食用豆类资源中酚类化合物研究进展
2023-12-25唐双庆陈禅友王亚珍王红波
唐双庆 陈禅友 王亚珍 王红波
摘要:食用豆类资源中酚类化合物种类丰富。酚酸化合物主要通过莽草酸代谢途径和苯丙烷代谢途径生物合成。酚类化合物作为豆类中主要的活性成分,具有抗氧化、降血糖和抗炎等生理功能。微生物发酵、发芽、膨化挤压和蒸煮烘烤等不同加工方法能够改变豆类酚类化合物的组成和含量,改良酚类化合物的生物利用度。文章综述了食用豆类资源中酚类化合物的结构、生物合成代谢、生理功能以及不同加工方法对酚类化合物的影响,为以酚类化合物为主要活性成分的豆类功能食品的开发提供了理论依据。
关键词:食用豆类资源;酚类化合物;生理功能;加工方法
中图分类号:TS201.2 文献标志码:A 文章编号:1000-9973(2023)12-0200-06
Research Progress on Phenolic Compounds in Edible Legume Resources
TANG Shuang-qing1, CHEN Chan-you2, WANG Ya-zhen3, WANG Hong-bo1,2,3*
(1.Research Center of Food Nutrition and Safety, School of Life Sciences, Jianghan University,
Wuhan 430056, China; 2.Hubei Province Engineering Research Center for Legume Plant,
Wuhan 430056, China; 3.Hubei Enterprise-School Joint Innovation Center of
Healthy Sugar Substitute Product, Wuhan 430056, China)
Abstract: There are abundant types of phenolic compounds in edible legume resources. Phenolic acid compounds are mainly biosynthesized through the metabolic pathways of shikimic acid and phenylpropane. Phenolic compounds, as the main active components in legumes, have the physiological functions such as antioxidation, regulating blood sugar level and anti-inflammation. Different processing methods such as microbial fermentation, germination, extrusion and steaming could change the composition and content of phenolic compounds in legumes and improve the bioavailability of phenolic compounds. In this paper, the structure, biosynthetic metabolism, physiological functions and the effects of different processing methods on phenolic compounds in edible legume resources are reviewed, which have provided a theoretical basis for the development of legume functional foods with phenolic compounds as the main active components.
Key words: edible legume resources; phenolic compounds; physiological functions; processing methods
