塘类海绵生境鸟类多样性特征及其影响因素研究
2023-12-21陈珏洁郭雨蕊阳佩琳刘家琳
陈珏洁 郭雨蕊 阳佩琳 刘家琳
1 研究背景
2020年联合国生物多样性大会强调了生物多样性保护、恢复、监测维护的重要性[1-2]。城市生态环境中,绿地的生物多样性效益在近年来受到持续关注[3-5]。在此背景下,海绵生境作为发挥生物多样性效益的重要小微生境载体[6-7],国内对其生物多样性提升的实践与研究还较为不足。目前,国内对于海绵生境的相关研究主要集中在雨洪管理调控绩效层面[8-11],有少量研究关注其植物生境营建的合理性[12-13],对于海绵生境生物多样性效益的监测与优化的研究不足。国际研究表明,海绵生境具有提高城市生物多样性的潜力[14],在城市动植物多样性营建中发挥着重要的生态功能,为传粉昆虫、鸟类或其他动物提供栖息地[15-19]。海绵设施中的干塘与湿塘这一类塘类设施具有蓄水层空间[20],对于鸟类这一生态敏感指示种群具有吸引力,能作为小微生境为其提供躲藏、栖息、饮水、觅食功能,充当鸟类活动过程中的关键踏脚石[6,14,19],其具有增加生物多样性、调节城市微气候的效益[7]。本研究聚焦具有蓄水层空间、功能结构有一定相似性的塘类海绵设施,包含干塘、湿塘等。对其4个季度的鸟类数据进行持续观测,并探究塘类海绵生境的设计因子与周边环境因子对鸟类群落多样性特征的影响。本文着重探究了塘类海绵生境中鸟类群落多样性特征,以及影响塘类生境群落鸟类多样性的关键因素,相关研究结果对于海绵城市精细化营建、提升城市生物多样性具有指导价值。
2 研究方法
2.1 研究对象
研究区域为重庆市渝北区悦来国际会展城(29°41′26″N~29°43′6″N,106°31′30″E~106°31′50″E),是国内首批海绵城市建设试点区域之一,总面积为18.67km2,其植被带属于重庆中西部丘陵低山常绿阔叶林区,临嘉陵江段下游,属亚热带季风性湿润气候,常年平均气温18.3℃,年平均降雨量为1 210.8mm。目前该区域已完工的海绵绿地建设面积约4km2,涉及6个子流域、28个雨水管理片区[21]。
本研究选取悦来国际会展城中的塘类海绵生境为研究对象,涉及干塘12个、湿塘12个(图1)。研究样地具有典型干塘或湿塘的结构特征及雨水受纳功能,所选样地分布于公园绿地、广场用地、公共服务设施用地附属绿地、居住用地附属绿地、溪河防护绿地和生态保育绿地中,其周边环境特征具有差异性。所有样地均已建成1年以上,具有稳定的植被结构,样地间隔直线距离大于200m[22]。

图1 悦来国际会展城塘类海绵塘样地航拍图片(a~l为湿塘样地,m~x为干塘样地,重庆悦来投资集团有限公司提供)
2.2 调查方法
2.2.1 鸟类调研方法
鸟类调查在2021年10月,2022年1、4、6月,采用样点法对样地生境中4个季节的鸟类情况进行监测。每月调查时,样点调查先后顺序进行调换,以排除固定调查顺序对采集数据产生干扰。调查当日均为晴天,于日出后3h内鸟类活跃时段进行观测。观测时,先静置5min再采用8×32倍双筒望远镜进行观察。每个样点以生境驳岸线或表面蓄水层边缘向外10m、距地面20m高度为观测范围,采用直接计数法,记录样点内听到、看到的所有鸟类,统计其种类、数量等信息,偶尔采用拍照记录、叫声识别等方法进行内业辅助鉴定。鸟种的识别鉴定参照《中国鸟类野外手册》[23]。鸟类居留型及食性划分参照《中国鸟类分类与分布名录(第三版)》[24]。此外,重庆市观鸟协会专家在研究过程中协助进行鸟类识别。
