农田边界和有机生产对稻田节肢动物天敌和害虫多样性的影响*
2023-12-21李佳宁范顺祥刘欣宇刘云慧
孟 璇 ,李佳宁 ,范顺祥 ,刘欣宇 ,刘云慧**
(1.中国农业大学有机循环研究院(苏州) 苏州 215000;2.中国农业大学资源与环境学院生物多样性与有机农业北京市重点实验室 北京 100193;3.江苏昆山果蔬科技小院 昆山 215300)
过去几十年间,为了满足不断增加的粮食需求,农业集约化程度不断增加,逐渐成为全球生物多样性丧失的重要因素[1]。农业化学品投入增长、土地集约化程度加深导致了生态环境恶化、半自然生境与自然栖息地丧失、景观均质化和破碎化等一系列问题[2],进一步影响了农业景观中生物多样性保护和相关生态系统服务功能的发挥,最终可能威胁到粮食生产的可持续性和人类福祉[3]。农田中天敌和害虫种群的发生情况是生物多样性和生态系统服务功能状况的重要指针。为了更好地保障粮食安全,加快推进农业绿色可持续发展,认识了解农田景观影响害虫天敌多样性和分布的因素,是制定农田景观生物多样性保护和绿色生产措施的基础[4]。
有机生产作为一种减少生产过度集约性并采用生物友好方法的生产方式,具有显著的生态优势。强调生产过程中不采用基因工程获得的生物及其产物,不使用化学合成的农药、化肥、生长调节剂、饲料添加剂等物质,同时重视周围景观中自然半自然生境对农田生产的调节和支持作用,强调生态系统的平衡,通过种植不同的植物,吸引有益的昆虫和动物[5]。农田景观中的半自然生境包括田埂、花带、稻田边界、绿篱、林地、草地等相对不受干扰的区域,为许多生物提供了食物资源和生存环境[6],这对农田景观中生物多样性保护和相关生态系统服务的维持具有重要意义。增加农田中植物多样性、植被覆盖面积能够通过直接增加天敌食物资源(如花粉、花蜜、种子)或间接增加猎物,增加天敌数量,促进田间害虫的控制[7-8]。欧盟农业环境计划措施[9]鼓励各成员国重视农田半自然生境,尤其是开花植物带的建设[10-12]。虽然国内开展了不少关于有机生产方式对生物多样性影响的研究[13-15],但鲜有研究评估开花植物带建设对农田生物多样性、天敌和害虫的影响[16-17],尤其是缺乏对有机栽培水稻(Oryza sativa)种植系统开花植物带建设对生物多样性以及天敌和害虫的影响研究。此外,半自然生境的节肢动物会随着作物生产情况变化在农田和半自然生境迁移移动[11]。在害虫发生早期,半自然生境中的自然天敌可能向作物生境扩散,从而抑制了害虫种群的增长[18]。但是,受节肢动物自身扩散能力的限制,它们从半自然生境向农田内部扩散存在有效距离[19]。因此,本研究将深入了解稻田天敌和害虫随边界半自然生境距离的变化,从而为合理规划半自然生境在农田景观中的配置、促进农田生物害虫防治提供参考。
针对上述问题,本研究在江苏省昆山市对不同稻田边界及生产模式下的稻田主要生物类群(节肢动物)展开调查,研究水稻种植系统有机生产管理以及开花植物建设对稻田天敌、害虫多样性及二者相对数量的影响,为水稻绿色生产、稻田生物多样性和害虫生物控制服务提升提供科学依据。研究将重点回答如下科学问题: 1)在有机、常规生产模式下,稻田中天敌维持和害虫发生是否不同? 2)有无开花植物带边界的稻田中天敌维持和害虫发生是否存在不同,随着边界距离增加,天敌维持和害虫发生有何变化? 3)稻田边界地表覆盖植被状况是否影响稻田内及边界田埂天敌、害虫多样性及其相对多度?