我國食用豆类资源丰富,豆类加工制备的食品深受消费者喜爱。豆类营养物质丰富,还富含多酚、多肽、γ-氨基丁酸和低聚糖等多种生物活性物质,使豆类食品具有抗氧化、降血糖、降血脂和预防肥胖等生理功能[1-5]。豆类制品对改善国民饮食结构和提高国民健康水平具有不可替代的作用。酚类物质作为豆类重要的功能成分,是食品科学研究关注的热点。目前,豆类多酚化合物的结构、生物合成代谢和生理功能研究已经取得一定的进展。不同加工方法对豆类酚类化合物的组成和含量具有显著影响,已经引发关注。豆类作为我国重要的食品加工资源,豆类酚类化合物的深入研究有利于增加豆类食物的消费,服务国家乡村振兴战略。
1 豆类酚类化合物结构研究
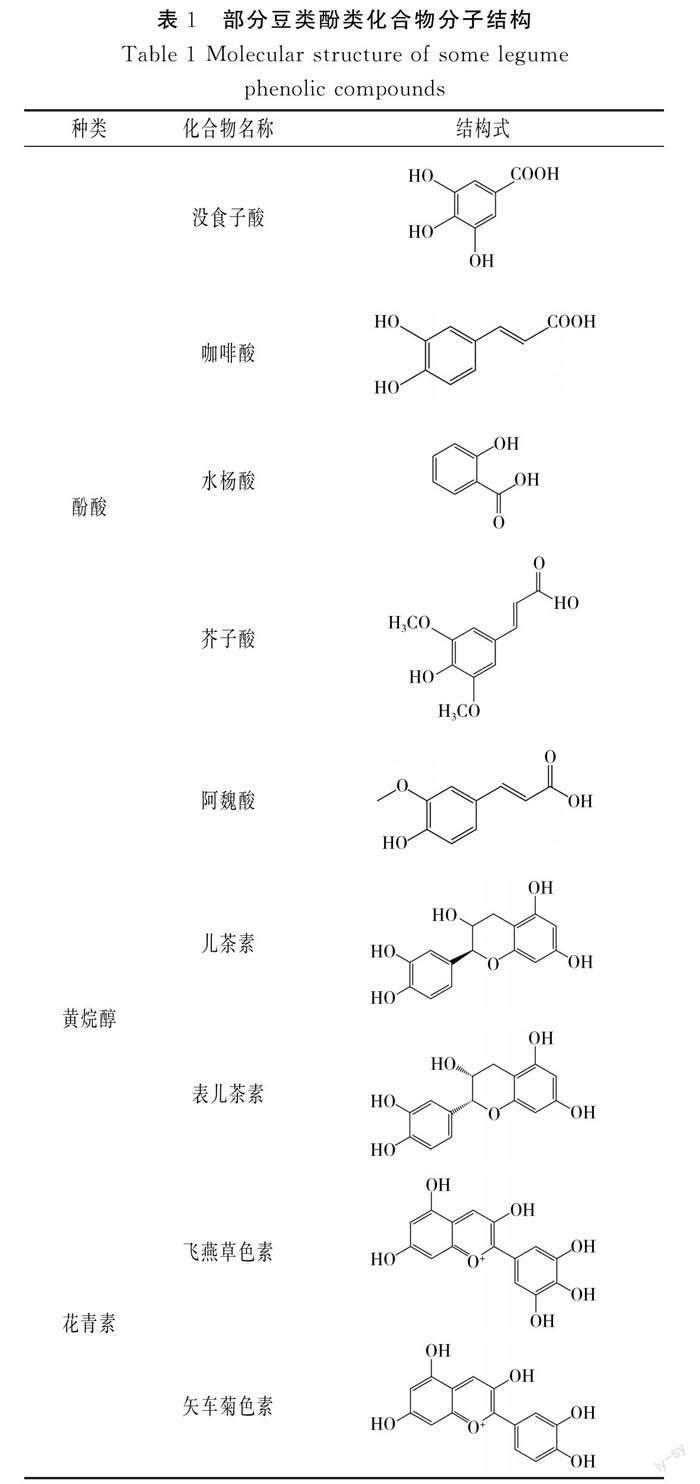
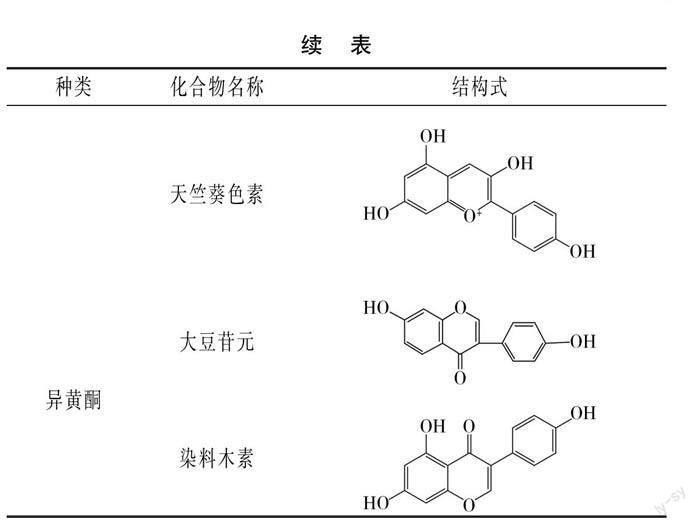
酚类化合物是一类广泛存在于植物体内的次生代谢产物,主要存在于植物体的果实、皮、根和叶等组织器官中,不同植物所含酚类化合物组成不同。酚类化合物包含一个或多个羟基基团,自然界中存在8 000多种酚类物质,可以分为类黄酮和非类黄酮两类。非类黄酮包括酚酸、木脂素和二苯乙烯。类黄酮是多酚两个苯环之间通过三碳链连接而成的C6-C3-C6结构,包括黄酮醇、黄酮、黄烷醇、花青素和异黄酮等[6-7]。豆类含有的酚类化合物主要有酚酸、黄烷醇、黄酮、花青素和异黄酮等,豆类常见酚类化合物部分结构见表1。
酚酸类化合物是由苯环和有机羧酸基团结合的C6-C1结构,是豆类酚类化合物的重要成分,包括没食子酸、阿魏酸、对香豆酸和芥子酸等。黄酮类物质主要存在于大豆和具有多种颜色种皮的豆类中,黑豆种皮是黄酮类物质的有效来源[8]。李文婷等[9]测出赤豆酚类提取物酚酸含量为3.44 mg/g,而黄酮含量为2.50 mg/g;赤豆中含有原儿茶酸、对羟基苯甲酸、绿原酸和香豆酸等酚酸化合物以及芦丁、芹菜素、槲皮素、原花青素和牡荆素等黄酮类化合物。Luthria等[10]分析得出菜豆总酚酸含量为19.1~48.3 mg/100 g,菜豆中含有阿魏酸、对香豆酸和芥子酸等酚类化合物。Yang等[11]鉴定出绿豆中含有牡荆素、异牡荆素、槲皮素、黄豆黄素和菊苣苷等16种酚类化合物。Telles等[12]检测出10种隶属于菜豆属和豇豆属的豆类中含有没食子酸、绿原酸、咖啡酸、香草醛和丁香酸等酚类化合物。蚕豆中含有对香豆酸、阿魏酸、表儿茶素、表儿茶素葡萄糖苷、表儿茶素没食子酸酯和没食子酸原花青素等酚类化合物[13]。
2 豆类酚类化合物合成代谢研究
2.1 酚酸类化合物的合成代谢
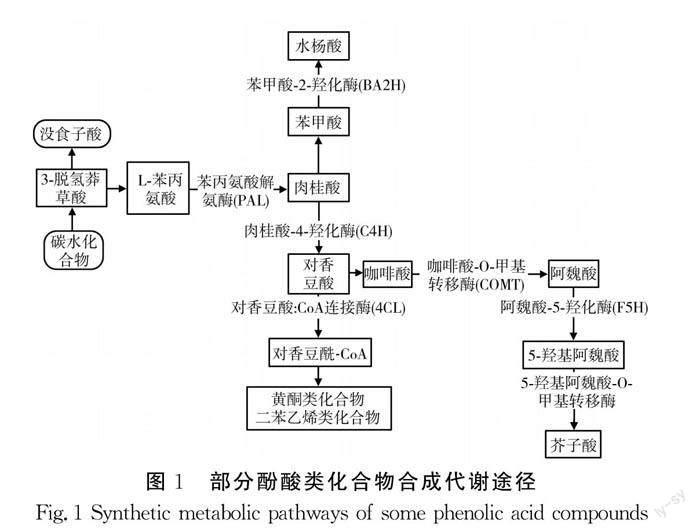
酚酸类化合物主要通过莽草酸代谢途径和苯丙烷代谢途径进行生物合成。莽草酸途径是由3-脱氢莽草酸生成没食子酸,并且通过莽草酸途径和糖酵解途径共同作用生成苯丙氨酸;苯丙烷代谢途径由苯丙氨酸开始,分别在苯丙氨酸解氨酶(PAL)、肉桂酸-4-羟化酶(C4H)、苯甲酸-2-羟化酶(BA2H)、咖啡酸-O-甲基转移酶(COMT)、阿魏酸-5-羟化酶(F5H)和5-羟基阿魏酸-O-甲基转移酶的催化下,依次生成肉桂酸、对香豆酸、水杨酸、阿魏酸、5-羟基阿魏酸和芥子酸。这些化合物作为底物参与下一步反应,形成苯丙类化合物。此外,以对香豆酸为底物,在对香豆酸:CoA连接酶(4CL)等作用下,参与后续各类反应生成黄酮类和二苯乙烯类化合物[14-15]。酚酸类化合物部分合成代谢途径示意图[14]见图1。
酚类物质合成代谢途径涉及多种酶促反应,易受内部与外部因素的调控。光照、温度、病原微生物和外源性诱发剂等均能显著影响苯丙烷代谢途径[15]。Mahmood等[16]研究發现酪氨酸可激活苯丙烷代谢途径,经酪氨酸灌注绿豆植株,PAL活性显著增加。外源γ-氨基丁酸可以调节PAL活性,促进发芽大豆生长,诱导多酚物质积累,增强抗氧化酶活性和体外自由基清除活性[17]。PAL活性增强能够强化几种核心底物合成,例如阿魏酸、咖啡酸、对香豆酸和苯甲酸,有助于酚类代谢物的形成[18]。壳聚糖可以有效启动次生代谢物的形成和酚类物质的积累。Manoj等[19]研究发现壳聚糖灌注可强化菜豆几种酚酸化合物的生物合成。
2.2 黄酮类化合物的合成代谢