2.2.2 植被调研方法
塘类海绵生境植物调查采用样方法,每个生境样地依据其面积,并考虑设施中植被结构与种类的异质性设置1个或多个整体样方,共设置整体样方54个。每个整体样方中设置1个10m×10m的乔木样方,在其角点处设置4个5m×5m的灌木样方;在乔木样方中心及角点处设置5个1m×1m的草本样方。记录乔木层中,距地面1.3m处胸径大于4cm的乔木(含竹类)种名、株数、高度、胸径、冠幅等信息;灌木层(包含胸径小于4cm的乔木小苗及木质藤本)的植物种名、株数、高度、盖度等数据;草本层植物种名、盖度等信息。
2.3 鸟类多样性特征指标
鸟类群落多样性特征指标以α多样性相关指数进行测度[25]。本研究采用了5个测度指标:1)物种多度指标,即单个样地内记录到的鸟类总数,反映了鸟类在样地中的数量;2)物种密度指标,表示单位面积内鸟类数量的多少,按公式(1)计算,单位为ind./hm2;3)物种丰富度指数采用Margalef丰富度指数,表示单个样地内鸟类物种的丰富程度,按公式(2)计算;4)物种多样性指数采用Shannon-Wiener多样性指数,按公式(3)计算;5)物种均匀度指数采用Pielou均匀度指数,按公式(4)计算[25-27]。
式中,D表示鸟类密度;N表示样地中鸟类个体总数;A表示样地面积。
式中,R表示Margalef丰富度指数;S表示样地中鸟类物种数;N表示样地中鸟类个体总数。
式中,H表示Shannon-Wiener多样性指数;S含义同上;Pi表示鸟类i的相对多度。
式中,J表示Pielou均匀度指数;H、S含义同上。
2.4 塘类海绵生境特征因子
研究预判塘类海绵生境鸟类特征受到设计与环境等生境特征因子的影响,故对生境的面积特征、植被特征等设计因子,以及人为干扰环境相关因子进行数据采集与计算(表1)。运用Excel 2016来计算各多样性相关指数。

表1 塘类海绵生境特征因子与取值依据
2.5 数据分析方法
为明确塘类海绵生境中鸟类群落组成特征,划分鸟类集团对于研究鸟类群落间的资源分割及研究鸟类群落结构具有重要意义。本研究依据鸟类食性、活动范围、迁徙习性进行鸟类集团划分[28-30]。鸟类食性分为食虫性、食肉性、杂食性和植食性4类;活动范围分为林冠层、林下灌丛层、草本层、水面4类;迁徙习性即鸟类是否具有迁徙属性。针对上述9个习性属性,对鸟类各习性属性进行赋值,具有某种习性赋值为1,不具有某种习性赋值为0。依据赋值结果,采用K-modes聚类方法划分鸟类集团,该方法划分出的集团具有一定的主观性,当选取不同的汉明距离时可将鸟类群落划分为不同的集团。