1 研究区概况和研究方法
1.1 研究区概况
研究地点位于江苏省昆山市巴城悦丰岛水稻基地(31°39′49″N,120°85′57″E)。研究区属于亚热带季风湿润气候,四季分明,阳光充沛,气候温和,无霜期长。2022 年7-10 月有效积温为1891 ℃、月平均气温为25.36 ℃,降水量为1187 mm。镇域面积157 hm2。水稻基地被一条河流划分成不同的区块。大多稻田田埂上有自然演替的草花,农场外围有林带和花带绿化,大片花带主要集中在东北侧。稻田主要种植的水稻品种是‘南粳46’。土壤类型为黄土。取样区地理位置详见图1。
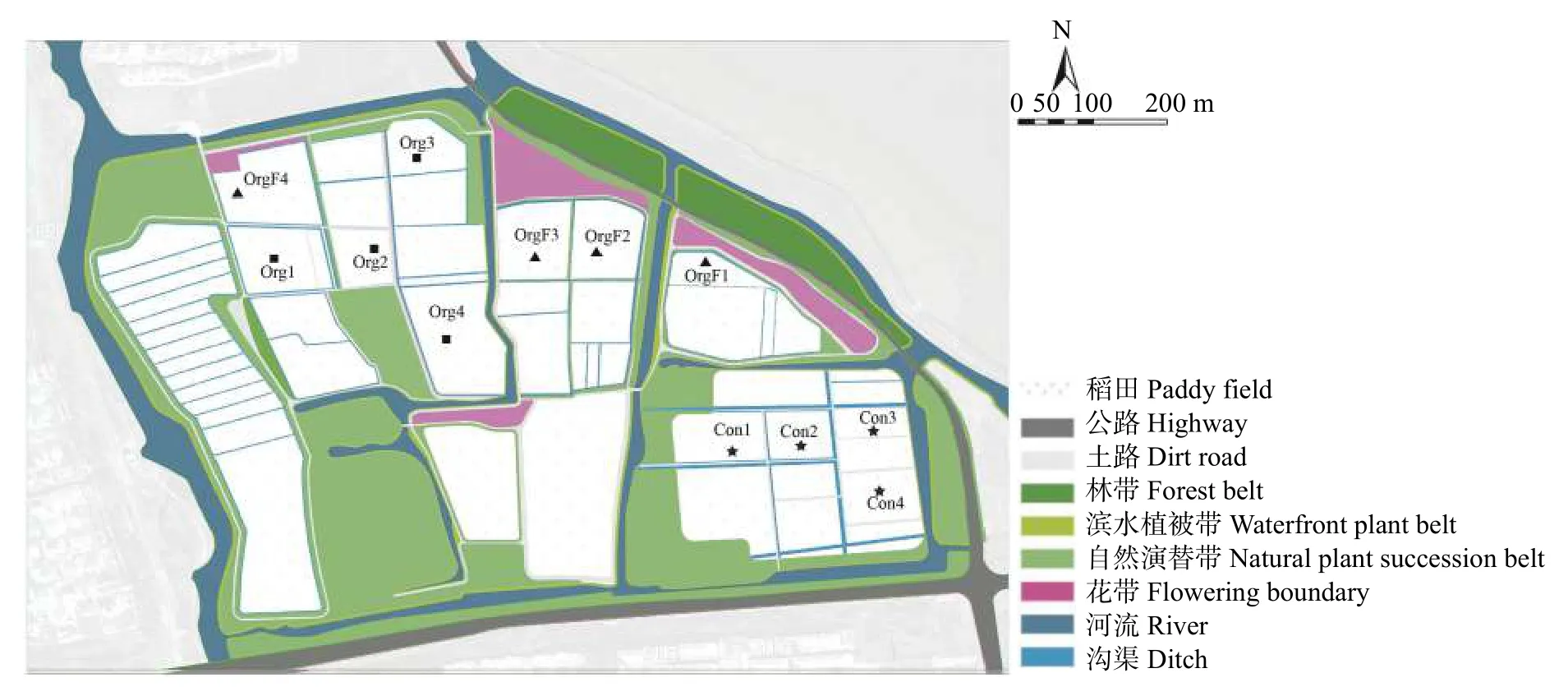
图1 昆山市巴城悦丰岛水稻基地采样稻田分布图Fig.1 Distribution of the sampling paddy fields in Yuefeng Island in Bacheng Town in Kunshan City
研究区内,有机稻田使用生物农药和人工除草进行病虫害防治,传统生产系统则依赖于杀虫剂。在水稻收获前,所有稻田播种油菜(Brassica napus)和苕子(Vicia dasycarpa)作为冬季覆盖作物,在次年播种绿肥。有机稻田和常规稻田的管理信息详见表1。
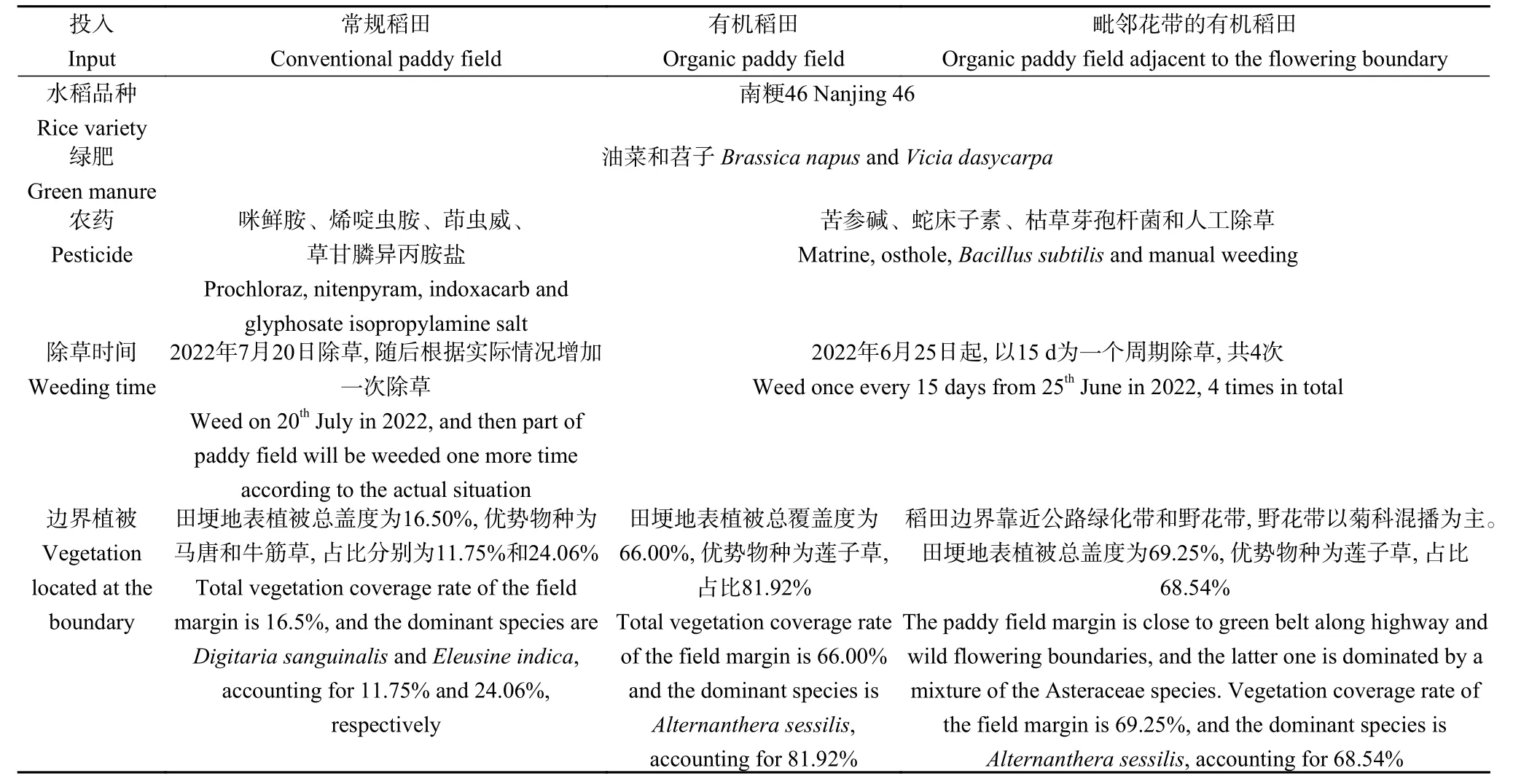
表1 有机稻田和常规稻田管理信息Table 1 Management information of organic and conventional paddy fields
1.2 取样点设计
2022 年7-10 月,在研究区选取常规稻田(Con)、有机稻田(Org)和毗邻花带的有机稻田(OrgF) 作为3 个处理,每个处理4 个重复,总计12 块稻田进行节肢动物生物多样性调查。在每个样地地块设置3 条样带,第1 条样带是稻田作物边界(即样带①),随后在平行边界且距离边界5 m、20 m 的农田内部设置第2、3 条样带(即样带②和样带③),每条样带上设置3 个取样样方,分别位于样带中心及距离中心两侧各3 m 的位置。研究区取样点位置如图2 所示。
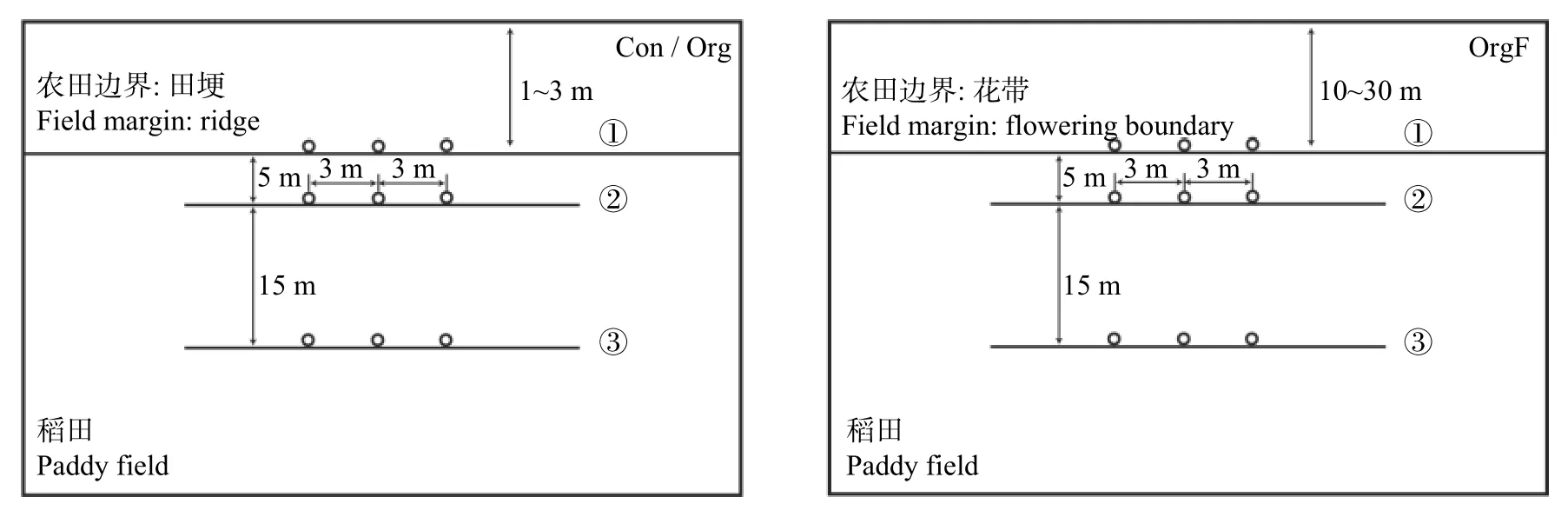
图2 不同处理稻田中节肢动物取样样方位置示意图Fig.2 Schematic diagrams of the location of sampling plots of arthropod in paddy fields of different treatments
1.3 节肢动物调查