黄酮类化合物的生物合成是以对香豆酰-CoA和丙二酰CoA为前体,由查尔酮合酶(CHS)和查尔酮还原酶(CHR)催化,分别生成柚皮素查尔酮和异甘草素,在查尔酮异构酶(CHI)和异黄酮合成酶(IFS)的共同催化下,生成大豆苷和染料木素异黄酮化合物。此外,以柚皮素为底物,经过一系列酶促反应,生成儿茶素和表儿茶素等。部分类黄酮化合物生物合成代谢途径[14]见图2。
黄酮生物合成代谢途径受到各种环境因素调节,从而改变黄酮化合物的种类[14,20]。温度、光照、水分、辐照和生物因子等因素均能显著影响类黄酮代谢途径[21]。蔗糖通过提供己糖分子引导苯丙烷的生物合成,强化黄酮、黄酮醇和异黄酮的合成代谢 [22]。
3 豆类酚类化合物生理活性研究
食用豆类资源含有多种酚类化合物。酚类化合物具有抗氧化、降血糖、降血脂和抗炎等多种生理功能。国内外研究报道表明酚类物质作为植物中的天然活性成分,对人体健康有着重要的作用。
3.1 酚类化合物的抗氧化作用
酚类化合物最重要的一种生理活性是抑制自由基,延缓机体损伤。自由基指的是一系列具有一个或者多个不成对电子的高活性中间体,可以诱导生物系统中底物的氧化分解[23]。自由基参与许多生理生化过程,如细胞中的信号传导等,但累积过量的自由基会引起机体损伤。人体内含有谷胱甘肽过氧化物酶(glutathione peroxidase,GPx)、超氧化物歧化酶(superoxide dismutase,SOD)和过氧化氢酶(catalase,CAT)等多种天然抗氧化酶,可以预防或减少对靶向底物的氧化损伤。如果自由基的产生和消除间的平衡被细胞中抗氧化剂不足打破,就会发生氧化应激,造成机体表现出病理性特征[24]。酚类物质具有良好的抗氧化效果。酚类化合物抗氧化过程主要分为两个阶段:自由基捕获阶段:S-OO·+AH→SOOH+A;自由基终止阶段:A→非自由基。其中,S是氧化物质,S-OO·是S的过氧自由基,AH是抗氧化剂,A是抗氧化自由基[25]。自由基捕获阶段是可逆过程,但自由基终止阶段是不可逆过程,必须产生稳定的自由基终止产物[25]。酚类抗氧化机制主要有单电子转移机制(single electron transfer,SET)、顺序质子损失电子转移机制(sequential proton loss electron transfer,SPLET)、氢原子转移机制(hydrogen atom transfer,HAT)、顺序电子质子转移机制(single electron transfer followed by proton transfer,SETPT)和自由基加合物形成机制(radical adduct formation,RAF)5种,不同酚类物质的抗氧化机制不同。儿茶素和水杨酸等酚酸类物质的抗氧化机制主要是氢原子转移机制、单电子转移机制和顺序质子损失电子转移机制[26]。黄酮醇类、黄烷醇和异黄酮类等黄酮类化合物的抗氧化机制主要是氢原子转移机制、单电子转移机制、顺序质子损失电子转移机制以及自由基加合物形成机制等[27]。赵艳等[28]对红豆、扁豆、红芸豆、蚕豆和鹰嘴豆5种豆类提取物进行体外抗氧化研究,发现5种豆类总酚含量为1.24~15.07 mg/g,总黄酮含量为0.68~17.55 mg/g,且扁豆和红豆的总酚含量高,而鹰嘴豆的总酚含量最低;红芸豆的总黄酮含量最高,鹰嘴豆的总黄酮含量最低;5种豆类提取物具有较好的清除1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基和羟基自由基活性,以及良好的铁还原能力和亚铁离子螯合能力。García等[29]测定不同储存时间下黑豆总酚含量的变化规律,研究发现270 d以后,黑豆总酚含量从(333±6) mg GAE/g DW下降至(240±9) mg GAE/g DW,而DPPH自由基清除活性由(2 188±5) mg TE/g DW上升至(2 640±14) mg TE/g DW。Hao等[30]利用乙酸乙酯、正丁醇、石油醚和水4种溶剂提取绿豆皮中的酚类化合物,研究发现异牡荆素和牡荆素对H2O2在HaCaT和HSF中引起的损伤和衰老起到良好的保护作用。丁羽萱等[31]在大豆发芽过程中添加外源γ-氨基丁酸诱导苯丙氨酸解氨酶基因表达,使总酚和游离酚含量增加,DPPH自由基清除活性和2,2′-联氮-双-(3-乙基苯并噻唑啉-6-磺酸)二胺盐(2,2'-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) ammonium salt,ABTS)阳离子自由基清除活性均增强。
