树叶快,而生长后期树干生物量积累速率相比于生长前期较树枝、树叶有所减小。与林分密度符合对数函数模型,随林分密度增大而减小,说明随着桉树的生长,根系所占生物量比例一直在减小,这主要是由于地上部分生物量积累速率较地下部分更快。
相思生物量估算系数BCEF、BEF与胸径的关系分别符合对数和二次多项式模型,均随胸径增大而减小,表明随着相思的径向生长,树干部分所占生物量比例逐渐增大。与蓄积量的关系符合幂函数模型,随蓄积量的增大急剧减小后趋于平缓,表明相思生长后期地上与地下部分生物量所占比例基本维持不变。
湿地松生物量估算系数与林分结构特征因子的回归关系不明显。

ASL 海拔;T 年均气温;P 年均降水量。ASL: altitude; T: average annual temperature; P: average annual precipitation.
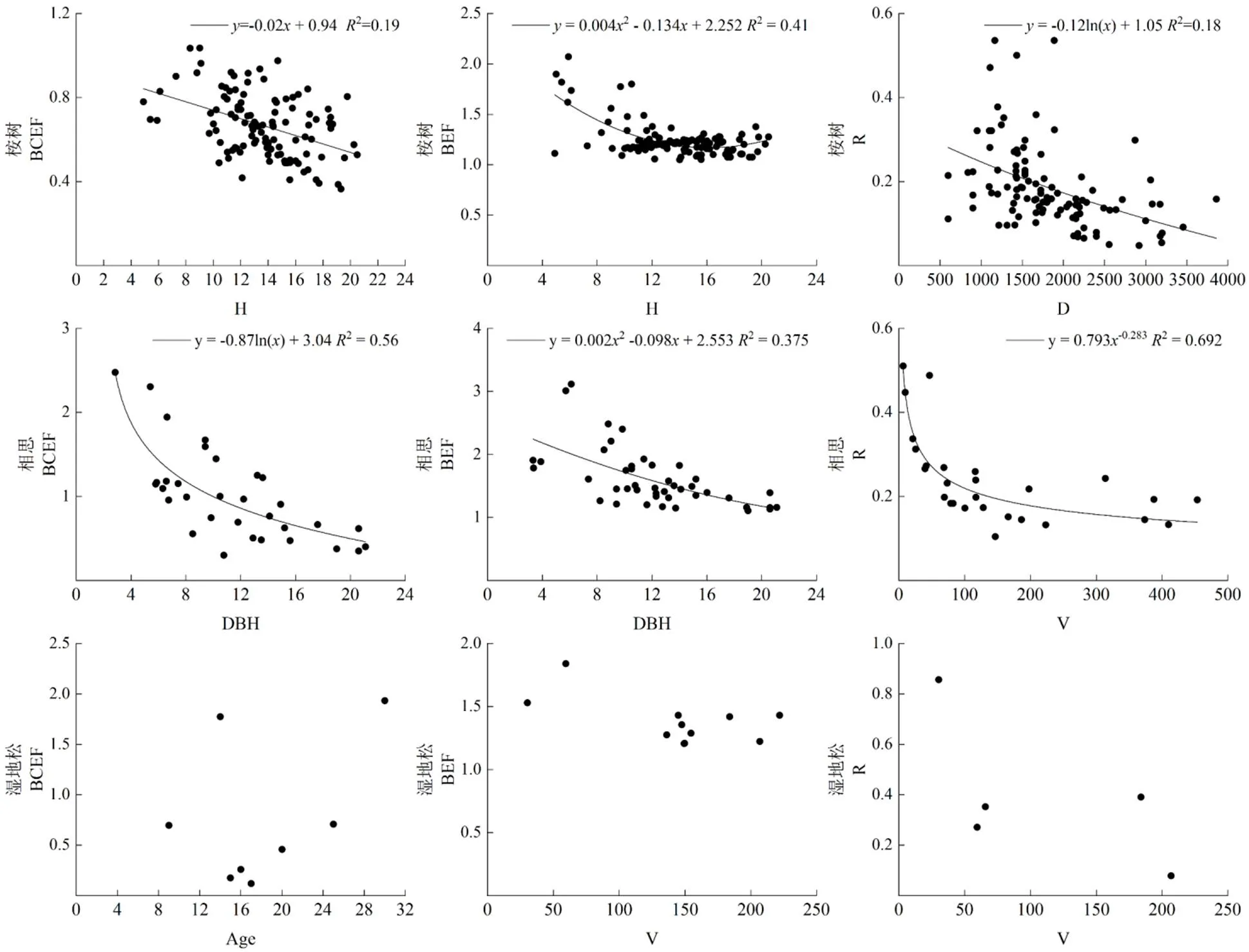
Age 林龄;DBH 平均胸径;H 平均树高;D 林分密度;V 林分蓄积量。Age: stand age; DBH: mean diameter at breast height; H: mean tree height; D: stand density; V: stand volume.