采用吸虫器法于2022 年7-10 月对稻田节肢动物进行取样调查。以底部直径为41 cm、高为48 cm、口径为51 cm 的圆桶(70 L)作为取样样方,取样时将桶底靠在水稻田表面,用纱网反套在桶口边缘并进行固定,防止取样时虫子飞出样方范围。将尼龙网袋反套在汽油式吸虫器(Oleo Mac BV300)上,然后在规定的样方面积范围内,沿着水稻植株的垂直方向,自顶部至底部有序地采样,每个样方的采集过程为2 min,以确保高质量的科学数据采集。每个样方的标本单独收集于尼龙网袋中,随后将其放入装有75%酒精的塑料瓶中,并标注时间、地点、编号及天气等,带回实验室进行分类鉴定标号,并保存于离心管中。每次取样间隔半个月左右,共取样6 次。将每个样地重复3 条样带捕获的节肢动物进行合并,并作为该样地的数据,共计12 块样地;将每个样带上3 个样方的数据合并分别作为边界、距离边界5 m、20 m 样地的数据,共计36 条样带。将节肢动物分为天敌和害虫两类。其中,稻田主要天敌蜘蛛(Araneae)标本送请分类专家鉴定到种水平(包括蜘蛛的种、年龄、性别),其他节肢动物鉴定到科水平。
1.4 植被调查
在节肢动物取样期间,在每个稻田边界田埂内随机选取3 个0.5 m×0.5 m 的样方,记录样方中植物的物种和盖度,并将3 个样方的植物物种和盖度的均值作为每个稻田样地的植被数据。
1.5 数据分析
统计各处理和样带样方中天敌和害虫的物种数和多度。为全面评估本次样本量的饱和情况,将各样带天敌和害虫数据汇总分析,在PAST 4.10 中构建稀释曲线,若曲线趋于平缓,说明样本量充足,否则需进一步增加样本量。为分析不同生产管理和稻田边界处理的生物多样性差异,在PAST 4.10 中对其进行α 多样性指数的计算,随后在IBM SPSS Statistics 25 中对天敌和害虫的多度和丰富度进行正态性及方差齐性检验,再采用单因子方差分析[20-22]或Kruskal-Wallis (K-W)检验[23]对多样性指数进行显著性检验。
为分析生产管理和稻田边界处理节肢动物的群落组成差异,采用基于CNESS (Normalized Expected Species Shared)相似系数的非度量多维尺度法(NMDS) 分析比较稻田边界和不同生产管理的群落结构的相似性;并通过胁强系数(stress)衡量结果的可信度,当模型拟合结果stress 值小于 0.2 时,则差异性在可接受范围内,结果总体可信[24]。为进一步研究群落组成之间的差异,采用非参数多元方差分析进行比较,当P<0.05 时,认为不同处理群落物种组成存在显著差异[25]。为分析边界植被对稻田内及边界田埂节肢动物总多度和丰富度的影响,采用一般线性模型研究节肢动物对半自然生境的田埂响应。将天敌和害虫的丰富度、多度纳入模型中作为响应变量,将地表植被的盖度和丰富度纳入模型中作为解释变量,这些模型变量符合正态分布。以上所有分析均在R 4.13[26]中进行,主要采用vegan、ggplot2、plyr 包分析。