3.2 酚类化合物的降血糖作用
糖尿病患者以Ⅱ型糖尿病为主。Ⅱ型糖尿病是一种高葡萄糖血浆水平疾病,涉及多种细胞途径,包括胰岛素分泌、胰岛素抵抗和碳水化合物吸收[32]。多酚调节血糖水平常见方式即通过抑制小肠中α-葡萄糖苷酶,减少葡萄糖吸收,或者增加胰岛素和减少胰高血糖素分泌来降低葡萄糖水平,以及调节肠道菌群等对血糖起调节作用[33-35]。多酚的降糖机制可以通过不同方式发生:第一,通过多种途径中的代谢活动,改变底物或中间产物浓度,从而改变细胞信号传导;第二,可以作为转录因子配体;第三,直接与信号通路相互作用。所有机制对基因表达具有修饰作用,从而对胰腺、脂肪和肝脏等组织和器官起到积极影响,达到控制Ⅱ型糖尿病的作用[32]。黄烷酮通过促进糖原合成,降低葡萄糖-6-磷酸酶(glucose-6-phosphatase,G6P)和磷酸烯醇式丙酮酸羧激酶(phosphoenol pyruvate carboxykinase,PEPCK)的表达,抑制糖异生,起到降糖作用[36]。对香豆酸、肉桂酸和香草酸等酚酸化合物能够通过抑制α-葡萄糖苷酶和增强胰岛素分泌等方式降糖。通过进食富含黄酮醇、酚酸和异黄酮的食物可降低Ⅱ型糖尿病风险[37]。张雪仓等[38]研究发现白芸豆及复合物具有降血糖作用,在餐前服用相关产品,能够有效降低进食后的血糖反应。王德萍等[39]研究鹰嘴豆提取物对糖尿病小鼠的影响,发现鹰嘴豆的提取物能够显著降低糖尿病小鼠血糖水平, 减轻胰岛素抵抗。吴琼等[40]发现紫皮豇豆多酚提取物能够明显抑制α-葡萄糖苷酶活性。
3.3 其他生理作用
酚类化合物可在炎症期间调节细胞信号传导,起到抵抗炎症的作用。在脂多糖等炎症刺激物的刺激下,巨噬细胞分泌多种促炎细胞因子,如肿瘤坏死因子-α(TNF-α)和白细胞介素等,它们参与多种疾病发病机制,影响人体健康。酚类化合物可在不同水平上调节炎症级联反应,包括编码白细胞介素的炎症基因的表达[41]。García-Lafuente等[41]研究发现豆类酚类提取物能够通过抑制一氧化氮合成酶(inducible nitric oxide synthase,iNOS)的mRNA表达来减少脂多糖刺激的巨噬细胞中NO的产生,并且抑制巨噬细胞RAW 246.7分泌的白细胞介素IL-1β和IL-6以及TNF-α基因的表达。酚类化合物对预防心血管疾病也具有积极作用,酚酸、类黄酮和花青素等酚类化合物可以抑制活性氧和其他自由基的产生,有利于减少低密度脂蛋白(LDL-c)过氧化,而LDL-c是动脉粥样硬化生成的重要危险因素[42]。刘锦绣等[43]研究发现异黄酮对癌细胞具有抗肿瘤作用,在β-葡萄糖苷酶作用下,糖苷型异黄酮转化为苷元型异黄酮,提升了抗肿瘤能力。没食子酸和鞣花酸等可以通过调控核因子相关因子E2/血红素氧合酶-1(nuclear factor E2-related factor 2/heme oxygenase-1,Nrf2/HO-1)信号通路抑制酒精肝损伤[44]。酚类化合物还具有抑菌作用,豆类酚类提取物能够抑制米曲霉等真菌产生的α-淀粉酶活性,通过主成分分析证明绿原酸和没食子酸等酚类化合物对真菌毒素(黄曲霉毒素)具有抑制作用[12]。
4 不同加工方法对豆类酚类化合物影响
豆类产品初加工通常采用浸泡、脱壳和研磨等常规方法。二次加工方法包括发酵、发芽、挤压和烘烤等[45]。不同加工方法对豆类酚类组成和含量均具有显著影响。
4.1 发酵加工处理
微生物发酵作用能够产生有利于物质转化的酶,这些酶类通常有利于细胞壁中结合的生物活性物质得以释放,从而改善豆类营养品质,赋予豆类食品独特的风味[46]。在发酵过程中,豆类含有的结合态酚在微生物产生的β-葡萄糖苷酶作用下,变为游离态,从而使总酚含量提高,抗氧化活性也随之增强。James等[47]将红豆总酚含量增加归因于微生物酶活性,有利于生成更能利用的植物次生代谢物比如黄酮类、单宁和生物碱等物质。Li等[48]研究发现,利用干酪乳杆菌发酵能够提高大豆酚类化合物含量和抗氧化活性。Sanjukta等[49]研究表明枯草芽孢杆菌发酵大豆和黑豆总酚含量从1.93 mg GAE/g和1.64 mg GAE/g分别增加至7.9~8.4 mg GAE/g和6.9~7.5 mg GAE/g,且大豆的DPPH自由基清除率从1.3 mg AAE/g增加至29.9 mg AAE/g。