由图5可知,桉树生物量估算系数与海拔的关系符合线性模型,随海拔升高而增大。相思生物量估算系数BCEF、BEF、分别与气温、海拔、气温呈指数、幂函数、幂函数模型,均随影响因子增大而减小。湿地松生物量估算系数BCEF、BEF、分别与气温、海拔、降水量符合线性、线性、指数函数模型,均随影响因子增大而增大。
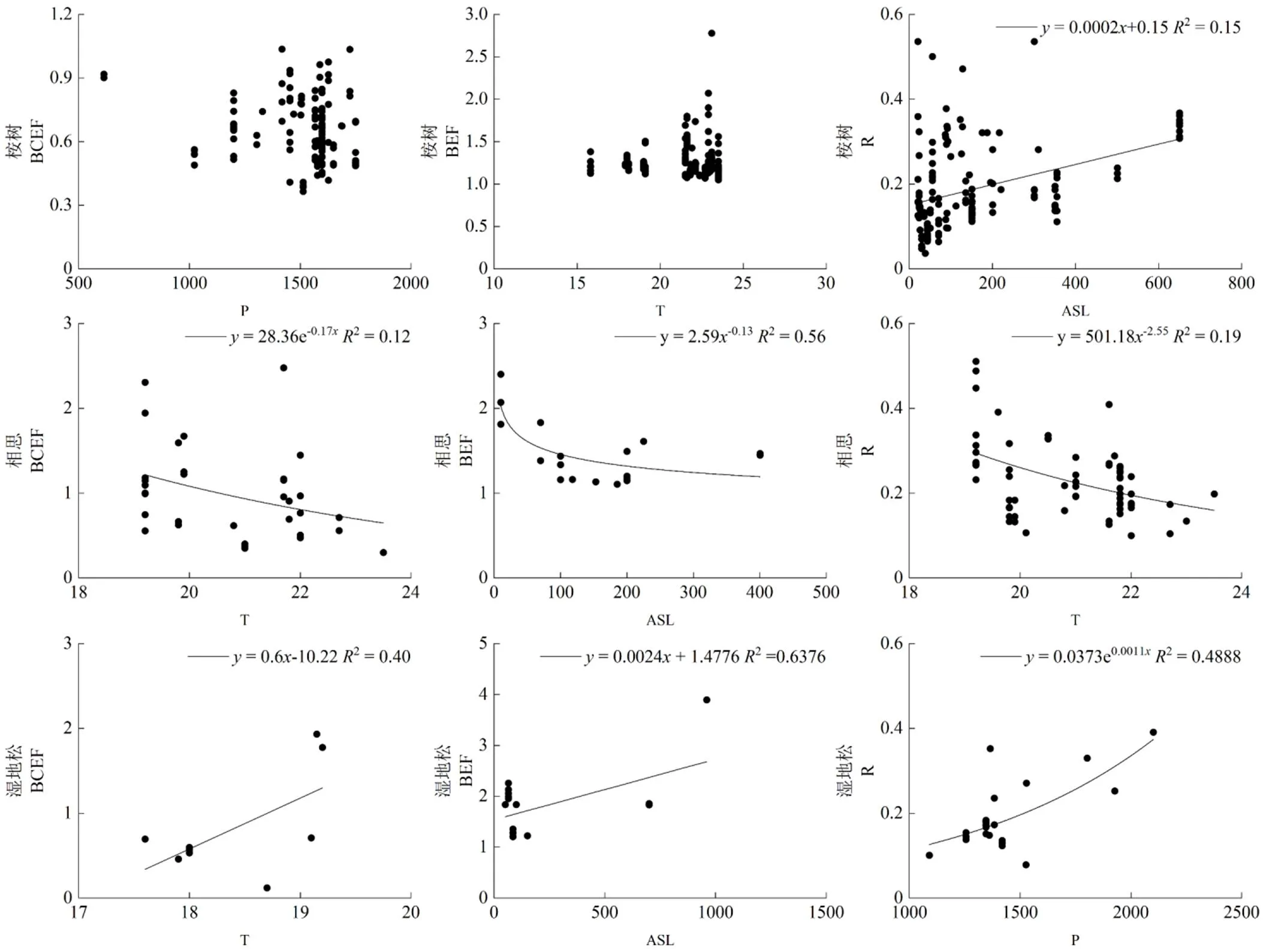
ASL 海拔;T 年均气温;P 年均降水量。ASL: altitude; T: average annual temperature; P: average annual precipitation.
3 讨论与结论
BCEF表征林分生物量和蓄积量之间的关系,本研究中,桉树、相思、湿地松人工林的BCEF平均值分别为0.66、0.98、0.69 Mg·m−3,其中,桉树和湿地松BCEF与亚热带阔叶林接近(0.66 Mg·m−3),而相思BCEF则远高于桉树和湿地松,可能是由于相思树干和树枝生物量所占比例较大[8],地上部分生物量相对较大。
BEF和表征林分器官生物量的分配状况。本研究中,桉树BEF平均值(1.26)低于温带阔叶林(1.4)和部分亚热带常绿阔叶林(1.39),相思(1.61)和湿地松(1.66)则高于温带和部分亚热带常绿阔叶林,但三者都低于热带阔叶林(3.4)[91]。桉树属于速生树种,自然整枝能力较强,且冠幅较小,树枝、树叶生物量占比小于相思、湿地松及其他常绿阔叶树种,且树干生物量占比高于其他树种[5],因此桉树BEF平均值较小。桉树平均值(0.19)低于相思(0.23)和湿地松(0.22),且三者均低于IPCC报告的部分阔叶林(0.24 ~ 0.46),可能是因为桉树、相思、湿地松适生区降水量较大,土壤水分条件较充足,减弱了植株生物量向根系的分配。
林分结构特征对生物量估算系数的影响在不同树种之间存在较大差异。本研究中,桉树生物量估算系数的主要影响因子为树高和林分密度,相思生物量估算系数则主要受到胸径和蓄积量的影响,湿地松生物量估算系数与林分结构特征的关系特征不明显,但却受到气候因子(温度和降水量)和地形因子(海拔)较大影响。因此,在对不同树种生物量估算时,应考虑林分结构特征(树高、胸径、林分密度、蓄积量等)、气候因子(温度、降水量)及地形因子(海拔)等对生物量估算系数的影响。
[1] PAN Y, BIRDSEY R A, FANG J, et al. A large and persistent carbon sink in the world’s forests[J]. Science, 2011, 333(6045): 988-993.
[2] 刘魏魏,王效科,逯非,等.全球森林生态系统碳储量、固碳能力估算及其区域特征[J].应用生态学报,2015, 26(9):2881-2890.
[3] TEMESGEN H, AFFLECK D, POUDEL K, et al. A review of the challenges and opportunities in estimating above ground forest biomass using tree-level models[J]. Scandinavian Journal of Forest Research, 2015, 30(4): 1-10.