此外,通过对稻田节肢动物天敌和害虫多度比的对比,分析不同稻田边界和生产方式下稻田天敌对害虫服务控制的潜力。
2 结果与分析
2.1 节肢动物基本组成情况
在12 块稻田中,共捕获节肢动物9531 头,分属50 个科。其中,天敌类群有28 个科2653 头,个体数的优势科有皿蛛科(Linyphiidae,15.23%)、跳蛛科(Salticidae,12.02%)、狼蛛科(Lycosidae,25.52%)。害虫类群有18 个科3971 头,个体数的优势科为叶蝉科(Cicadellidae,84.01%)。在所捕获的所有天敌中,共捕获蜘蛛2253 头,分属14 个科41 个种。蜘蛛约占天敌类群的85%,为天敌的优势物种。其中,物种数的优势科为皿蛛科、狼蛛科和跳蛛科,分别占蜘蛛物种数的14.63%、12.20%和17.07%。个体数的优势物种为雾豹蛛(Pardosa nebulosa),占捕获蜘蛛总数的13.94%。
2.2 农田边界和生产管理对稻田节肢动物生物多样性的影响
2.2.1 节肢动物中天敌和害虫的稀释曲线
节肢动物中天敌和害虫的样本稀释曲线趋于平缓,表明样本量充足,其多样性指标达到稳定;不同稻田边界和生产管理方式下稀释曲线显示(图3),OrgF 处理在相同天敌个体数条件下可能拥有更多天敌物种数,且OrgF 与Org 在最大共同物种数时,误差线内无交叠,表明OrgF 与有机Org 存在显著差异;害虫稀释曲线结果显示(图3),Con 与Org 在最大共同物种数时,误差线内无交叠,表明Con 较Org 拥有更多害虫物种数,且存在显著差异。
2.2.2 节肢动物中天敌和害虫的α 多样性分析
单因子方差分析以及 K-W 检验的结果显示(图4),不同稻田边界和生产管理方式下稻田节肢动物中天敌丰富度 (Chi2=8.439,P=0.015)和多度 (F=7.412,P=0.013)及害虫丰富度 (F=4.748,P=0.039)和多度(F=17.910,P=0.001)均差异显著。
OrgF 平均捕获约21 种天敌,显著高于Con (15)与Org (19);OrgF 平均捕获约292 头天敌,显著高于Con (136)。OrgF 平均捕获13 种害虫,显著高于Org(10);Org 平均可捕获506 头害虫,OrgF 平均捕获387 头害虫,两种有机稻田均显著高于Con (100) (图4)。
2.2.3 节肢动物中天敌和害虫的β 多样性分析