徐春明等[50]发现纳豆发酵过程中, 总酚含量提高了65.12%,ABTS+自由基清除能力和氧化自由基吸收能力(oxygen radical absorbance capacity,ORAC)提高。大豆异黄酮是大豆中重要的生物活性物质,β-葡萄糖苷酶可以将大豆异黄酮的糖苷结构转化为苷元结构,提升大豆异黄酮活性[51]。黄玉军等[52]比较12种乳酸菌发酵豆乳产大豆異黄酮苷元的能力,发现菌株编号为58和m91的两种细菌发酵后大豆异黄酮苷元含量分别为59.64 mg/L和58.06 mg/L,均是原豆乳的8倍以上。
4.2 发芽加工处理
发芽是一种天然的生理生化过程,可以充分增强豆类的营养品质,减少或消除植酸等抗营养成分,还能增加生物活性物质含量和抗氧化能力。在发芽过程中,部分结合态多酚可以转化为游离态多酚,同时合成新的多酚物质,从而增加多酚含量[53]。Hung等[54]研究发现,发芽绿豆比未发芽绿豆总酚含量提高,可能是由于在发芽过程中,与碳水化合物或蛋白质等有机物质结合的酚类物质通过酶水解作用,结合酚类物质从细胞壁中释放,从而增加酚类物质的含量。此外,随着发芽时间延长,环境胁迫作用形成的防御机制强化了酚酸化合物的生物合成和累积,导致总酚含量增加。Ma等[55]研究发现大豆发芽过程中,参与酚酸合成的PAL、C4H和4CL关键酶活性增强,没食子酸、丁香酸、对香豆酸、阿魏酸和芥子酸等8种酚酸以游离或结合方式存在,并且其含量均显著增加。
4.3 膨化挤压加工处理
挤压加工是生产膨化食品的有效技术,具有出色的性能,能够通过强烈混合引起原料的结构变化,通过多酚含量改变、淀粉糊化、色素降解、美拉德褐变和抗营养因子(如植酸、凝集素、胰蛋白酶)的失活等方式实现成分的分散和均质化,改善产品特性。Felix-Medina等[56]使用玉米/普通豆混合物生产一种健康的即食膨化零食,从酚类提取物中鉴定出阿魏酸、二聚阿魏酸和对香豆酸3种酚酸以及柚皮素、山奈酚和甲基异黄酮3种类黄酮。Arribas等[57]研究显示挤压过程中的高温高压可能导致热不稳定性酚类化合物的分解,某些多酚(如单宁)的聚合降低了酚类物质的可萃取性,使得挤压过程大米-菜豆酚类化合物含量低于非挤压过程。
4.4 其他加工处理
浸泡和脱壳等加工方式也会降低豆类酚类物质含量。蚕豆和菜豆壳中富含酚类化合物,脱壳后会降低酚类物质含量[58]。蒸煮有利于豆类生物活性物质的释放。Sangsukiam等[59]研究发现高压蒸煮红豆后4-羟基苯甲酸、香草酸、丁香酸、对香豆酸、牡荆素和芦丁的含量均显著增加。Corzo-Ríos等[60]发现蒸煮和烘烤对不同品种蚕豆酚类物质含量的影响不同,蒸煮导致HTP蚕豆总酚含量上升,HTA蚕豆和HPA蚕豆总酚含量下降;烘烤导致HTP、HTA和HPA 总酚含量均上升。
5 总结与展望
食用豆类资源中天然酚类化合物种类丰富。运用现代食品加工技术,开发富含酚类化合物的豆类功能食品将具有重要的经济社会价值。豆类中酚类化合物的理论和应用研究还需加强。豆类中结合态酚类物质的存储和结合机理以及微生物发酵作用对豆类酚类物质分子结构的修饰和功能改良将是今后豆类酚类化合物研究的两个重要方向。
参考文献:
[1]MEJRI F, SELMI S, MARTINS A, et al. Broad bean (Vicia faba L.) pods: a rich source of bioactive ingredients with antimicrobial, antioxidant, enzyme inhibitory, anti-diabetic and health-promoting properties[J].Food & Function,2018,9(4):2051-2069.
[2]谢莎丽,石元刚.大豆低聚糖及大豆多肽对大鼠血脂代谢的影响[J].第三军医大学学报,2006(9):945-948.
[3]辛玥,宋萧萧,王玉箫,等.豇豆不同部位多糖结构特征及抗氧化性能比较[J].食品科学,2022,43(16):61-67.
[4]郑虹君,朱叙丞,李耀基,等.白蕓豆功能成分、生物活性及其产品开发研究进展[J].中国粮油学报,2022,37(12):277-285.