[4] TIMMONS D S, BUCHHOLZ T, VEENEMAN C H. Forest biomass energy: assessing atmospheric carbon impacts by discounting future carbon flows[J]. Gcb Bioenergy, 2015, 8(3):631-643.
[5] 竹万宽,许宇星,王志超,等.中国桉树人工林生物量估算系数及影响要素[J].林业科学,2020,56(5):1-11.
[6] 罗云建,张小全,王效科,等.森林生物量的估算方法及其研究进展[J].林业科学,2009,45(8):129-134.
[7] 袁锋,陈灿,夏心慧,等.马尾松林生物量估算系数及其影响因子[J].生态学杂志,2021,40(6):1557-1566.
[8] 竹万宽,揭凡,杜阿朋.区域尺度相思、湿地松生物量相对生长方程及分配格局[J].桉树科技,2018,35(4):7-13.
[9] 张琼.桉树工业原料林生态系统生物量和碳贮量初步研究[D].福州:福建农林大学,2005.
[10] 郭明丽.幼林桉树生长状况与抚育方式不同的试验研究[J].华东森林经理,2006,20(3):24-26.
[11] 韩斐扬,周群英,陈少雄,等.2种桉树不同林龄生物量与能量的研究[J].林业科学研究,2010,23(5):690-696.
[12] 韩斐扬,周群英,陈少雄.6年生11种桉树无性系生物量与炭化研究[J].桉树科技,2012,29(3):1-8.
[13] 周群英,陈少雄,韩斐扬,等.短周期尾巨桉能源林生物量与能量特征研究[J].热带亚热带植物学报,2013, 21(1):45-51.
[14] 王志超.不同整地措施对桉树幼林生长及林地环境变化影响[D].北京:中国林业科学研究院,2014.
[15] 张利丽,王志超,陈少雄,等.不同林龄尾巨桉人工林的生物量分配格局[J].西北农林科技大学学报(自然科学版),2017,45(6):61-68.
[16] 周建辉,郑磊,张婧,等.不同林龄尾巨桉林木碳贮量研究[J].桉树科技,2013,30(4):11-14.
[17] 潘永言.桉树生理特征与生物量的研究[J].桉树科技,1990(增刊):24-36.
[18] 肖文光,陈悦.桉树与厚荚相思混交林生物量及对土壤影响研究[J].桉树科技,1999 (1):20-30.
[19] 谭绍满,宋智毅.雷州林业局柠檬桉林分地力和生产力关系的研究[J].桉树科技,1984(2):1-11.
[20] 许宇星,陈少雄.超高密度尾巨桉能源林追肥效应[J].林业工程学报,2013,27(6):35-38.
[21] 余雪标,徐大平,龙腾,等.连栽桉树人工林生物量及生产力结构的研究[J].华南热带农业大学学报,1999, 5(2):11-14.
[22] 郭乐东,周毅,钟锡均,等.西江流域桉树生态系统碳贮量与碳汇功能经济价值评价[J].广东林业科技,2009,25(6):8-13.
[23] 邓瑞文,陈天杏,冯咏梅.热带人工林的光能利用与生产量的研究[J].生态学报,1985,5(3):39-48.
[24] 李志辉,陈少雄,黄丽群,等.林分密度对邓恩桉生物产量及生产力的影响[J].中南林业科技大学学报,2007, 27(5):1-5.
[25] 梁坤南,周文龙,李贻铨.尾叶桉实生林施肥6a试验研究[J].林业科学研究,2002,15(6):644-653.
[26] 郑海水,曾杰,周再知,等.胶园防护林更新树种的选择[J].广东林业科技,1997,13(1):15-21.
[27] 吴立素.桉树退耕还林模式生物量、碳储量及营养元素研究[D].南宁:广西大学,2016.
[28] 骆栋卿.高峰林场巨尾桉密植林生物量生产力及土壤水文功能研究[D].南宁:广西大学,2016.
[29] 叶绍明,龙滔,蓝金宣,等.尾叶桉与马占相思人工复层林碳储量及分布特征研究[J].江西农业大学学报,2010, 32(4):735-742.