对不同处理稻田节肢动物群落结构分析结果显示(图5),天敌和害虫的stress 值均小于0.2,说明结果具有较好的可信度。P值均小于0.05,说明不同处理稻田内部生物组成均差异显著。就天敌而言,它们在两种有机稻田中群落组成较相似,但区别于Con,说明有机管理的稻田中天敌群落结构不同于Con。害虫在3 种稻田中均有重叠,说明几种处理模式下稻田害虫群落结构相似。

图5 基于非度量多维尺度分析(NMDS)与群落相似度分析的不同处理稻田节肢动物群落组成对比Fig.5 Comparison of community composition of arthropods in paddy fields under different treatments based on non-linear multidimensional scaling (NMDS) and community similarity analysis
2.3 农田边界对稻田内部不同距离节肢动物多样性的影响
2.3.1 节肢动物中天敌和害虫的α 多样性
方差分析结果显示(图6),Org 处理边界0 m 处天敌丰富度显著低于距离边界5 m 和20 m 的稻田内部(P<0.05);OrgF 处理天敌丰富度在5 m 时显著高于20 m,天敌多度在5 m 时显著高于0 m;OrgF 处理在0 m 时,天敌丰富度、多度均高于Con 和Org。

图6 距农田边界不同距离的不同处理稻田内部采样样方的节肢动物多样性指数Fig.6 Arthropod diversity indexes in sampling plots with different distances from the field margin in paddy fields under different treatments
就害虫多样性而言(图6),3 个处理下农田边界与稻田中害虫多度均差异显著,Con 处理农田边界害虫多度最高,5 m 处最低;Org 和OrgF 处理下,害虫多度在距离农田边界20 m 处的稻田内部显著高于边界;OrgF 处理害虫丰富度随着样方与边界距离的增加而减小,害虫多度随着样方与边界距离的增加而增加。
2.3.2 节肢动物中天敌和害虫的β 多样性分析
图7 结果显示,距农田边界不同距离的3 种处理稻田内部天敌的stress 值均小于0.2,说明结果具有较好的可信度;P值均小于0.001,说明与农田边界距离不同的稻田内部天敌群落组成差异显著;距离农田边界5 m 和20 m 处的稻田内部天敌物种有交叉,说明其群落结构较相似,但不同于边界的天敌物种。

图7 距农田边界不同距离的不同处理稻田内部节肢动物中天敌群落的非度量多维度分析(NMDS)Fig.7 Non-linear multi-dimensional scaling (NMDS) based on natural enemies from arthropods in sampling plots with different distances from the field margin in paddy fields under different treatments