研究使用结构方程模型(Structural Equation Modeling,SEM)来量化分析生境特征因子对鸟类群落多度、密度、丰富度指数、多样性指数、均匀度指数的影响。结构方程模型能够分析多变量数据间的因果关系,并且判断各变量之间关系的强度,同时对整体模型进行拟合和判断,有助于全面了解复杂的自然系统,为解决生态学中许多复杂的问题提供了新的视角[31-33]。首先基于已有研究基础与研究假设构建初始模型(图2),该模型中只有观测变量,不包含潜变量。该模型包含了生境特征因子之间,以及生境特征因子与鸟类α多样性指标之间可能存在的影响关系。模型使用最大似然法来估计参数值,故采用卡方检验的P值(Probability level)和卡方自由度比(x2/df)来判断模型的拟合能力。当该模型P值>0.05时,表示模型得到了观测数据的支持;卡方自由度比小于3时,表示模型拟合效果良好[33]。建立初始模型之后,运用Amos模型筛选功能,将不确定的路径排列组合成多个假设模型,用样本数据进行拟合估计,估计出各个模型的拟合度指标值,选取其中P值最大的模型为最优模型。再将Amos中修正指数(modification indices)提示初始模型可能缺少的路径加入模型中,对模型进行修正,最终得到塘类海绵生境特征因子对鸟类α多样性各指数的结构模型方程。
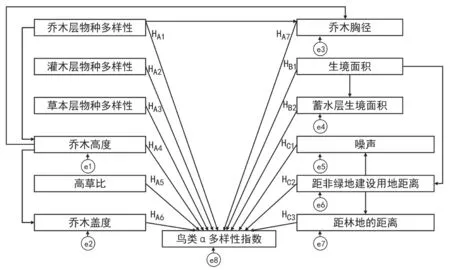
图2 结构方程初始模型(e表示误差变量;HA表示生境植被特征因子对于鸟类α多样性特征有影响;HB表示生境面积特征因子对于鸟类α多样性特征有影响;HC表示生境周边人为环境干扰因子对于鸟类α多样性特征有影响;箭头表示两因素之间可能存在影响关系,作者绘)
此外,由于分类变量无法纳入结构方程模型中,采用t检验,对各鸟类多样性指标在干塘与湿塘生境中是否存在显著性差异进行检验;采用方差分析(ANOVA)对鸟类多样性指标在不同绿地维护强度下是否存在显著性差异进行检验。
3 结果与讨论
3.1 塘类海绵生境鸟类组成与集团划分
塘类海绵生境中总共观察到鸟类64种,其中干塘生境中出现44种,湿塘生境中出现57种。整体调查结果显示,塘类海绵生境整体鸟类多度均值为67.58,密度均值为309.86ind./hm2,丰富度均值为3.38,多样性均值为2.25,均匀度均值为0.85。本研究中塘类海绵生境的鸟类多样性特征,与以往研究中的自然栖息地的鸟类特征相比偏低[34-35],但与部分研究中城市其他绿地生境的鸟类特征相比偏高[36-37],特别是在湿塘生境中的鸟类多度较高。因此,塘类海绵生境能够增加城市生境斑块的多样性,为鸟类捕食、繁衍筑巢提供条件,对生态环境产生积极影响。
结果显示,湿塘生境的各鸟类α多样性指数除密度外均略高于干塘生境,但两者间无显著性差异(p>0.05)。2种生境中鸟类群落多样性无显著差异,因此本文在讨论塘类海绵生境因子对鸟类特征的影响时,未将干塘与湿塘分开探讨。
本研究依据聚类分析结果并结合文献与调研实际情况,将鸟类集团划分为4类:第1鸟类集团为食水生生物、水面活动鸟类(简写为CW);第2鸟类集团为食虫性或植食性、林冠层活动鸟类(简写为IHC);第3鸟类集团为食虫性、林下灌丛活动鸟类(简写为IU);第4鸟类集团为杂食性、林冠层或草本层活动鸟类(简写为OCG)。各鸟类集团及总体鸟类(简写为AB)的多度、密度、丰富度、多样性和均匀度特征见图3。
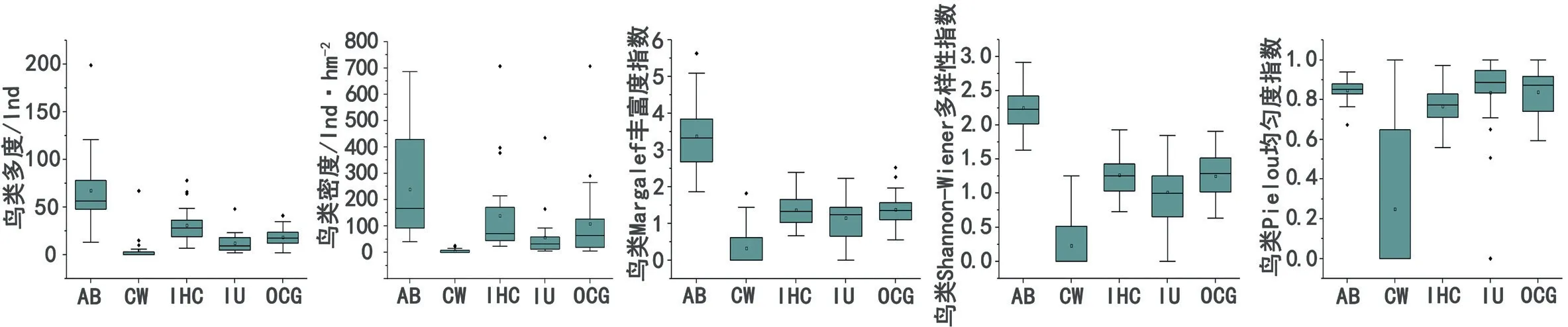
图3 塘类海绵生境中各鸟类集团及总体鸟类的α多样性相关指数特征(AB为总体鸟类;CW为食水生生物,水面活动鸟类集团;IHC为食虫性或植食性,林冠层活动鸟类集团;IU为食虫性,林下灌丛活动鸟类集团;OCG为杂食性,林冠层或草本层活动鸟类集团,作者绘)