[30] 李春宁,韦建宏,付军,等.2种更新方式尾巨桉中龄林地上部分生物生产力比较分析[J].农业研究与应用,2017(6):17-20.
[31] 陈婷,温远光,孙永萍,等.连栽桉树人工林生物量和生产力的初步研究[J].广西林业科学,2005,34(1):8-12.
[32] 付威波,彭晚霞,宋同清,等.不同林龄尾巨桉人工林的生物量及其分配特征[J].生态学报,2014,34(18):5234-5241.
[33] 李况.不同年龄桉树人工林生态系统碳氮储量分配特征[D].南宁:广西大学,2013.
[34] 杨卫星,何斌,卢开成,等.桂西南连续年龄序列尾巨桉人工林的生物生产力[J].农业研究与应用,2016(3):6-11.
[35] 刘正富.巨尾桉人工林生长、生物量及植物多样性的动态变化[D].南宁:广西大学,2012.
[36] 陶玉华,隆卫革,马麟英,等.柳州市马尾松、杉木、桉树人工林碳储量及其分配[J].广东农业科学,2011, 38(22):42-45.
[37] 卢文科.林下植被去除对桉树人工林碳收支的影响[D]. 南宁:广西大学,2017.
[38] 韦宇静,梁士楚,黄雅丽,等.巨尾桉与几种阔叶树和针叶树碳储量的比较研究[J].广西科学院学报,2014,30(4): 229-232.
[39] 赵瑞,孙保平,于明含,等.广西壮族自治区平果县退耕还林植被碳储量特征[J].水土保持通报,2015,35(3): 350-353.
[40] 温远光,梁宏温,招礼军,等.尾叶桉人工林生物量和生产力的研究[J].热带亚热带植物学报,2000,8(2):123-127.
[41] 谢伟东,温远光,周敏毅,等.新栽培区尾叶桉人工林的生物量和生产力[J].中南林业科技大学学报,2007, 27(5):13-18.
[42] 甘桂春,王伟,王晓宁,等.贵州南部桉树人工林生物量及碳储量研究[J].林业调查规划,2016,41(2):34-37.
[43] 向仰州.海南桉树人工林生态系统生物量和碳储量时空格局[D].北京:中国林业科学研究院,2012.
[44] 李东海,杨小波,吴庆书.不同桉树人工林Cu元素积累和循环的比较研究[J].水土保持学报,2007,21(5):59-63.
[45] 李跃林,李志辉,谢耀坚.巨尾桉人工林养分循环研究[J]. 生态学报,2001,21(10):1734-1740.
[46] 李志辉,陈少雄,谢耀坚,等.林分密度对尾巨桉生物量及生产力的影响[J].中南林业科技大学学报,2008, 28(4):49-54.
[47] 朱宾良,李志辉,陈少雄.不同林分密度对尾巨桉生物产量及生产力的影响研究[J].湖南生态科学学报,2007, 13(4):11-14.
[48] 谢贤健,张健,赖挺,等.短轮伐期巨桉人工林地上部分生物量和生产力研究[J].四川农业大学学报,2005, 23(1):66-74.
[49] 杨忠,张建平,王道杰,等.元谋干热河谷桉树人工林生物量初步研究[J].山地学报,2001,19(6):503-510.
[50] 张胜伟.巨尾桉工业原料林生物量研究[D].昆明:昆明理工大学,2008.
[51] 李彬,唐国勇,李昆,等.元谋干热河谷20年生人工恢复植被生物量分配与空间结构特征[J].应用生态学报,2013, 24(6):1479-1486.
[52] 陈昭.肯氏相思与木麻黄人工林养分生物循环研究[D].福州:福建农林大学,2012.
[53] 陈国华.沿海木麻黄更新迹地6种相思的生态适应性研究[D].福州:福建师范大学,2012.
[54] 潘辉,洪伟,陈国荣,等.相思人工林生物量分配格局与林下植被多样性分析[J].福建林业科技,2008,35(1):6-10.