图8 中的结果显示,距农田边界不同距离的3 种处理稻田内部害虫的stress 值均小于0.2,说明结果具有较好的可信度;P值均小于0.001,说明与农田边界距离不同的稻田内部害虫群落组成差异显著;Con和OrgF 处理中,距离农田边界5 m 和20 m 的稻田内部害虫物种有交叉,说明其群落结构更为相似,但显著不同于农田边界处;Org 处理在与半自然田埂不同距离处的稻田内部害虫物种均有重叠,说明边界与农田内部害虫群落结构较为相似。
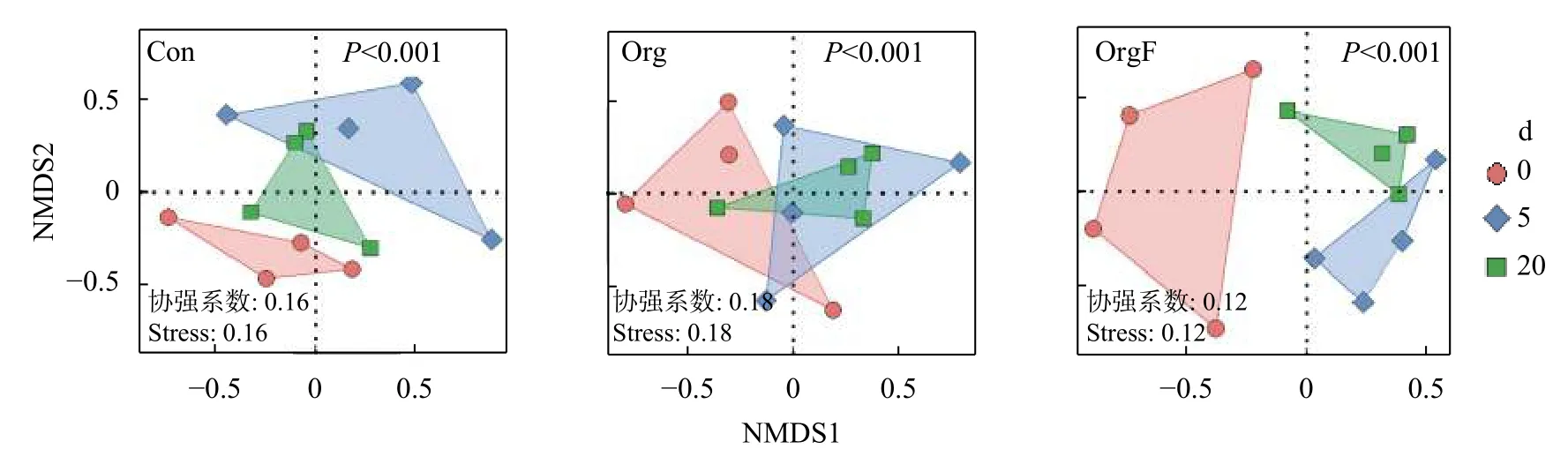
图8 距农田边界不同距离的不同处理稻田内部节肢动物中害虫群落的非度量多维度分析(NMDS)Fig.8 Non-linear multi-dimensional scaling (NMDS) based on pests from arthropods in sampling plots in paddy fields with different distances from the field margin in paddy fields under different treatments
2.4 农田边界地表覆盖植被对稻田内及田埂节肢动物多样性的影响
对稻田边界地表覆盖植被的调查结果显示(表2),稻田边界的植被盖度与丰富度在Con 处理最低,在OrgF 处理最高。一般线性回归分析结果显示天敌丰富度、多度及害虫多度与稻田边界地表覆盖植被盖度呈显著正相关(图9),OrgF 处理受到田埂边界植被盖度影响最大,能维持节肢动物较高的多样性。

表2 不同处理稻田边界地表覆盖植被基础数据Table 2 Basic data of the surface cover vegetation at field margin of paddy fields under different treatments
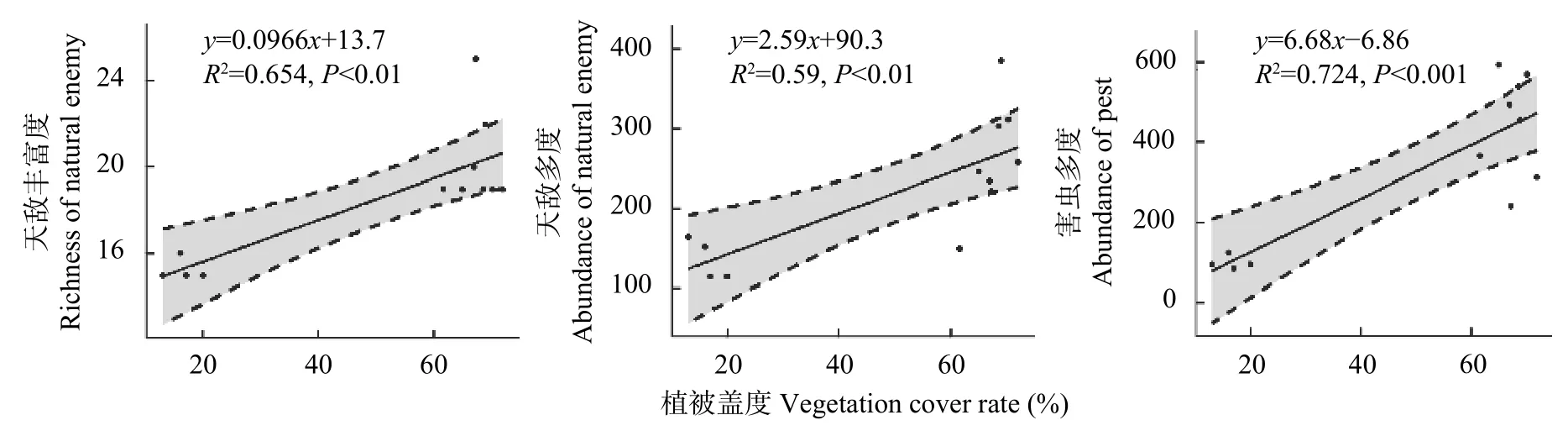
图9 稻田边界田埂地表植被盖度与稻田内部节肢动物多样性的关系Fig.9 Relationship between vegetation cover rate at the field margin and arthropod diversity in paddy fields
2.5 农田边界对相邻稻田节肢动物分布及天敌和害虫多度比的影响