IHC鸟类集团的鸟类种类占总数的31%,数量占所记录鸟类总数的46%,且鸟类多度与鸟类Shannon-Wiener多样性指数为4个鸟类集团中最高,由此可知塘类生境为食虫性或植食性,且喜爱为树冠中活动的鸟类创造了良好的觅食与活动环境,对其具有较高吸引力。CW鸟类集团的各α多样性指数相比其他集团偏低,其观测总数为132只,且塘类海绵生境对特定品种水鸟的吸引力较高。
3.2 塘类海绵生境特征因子对鸟类α多样性的影响结果与分析
3.2.1 塘类海绵生境特征因子对鸟类α多样性的影响结果
塘类海绵生境特征因子对鸟类α多样性的结构方程模型在修正后P值均大于0.05,表明模型得到了观测数据支持,其卡方自由度比(x2/df)均小于2,塔克刘易斯指数(TLI)均大于0.9,表明该模型很好地拟合了实测数据。影响分析结果见表2。总体而言,灌木层的物种多样性、蓄水层面积、噪声、据林地的距离对鸟类多度有显著影响;噪声、据林地的距离、距非绿地建设用地距离对鸟类密度有显著影响;乔木胸径、生境面积、据林地的距离对鸟类丰富度有显著影响;乔木层物种多样性、生境面积对鸟类多样性有显著影响;乔木层物种多样性、乔木盖度、生境面积、蓄水层面积对鸟类均匀度有显著影响。
3.2.2 塘类海绵生境面积因子对鸟类α多样性特征的影响分析
生境面积对鸟类物种丰富度、多样性、均匀度均有显著正向的影响。这一结果符合“种-面积关系”理论,即物种数目会随着取样面积的增加而增加,且以往研究也验证了上述结论[38-39]。生境面积的增加满足了一些鸟类对栖息地水平方向活动空间梯度的要求[24],且通常面积越大的生境的栖息地类型也会越多[40]。面积更大的栖息地生境可以为不适应城市环境的鸟类提供更多的资源[41]。本研究中面积为26 738m2的湿塘生境的鸟类丰富度达5.10,多样性达2.92,均匀度达0.88,而面积为971m2的湿塘生境其鸟类丰富度仅1.86,多样性仅1.76,均匀度为0.85。在规划塘类海绵生境时,除考虑满足径流控制指标外,应充分利用城市绿地的汇水洼地区域,营造足够规模的塘类设施,充分发挥该类生境斑块的雨洪管理及生物多样性效益。
蓄水层面积对鸟类多度有显著正向影响,张强等研究表明在一定范围内(小于100hm2),鸟类的种类与数量随着蓄水层面积的扩大而增加[42]。观测发现小䴙鹈、黑水鸡等水鸟偏好于有常水位且水面开阔的生境,在蓄水层面积大于约1hm2时,它们常成群结队出现于湿塘中。但可能由于蓄水层对特定水鸟的吸引作用,使得蓄水层面积对鸟类均匀度产生显著的负向影响。尽管如此,观测发现干湿塘的缓坡边界可以方便林鸟立足饮水,因此在营造塘类海绵生境时,极有必要在塘周边设置绿地缓坡边界,提升蓄水层对林鸟的吸引,同时为水鸟和林鸟提供栖息资源,从而提升塘类海绵生境的鸟类均匀度。
3.2.3 塘类海绵生境植被特征因子对鸟类α多样性特征的影响分析