[5]俞德慧,杨杨,陈凤莲,等.γ-氨基丁酸及其在谷物发酵食品中的研究进展[J].食品与发酵工业,2022,48(11):290-296.
[6]LUND M N. Reactions of plant polyphenols in food: impact of molecular structure[J].Trends in Food Science & Technology,2021,112:241-251.
[7]CROZIER A, JAGANATH I B, CLIFFORD M N. Dietary phenolics: chemistry, bioavailability and effects on health[J].Natural Product Reports,2009,26:1001-1043.
[8]CHVEZ-SANTOSCOY R A, GUTIRREZ-URIBE J A, SERNA-SALDIVAR S O, et al. Production of maize tortillas and cookies from nixtamalized flour enriched with anthocyanins, flavonoids and saponins extracted from black bean (Phaseolus vulgaris) seed coats[J].Food Chemistry,2016,192:90-97.
[9]李文婷,顿倩,李红艳,等.基于UPLC-Q TOF-MS和HPLC-QQQ-MS技术的赤豆酚类化合物定性、定量分析及其抗氧化能力分析[J].食品科学,2019,40(8):112-118.
[10]LUTHRIA D L, PASTOR-CORRALES M A. Phenolic acids content of fifteen dry edible bean (Phaseolus vulgaris L.) varieties[J].Journal of Food Composition and Analysis,2006,19(2-3):205-211.
[11]YANG Q Q, GE Y Y, GUNARATNE A, et al. Phenolic profiles, antioxidant activities, and antiproliferative activities of different mung bean (Vigna radiata) varieties from Sri Lanka[J].Food Bioscience,2020,37:100705.
[12]TELLES A C, KUPSKI L, FURLONG E B. Phenolic compound in beans as protection against mycotoxins[J].Food Chemistry,2017,214:293-299.
[13]AMAROWICZ R, SHAHIDI F. Antioxidant activity of broad bean seed extract and its phenolic composition[J].Journal of Functional Foods,2017,38:656-662.
[14]CROZIER A, JAGANATH I B, CLIFFORD M N. Phenols, Polyphenols and Tannins: An Overview[M].Oxford:Blackwell Publishing Ltd.,2007:14-15.
[15]尚军,吴旺泽,马永贵.植物苯丙烷代谢途径[J].中国生物化学与分子生物学报,2022,38(11):1467-1476.
[16]MAHMOOD S, WAHID A, AZEEM M, et al. Tyrosine or lysine priming modulated phenolic metabolism and improved cadmium stress tolerance in mung bean (Vigna radiata L.)[J].South African Journal of Botany,2022,149:397-406.