单因子方差分析结果显示(图10),不同稻田边界和生产管理方式下稻田节肢动物中天敌和害虫多度比差异显著(F=32.388,P<0.001)。其中,Con 处理下节肢动物中天敌和害虫多度比最高,其次为OrgF,Org 处理天敌和害虫多度比最低;农田边界和与边界不同距离的稻田内部节肢动物中天敌和害虫多度比差异显著(F=27.362,P<0.001),其中稻田边界和距离边界5 m 处的稻田内部节肢动物中天敌和害虫多度比较高,且显著高于距离边界20 m 稻田处节肢动物中天敌和害虫多度比。
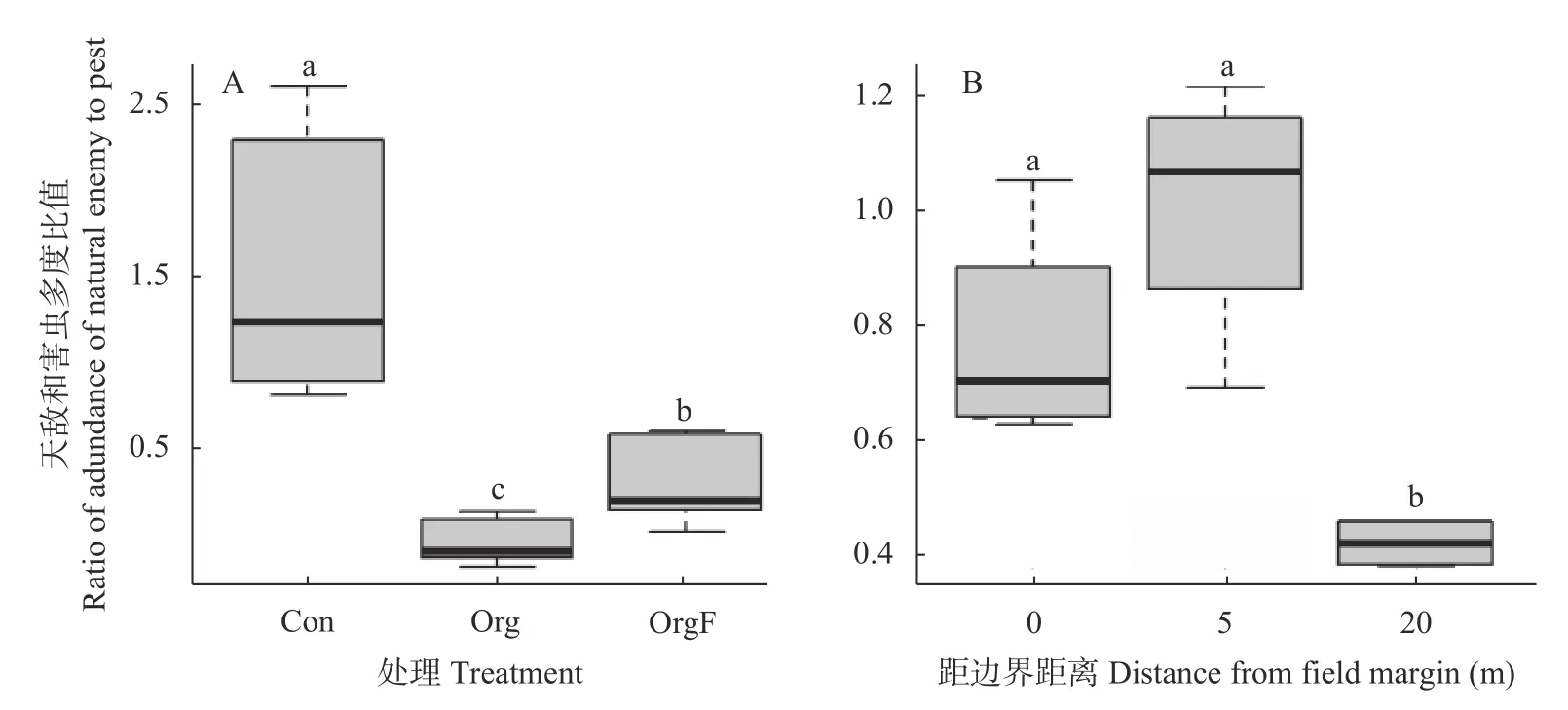
图10 不同处理稻田(A)及与农田边界不同距离稻田内部(B)节肢动物中天敌和害虫多度比Fig.10 Ratio of abundance of natural enemies to pests from arthropods in paddy fields under different treatments (A) and paddy fields with different distances from field margin (B)
3 讨论
3.1 稻田边界和生产管理对稻田节肢动物中天敌和害虫的分布及其多度比的影响
稻田生产管理措施会影响稻田节肢动物的密度、群落结构及物种丰富度等[27]。常规稻田由于集约化管理,大量使用化肥农药,从而导致生物多样性减少[28],而有机耕作方式避免了化学农药的使用,转而应用生物农药,维持了节肢动物中天敌群落的多样性[29]。本研究中,两种有机稻田天敌丰富度均显著高于常规稻田,说明从生物多样性保护角度,发展有机农业是有益的,这与早期研究结果一致[30-31]。尽管如此,值得强调的是,天敌多样性的增加未必带来更有效的害虫防治效果[31]。我们的研究也发现,常规耕作方式下,尽管保有的天敌物种数量和丰富度更少,但其天敌与害虫多度比却显著高于两种有机稻田,这可能是归因于常规稻田使用的化学杀虫剂具有广谱性[32-33],大幅降低了害虫多样性。相比之下,有机稻田由于未使用杀虫效果强烈的化学杀虫剂,其施用的生物农药如苦参碱主要针对稻飞虱(Nilaparvata lugens)和二化螟(Chilo suppressalis),蛇床子素主要针对纹枯病,枯草芽孢杆菌(Bacillus subtilis)主要针对稻瘟病,虽然这些病虫害得到了控制,但其他害虫却因为不受影响,多样性很高,因此本研究中害虫的优势物种是叶蝉科,这解释了害虫数量未快速下降的现象[34]。这一发现显示有机生产虽然有益于生物多样性的保护,但天敌多样性与害虫生物控制服务之间的关系尚不确定,需要进一步探究如何实现多样性保护与害虫生物控制服务协同提升的方法和途径,例如需要探究如何有选择性地种植功能植物,实现对目标天敌的精准保育或害虫的趋避或防治,在保护生物多样性的同时,提升生态系统服务功能[35]。