乔木层物种多样性对鸟类多样性及均匀度有显著的正向影响,这与以往研究结果一致。鸟类大多偏好发达且复杂的乔木生境[43],乔木是鸟类的主要营巢地之一[44],观测发现如珠颈斑鸠、白头鹎等杂食性、林冠层或草本层活动集团中的鸟类会在乔木上筑巢。此外,白鹭、夜鹭等食水生生物,水面活动集团的鸟类会在水域边的大乔木上栖息[45]。观测还发现,由落羽杉这一单一种类乔木构成的样地,鸟类多样性仅有2.05,均匀度为0.80,而在乔木多样性好的样地里,鸟类多样性达2.90,均匀度达0.87。有研究表明,针阔叶混交林能够在城市中支持最高的鸟类多样性[46]。因此在塘类海绵生境乔木层营造时,可将水杉(Metasequoia glyptostroboides)、落羽杉(Taxodium distichum)等耐水湿针叶乔木与枫杨(Pterocarya stenoptera)、楝树(Melia azedarach)等耐水湿阔叶乔木相结合,各配置适宜数量,以形成较高的乔木多样性。乔木胸径对鸟类丰富度为显著负向影响,这与以往部分研究不同[47-48]。推测其原因为:相比种植了大胸径乔木的样地而言,观测发现,种植了紫薇(Lagerstroemia indica)、日本晚樱(Cerasus serrulatavar.lannesiana)等小胸径乔木的样地,能够给鸟类提供更多花粉、种子等资源,从而更能吸引鸟类来取食,导致了上述分析结果。由此说明,相对乔木胸径而言,树种的选择很可能对于提升鸟类丰富度更为关键。
乔木盖度对鸟类均匀度有显著的负向影响,这种影响很微小。推测其原因为,塘类海绵生境中乔木盖度高的地方通常种植纯针叶林或纯阔叶林,高度相似,这样的生境群落的水平与垂直结构都较单一,生态位少,满足不了不同生态位的鸟类需求[49],导致乔木盖度越高的样地反而鸟类均匀度相对越低。以往研究证明,陆生鸟类偏好乔灌组合盖度在40%左右的生境,种植垂直结构单一的高密度乔木或高密度灌木都不利于鸟类觅食与栖息[50]。
灌木层物种多样性对鸟类多度有显著的正向影响。以往研究也表明灌木多样性是影响鸟类多样性的重要因子[51-53]。具有丰富多样性的灌木层能为鸟类提供取食来源。多样性高的灌木层也往往具有丰富的空间层次,可以作为乔木层与地被层的连接通道,为鸟类充当往返地面搜寻食物的踏脚石[53],同时也为鸟类提供了更多躲避捕食者的条件。观测发现在塘类海绵生境中栽植南天竹(Nandina domestica)、小叶女贞(Ligustrum quihoui)等较耐水湿的食源灌木,可增强对食虫性、林下灌丛活动鸟类集团,以及对杂食性、林冠层或草本层活动鸟类集团的吸引力。
值得注意的是,草本层物种多样性对鸟类α多样性没有显著影响,这与之前的部分研究结论存在差异。主要原因推测是,目前塘类海绵样地的草本植物配置较为相似且单一,缺乏足够的样本差异使草本多样性指标对鸟类特征产生影响。以往研究表明草本物种多样性能吸引更多的昆虫及无脊椎动物[6,14],从而为鸟类提供食物。观测发现,湿塘周边种植的挺水植物,如东方香蒲(Typha orientalis)、梭鱼草(Pontederia cordata)、芦苇(Phragmites australis)与灌木丛搭配,可为食水生生物、水面活动集团鸟类提供躲藏与筑巢的地方,所以有必要在蓄水层周边配置梭鱼草、芦苇之类的植物以吸引鸟类。
3.2.4 塘类海绵生境周边人为干扰环境因子对鸟类α多样性的影响分析