[17]WANG L D, LI X Q, GAO F, et al. Effect of ultrasound combined with exogenous GABA treatment on polyphenolic metabolites and antioxidant activity of mung bean during germination[J].Ultrasonics Sonochemistry,2023,94:106311.
[18]JEONG H, SUNG J, YANG J, et al. Effect of sucrose on the functional composition and antioxidant capacity of buckwheat (Fagopyrum esculentum M.) sprouts[J].Journal of Functional Food,2018,43:70-76.
[19]MANOJ B S, GUPTA M, JEELANI M I, et al. Chitosan augments bioactive properties and drought resilience in drought-induced red kidney beans[J].Food Research International,2022,159:111597.
[20]趙莹,杨欣宇,赵晓丹,等.植物类黄酮化合物生物合成调控研究进展[J].食品工业科技,2021,42(21):454-463.
[21]周明,沈勇根,朱丽琴,等.植物黄酮化合物生物合成、积累及调控的研究进展[J].食品研究与开发,2016,37(18):216-221.
[22]LYU C Y, ZHANG X Y, HUANG L, et al. Widely targeted metabolomics analysis characterizes the phenolic compounds profiles in mung bean sprouts under sucrose treatment[J].Food Chemistry,2022,395:133601.
[23]LIU Z Q. Bridging free radical chemistry with drug discovery: a promising way for finding novel drugs efficiently[J].European Journal of Medicinal Chemistry,2020,189:112020.
[24]ZHU L, LUO M Y, ZHANG Y F, et al. Free radical as a double-edged sword indisease: deriving strategic opportunities for nanotherapeutics[J].Coordination Chemistry Reviews,2023,475:214875.
[25]MASUDA T, AKIYAMA J, FUJIMOTO A, et al. Antioxidation reaction mechanism studies of phenolic lignans, identification of antioxidation products of secoisolariciresinol from lipid oxidation[J].Food Chemistry,2010,123(2):442-450.
[26]陈金祥.酚酸抗氧化活性的构效关系及抗氧化机制的研究[D].太原:中北大学,2020.
[27]谢虹.黄酮类化合物的RAF抗氧化机制[D].广州:广州中医药大学,2019.
[28]赵艳,杜双奎,王晗欣,等.5种杂豆体外抗氧化性研究[J].中国粮油学报,2015,30(10):6-10.
[29]GARCA M N, AVENDAO M P, BEZ D A, et al.Phenolic compounds profile by UPLC-ESI-MS in black beans and its distribution in the seed coat during storage[J].Food Chemistry,2022,395:133638.
[30]HAO R L, LI M Q, LI F, et al. Protective effects of the phenolic compounds from mung bean hull against H2O2-induced skin aging through alleviating oxidative injury and autophagy in HaCaT cells and HSF cells[J].Science of the Total Environment,2022,841:156669.
[31]丁羽萱,王尧,姚羿安,等.外源γ-氨基丁酸对发芽大豆酚类物质富集及抗氧化能力的影响[J].食品科学,2021,42(13):72-78.
[32]DAMIN-MEDINA K, SALINAS-MORENO Y, MILENKOVIC D, et al. In silico analysis of antidiabetic potential of phenolic compounds from blue corn (Zea mays L.) and black bean (Phaseolus vulgaris L.)[J].Heliyon,2020,6(3):3632.
[33]INTHONGKAEW P, CHATSUMPUN N, SUPASUTEEKUL C, et al. α-Glucosidase and pancreatic lipase inhibitory activities and glucose uptake stimulatory effect of phenolic compounds from Dendrobium formosum[J].Revista Brasileira de Farmacognosia,2016,7:475-482.
[34]HUANG C N, WANG C J, YANG Y S, et al.Hibiscus sabdariffa polyphenols prevent palmitate-induced renal epithelial mesenchymal transition by alleviating dipeptidyl peptidase-4-mediated insulin resistance[J].Food & Function,2017,27(4):480-487.
[35]黄修晴,初众,房一明,等.植物多酚降血糖机制的研究进展[J].食品工业科技,2021,42(18):461-469.
[36]JUNG U J, LEE M K, PARK Y B, et al. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mRNA levels in type-2 diabetic mice[J].The International Journal of Biochemistry & Cell Biology,2006,38(7):1134-1145.
[37]GOLOVINSKAIA O, WANG C K. The hypoglycemic potential of phenolics from functional foods and their mechanisms[J].Food Science and Human Wellness,2023,12(4):986-1007.
[38]张雪苍,薛红梅.白芸豆-馬齿苋餐前控糖片的工艺研究及控糖效果评价[J].食品科技,2020,45(7):38-43.
[39]王德萍,安馨,鱼晓敏,等.鹰嘴豆醇提物降血糖作用研究[J].食品研究与开发,2019,40(13):21-25.
[40]吴琼,王明月,吕岱竹,等.紫皮豇豆多酚类物质的含量及其降糖活性研究[J].中国调味品,2019,44(10):59-62.