[55] 张尚炬.沿海马尾松台湾相思次生林生态系统生物量及碳贮量的研究[D].福州:福建农林大学,2008.
[56] 陈青霞.不同坡位8年生卷荚相思人工林生物量分配格局[J].林业勘察设计,2011(1):107-110.
[57] 林宇.沿海沙地尾巨桉与厚荚相思造林成效比较[J].林业勘察设计,2009(1):171-174.
[58] 官国栋.滨海沙地卷荚相思和尾巨桉人工林养分生物循环和固碳能力[D].福州:福建农林大学,2013.
[59] 贺韶华.滨海沙地纹荚相思人工林碳氮贮量分配及生物循环[D].福州:福建农林大学,2013.
[60] 卢佳毅.闽南山地两种典型林分的生物量及养分积累与分布[D].北京:中国林业科学研究院,2015.
[61] 郑海水,曾杰,翁启杰,等.薄果相思的生长特性及其改良土壤效果研究[J].广东林业科技,1998,14(1):20-26.
[62] 吴晓芙,胡曰利.刚果12号桉与厚荚相思混交林的养分效应研究[J].中南林学院学报, 2004,24(8): 1-5.
[63] 黄丽铭,薛立,王相娥,等.不同密度下大叶相思幼林的生长和生物量分配格局[J].华南农业大学学报,2008, 29(3):52-55.
[64] 覃世赢.厚荚相思人工幼林生物量与生产力和养分循环的研究[D].南宁:广西大学,2006.
[65] 秦武明,何斌,覃世赢,等.厚荚相思人工林营养元素生物循环的研究[J].水土保持学报,2007,21(4):103-107.
[66] 唐继新.马占相思人工林生长特性及综合效益评价[D].南宁:广西大学,2008.
[67] 梁有祥,秦武明,韦中绵,等.桂西南地区黑木相思生长规律、生物量及生产力研究[J].福建林业科技,2010, 37(2):1-4.
[68] 黄勇,覃静,李松昌,等.卷荚相思人工林生长规律、生物量及生产力研究[J].安徽农业科学,2010,38(22): 12050-12053.
[69] 温中林,郑卫,陈日美.马占相思人工林生物量及营养元素分布研究[J].安徽农业科学,2010,38(27):15040-15045.
[70] 陈燕艺.马尾松马占相思纯林及混交林乔木层生物量和碳储量研究[D].福州:福建农林大学,2014.
[71] 杨绍增,王瑞荣,王达明,等.马尖相思利用价值的研究[J]. 云南林业科技,1996(2):20-30.
[72] 高成杰,唐国勇,孙永玉,等.不同恢复模式下干热河谷幼龄印楝和大叶相思生物量及其分配[J].浙江农林大学学报,2012,29(4):482-490.
[73] 涂育合,董建文.闽中山地湿地松不同立地条件和不同密度的经营效果研究[J].福建林业科技,2002,29(4):9-12.
[74] 陈继祥.柳杉与火力楠、湿地松混交林分结构和生物量的研究[J].亚热带水土保持,2007,19(2):10-12.
[75] 陈振勤.连江县湿地松人工林造林密度试验[J].陕西林业科技,2010(4):24-26.
[76] 叶功富,高伟,郑兆飞,等.闽北高海拔地区湿地松人工林的生物量和生长量研究[J].林业资源管理, 2013(6):76-80.
[77] 郑丽垠.长乐沿海山地不同坡位湿地松生物量分析[J].亚热带农业研究,2013,9(2):94-97.
[78] 刘海,叶功富,朱祥锦,等.马尾松、湿地松幼树生物量及养分积累[J].福建林业科技,2014,41(1):49-54.
[79] 张成典.杉木与湿地松纯林及其混交林的生长及生物量分配格局[J].林业科技,2014,39(5):34-37.
[80] 刘永金,谭绍满.两种湿地松群落的小气候特点及其与林分结构的关系[J].生态学杂志,1991,10(3):5-9.