噪声对鸟类多度与密度有显著的正向影响。塘类海绵生境中的部分鸟类如白头鹎、珠颈斑鸠、八哥、麻雀等,其长期暴露于人为干扰环境中,表现出适应性,而上述鸟类在观测数量上具有优势,且往往成群结队大量出现。以往研究也表明,长时间暴露在人为干扰环境中的鸟类,其对噪声环境的耐受度会增强,且人类活动给杂食性鸟类带来食物残渣,从而增加其数量[50,54],使得在城市区域中的鸟类多度会随人为干扰程度的加大反而增加。
方差分析表明,不同绿地维护强度间的各鸟类多样性特征指标无显著差异(p>0.05),即周围绿地维护强度改变不会对塘类海绵设施中的鸟类多样性产生显著影响。此外,距非绿地建设用地距离对鸟类密度有显著的负向影响,即距非绿地建设用地距离越近的样地中鸟类密度越大。在高度城市化的环境中鸟类往往缺乏栖息地,塘类海绵生境为鸟类提供了可供停歇觅食饮水的绿地斑块,吸引了大量城市中的鸟类停留,因此周边环境有大量建筑与公路等建设用地的样地中的鸟类往往更加聚集[55-56],这说明了在高密度城市环境中营造塘类生境的重要性。
距林地的距离负向地影响了鸟类多度、密度与丰富度,即离林地的距离越近时鸟类的多度、密度与丰富度会上升。大面积的林地可为鸟类供给更多食物资源[57],塘类设施可为鸟类提供水源,林地与干湿塘结合可为鸟类营造较好的生存环境。在考虑塘类海绵生境营造时,其布局位置多依据汇水区汇流分析确定,同时可尽量考虑靠近林地的地点。
4 结论
本研究聚焦海绵城市试点地区的塘类生境,探讨其设计因子和环境因子对生境中鸟类特征的影响。经过4个季节,塘类海绵生境中总共观察到鸟类64种。塘类海绵生境中,面积特征因子、植被特征因子、人为干扰环境因子均对鸟类α多样性特征有显著影响。蓄水层面积、灌木层物种多样性、噪声对鸟类多度有显著正向影响,距林地的距离对鸟类多度有显著负向影响;噪声对鸟类密度有显著正向影响,距非绿地建设用地距离、距林地的距离对鸟类密度有显著负向影响;生境面积对鸟类丰富度有显著正向影响,林地距离、乔木胸径对鸟类丰富度有显著负向影响;生境面积、乔木层物种多样性对鸟类多样性有显著正向影响;生境面积与乔木层物种多样性对鸟类均匀度也有显著正向影响,蓄水层面积、乔木盖度对鸟类均匀度有显著负向影响。
在营造塘类海绵生境时,除满足雨洪管理指标要求外,规划选址应靠近林地,尽可能充分利用汇水洼地区域,或通过挖填方营造充足的生境斑块及蓄水层面积(建议大于1hm2)。塘周边营建具有丰富多样性,且具有食源植物的乔木层,强调针叶树与阔叶树混合种植,避免乔木盖度过高,营造疏林生境。同时,种植多样性高、层次丰富,具有食源植物的灌木层,注重生境异质性。塘周边营造缓和边坡绿地,种植挺水草本植物。上述营造方法有利于提升塘类海绵生境的鸟类多样性特征。
本研究观测时间近1年,后续研究可增加观测时间与频次以提升分析结果的准确性。此外,除了本文研究的生境设计因子与环境因子之外,后续研究还可以探索水量、下垫面覆盖类型、外来入侵植物、植物总量等因素对塘类海绵设施中鸟类特征的影响[50,58]。整体而言,本研究内容对于探索城市绿地中小微生境的生物多样性效益,促进海绵城市精细化营建具有积极重要价值。
致谢:感谢重庆悦来投资集团有限公司规划设计部部长魏映彦、注册城市规划师申亚,重庆市观鸟协会会长危骞,西南大学园艺园林学院硕士生张蕊、张继升、晁欣然、徐颖在数据收集过程中提供的帮助;感谢百时美施贵宝公司的金时强博士为数据分析问题提供解答;感谢京都府立大学福井亘教授、华侨大学高江菡副教授为鸟类观测方法提供建议。