[41]GARCA-LAFUENTE A, MORO C, MANCHN N, et al. In vitro anti-inflammatory activity of phenolic rich extracts from white and red common beans[J].Food Chemistry,2014,161:216-223.
[42]VIEIRA N M, PEGHINELLI V V, MONTE M G, et al. Beans comsumption can contribute tothe prevention of cardiovascular disease[J].Clinical Nutrition,2023,54:73-80.
[43]刘锦绣,陈伟,李靖,等.混菌发酵豆豉中异黄酮含量及对小鼠肺癌细胞的抑制作用[J].食品与发酵工业,2013,39(3):61-66.
[44]刘晓丽,杨冰鑫,陈柳青,等.余甘子多酚对体外酒精性肝损伤的保护作用[J].中国食品学报,2021,21(10):108-115.
[45]KARTIKEYAN A, VASUDEVAN V, JOHN P A, et al. Effect of incubation period on the glycosylated protein content in germinated and ungerminated seeds of mung bean (Vigna radiata (L.) Wilczek)[J].International Journal of Biological Macromolecules,2022,217:633-651.
[46]SHAN L Y, TYAGI A, CHEN X Q, et al. Potential anti-obesity effect of fermented adzuki beans and their untargeted metabolomics using UHPLC-QTOF-MS[J].Food Bioscience,2023,52:102380.
[47]JAMES S, NWABUEZE T U, NDIFE J, et al. Influence of fermentation and germination on some bioactive components of selected lesser legumes indigenous to Nigeria[J].Journal of Agriculture and Food Research,2020,2:100086.
[48]LI S L, JIN Z Y, HU D J, et al. Effect of solid-state fermentation with Lactobacillus casei on the nutritional value, isoflavones, phenolic acids and antioxidant activity of whole soybean flour[J].LWT-Food Science and Technology,2020,125:109264.
[49]SANJUKTA S, RAI A K, MUHAMMED A, et al. Enhancement of antioxidant properties of two soybean varieties of Sikkim Himalayan region by proteolytic Bacillus subtilis fermentation[J].Journal of Functional Foods,2015,14:650-658.
[50]徐春明,陳莹莹,庞高阳.纳豆发酵和体外模拟消化中活性物质含量及抗氧化活性的变化研究[J].现代食品科技,2016,32(11):78-84.
[51]李参参,张淦,常徽.微生物发酵法转化大豆异黄酮研究进展[J].食品研究与开发,2023,44(12):218-224.
[52]黄玉军,周帆,于俊娟,等.高转化大豆异黄酮乳酸菌的筛选及在豆乳中的发酵特性[J].食品研究与开发,2021,42(3):157-162.
[53]LIU Y, SU C Y, SALEH A S M, et al. Effect of germination duration on structural and physicochemical properties of mung bean starch[J].International Journal of Biological Macromolecules,2020,154:706-713.
[54]HUNG P V, YEN N T H, PHI N T L, et al. Nutritional composition, enzyme activities and bioactive compounds of mung bean (Vigna radiata L.) germinated under dark and light conditions[J].LWT-Food Science and Technology,2020,133:110100.
[55]MA Y, WANG P, GU Z X, et al. Effects of germination on physio-biochemical metabolism and phenolic acids of soybean seeds[J].Journal of Food Composition and Analysis,2022,112:104717.
[56]FELIX-MEDINA J V, GUTIERREZ-DORADO R, LOPEZ-VALENZUELA J A, et al. Nutritional, antioxidant and phytochemical characterization of healthy ready-to-eat expanded snack produced from maize/common bean mixture by extrusion[J].LWT-Food Science and Technology,2021,142:111053.
[57]ARRIBAS C, PEREIRA E, BARROS L, et al.Healthy novel gluten-free formulations based on beans, carob fruit and rice: extrusion effect on organic acids, tocopherols, phenolic compounds and bioactivity[J].Food Chemistry,2019,292:304-313.
[58]何磊,于宁,陈颖.常见加工方式对杂豆品质的影响与调控[J].中国粮油学报,2023,38(1):177-185.
[59]SANGSUKIAM T, DUANGMAL K. Changes in bioactive compounds and health-promoting activities in adzuki bean:effect of cooking and in vitro simulated gastrointestinal digestion[J].Food Research International,2022,157:111371.
[60]CORZO-ROS L J, JIMNEZ-MARTNEZ C, CID-GALLEGOS M S, et al. Chemical and non-nutritional modification of faba bean (Vicia faba) due to the effect of roasting and boiling[J].International Journal of Gastronomy and Food Science,2022,30:100622.
收稿日期:2023-06-14
基金项目:武汉市属高校产学研项目(CXY202202);湖北省重点研发计划项目(2022BBA0064);湖北省高价值知识产权培育工程项目(鄂知发[2021]2号);江汉大学一流学科建设重大专项资助计划(2023XKZ023)
作者简介:唐双庆(1997-),女,硕士,研究方向:食品微生物。
*通信作者:王红波(1982-),男,副教授,博士,研究方向:食品加工与安全。