[81] 曹洪麟,任海,彭少麟.鹤山湿地松人工林的群落结构与能量特征[J].广西植物,1998,1(1):24-28.
[82] 刘之洲.喀斯特地区3种森林生物量和碳储量研究[D].长沙:中南林业科技大学,2017.
[83] 杨佳.封山育林对几种林分生物量与碳储量的影响[D].武汉:华中农业大学,2013.
[84] 肖兴翠,李志辉,唐作钧,等.林分密度对湿地松生物量及生产力的影响[J].中南林业科技大学学报,2011, 31(3):123-129.
[85] 汪企明,石有光.江苏省湿地松人工林生物量的初步研究[J].植物生态学报,1990,14(1):1-12.
[86] 刘学渊.红壤丘陵区湿地松生物量测定[J].林业科技通讯,1984(9):10-13.
[87] 揭建林,詹有生,黄文超,等.京九线井冈山段岗地湿地松林分生产力调查研究[J].江西林业科技,2002(2):17-20.
[88] 刘琪璟,曾慧卿,马泽清.江西千烟洲湿地松人工林碳蓄积及其与水分的关系[J].生态学报,2008,28(11): 5322-5330.
[89] 江波,袁位高,朱光泉,等.马尾松,湿地松和火炬松人工林生物量与生产结构的初步研究[J].浙江林业科技, 1992(5):1-8.
[90] LEHTONEN A, MÄKIPÄÄ R, HEIKKINEN J, et al. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests[J]. Forest Ecology and Management, 2004, 188(1/3): 211-224.
[91] 侯燕南,吴惠俐,项文化,等.亚热带4种森林生物量估算转换参数的研究[J].中南林业科技大学学报,2016, 36(8):57-65.
Differences in Biomass Estimation Coefficients ofspp.,spp. andand Their Key Influencing Factors
LIU Xiaohua1, DU Apeng2, XU Yuxing2, WANG Zhichao2, HUANG Runxia2, ZHU Wankuan2*
(1. Zhanjiang Mangrove National Nature Reserve, Zhanjiang 524022, Guangdong, China; 2. Research Institute of Fast-growing Trees, Chinese Academy of Forestry, Zhanjiang 524022, Guangdong, China)
Data from published literature on biomass measurements ofspp. (including×,×,×and),spp. (including,,,and) andforests in different locations in China were collected and analyzed. The characteristics of three common estimation coefficients of biomass (biomass conversion and expansion factor BCEF, biomass expansion factor BEF, and root/shoot ratio) in different forests and the relationship with stand structure characteristics (stand age, mean DBH, mean tree height, stand density, and standing volume), climatic factors (annual average temperature and annual average precipitation) and topographic factors (elevation) were summarized and differences examined. The results showed that the mean BCEF values ofspp.,spp., andforests averaged 0.66 Mg·m−3, 0.98 Mg·m−3and 0.69 Mg·m−3, respectively, the mean BEF values were 1.26, 1.61 and 1.66 respectively, and the meanvalues were 0.19, 0.23 and 0.22 respectively. The main factors influencing the biomass estimation coefficients offorests were tree height and stand density, while the biomass estimation coefficients offorests were mainly affected by DBH and stocking volume. Estimated biomass coefficients forforests were not significantly influenced by stand structural characteristics, but mainly by climatic factors (mean annual temperature, mean annual precipitation) and topographic factors (elevation). Therefore, differences in biomass estimation coefficients due to stand structure characteristics and climatic and topographic factors should be taken into account when estimating biomass of planted forests at the regional scale.
spp.;spp.;; estimation coefficients of biomass; impact factor
10.13987/j.cnki.askj.2023.04.002
S714.5
A
广东省林业科技创新项目(2022KJCX020);广东省林业生态网络监测平台项目(2022CG644)
刘晓华(1991— ),女,硕士,研究方向为人工林生态监测。E-mail: liuxh324@163.com
竹万宽(1989— ),男,助理研究员,从事森林生态学研究。E-mail:zwk_2015@163.com