本研究中,毗邻不同农田边界的稻田节肢动物中的天敌和害虫多样性差异显著。具体而言,在毗邻花带的有机稻田中,天敌物种的丰富度显著高于常规稻田和有机稻田;天敌多度显著高于常规稻田。这是因为在农田周边播种野花带和杂草带,能有效增加农田边界的结构复杂性和物种多样性,增加农田生物栖息地和食物资源,从而为节肢动物的繁衍和生产提供有益的生存环境,有利于节肢动物多样性维持[36],也有利于形成更加复杂的食物网,进而增加系统的稳定性,减少病虫害暴发[7]。此外,本研究中,常规稻田和有机稻田均采用了与油菜、苕子的轮作方式,在水稻生长季节结束后进行冬季绿肥的覆盖,这可为天敌越冬提供舒适的栖息环境,提高了它们的生存率[16],这种农田管理策略能够增加稻田内资源的可利用性,为天敌提供更多的食物来源,从而提高了其生态系统服务功能,确保来年稻田生物多样性的提高。因此,在农田管理中,综合采用田间生态管理措施和在农田边界布设开花植物带两种方式,能够更好地增强农田生态系统的稳定性,推动水稻生产的绿色可持续发展。
3.2 稻田边界对距离边界不同距离的稻田内部节肢动物分布的影响
本研究中,与边界不同距离的常规稻田节肢动物中天敌多度和丰富度无显著差异,造成这一现象的原因可能是常规稻田长期的种植管理模式使稻田中主要是一些数量较多、分布较广泛的物种[37-38],也缺乏可能迁入的物种源。此外,与农田边界不同距离的有机稻田节肢动物中天敌仅丰富度显著,而多度无显著差异,但在毗邻花带的有机稻田处理中,与稻田边界距离不同的稻田内部节肢动物中天敌多度与丰富度均差异显著,其中天敌丰富度在5 m 时显著高于20 m,天敌多度在5 m 时显著高于0 m,这可能是由于稻田边界开花植物改变了稻田边界的生境条件,能为捕食者提供更充足的水分、食物、隐蔽和繁衍场所等基本环境条件[39],这种多样化的资源分布使稻田边界可以适应不同种类天敌的生存,进而捕食者出现的概率更高。这种环境的改变也会提高稻田边界捕食者的种间竞争和捕食压力,从而导致种群生态位的重叠[40]。捕食者生态位的高度重叠带来的生存压力可能会迫使捕食者分化出不同的资源利用策略,以避免直接竞争。例如,发展出不同的捕食策略、生境选择,因此稻田边界部分捕食者会选择向稻田内部高度移动,这种迁徙行为使捕食者生态位分散开来[10]。同时,害虫种群可能受到稻田边界捕食者密度的影响,导致害虫会避开边界,而稻田内部作为一个生态系统,可以为害虫提供良好的生存环境。水稻植株能为害虫提供更加充足的生存空间和食物资源,所以更加有利于害虫生存。因此,在两种有机稻田内部,天敌的丰富度和多度在与稻田边界距离较近(5 m)处较高,而害虫的多度在与边界距离较远(20 m)处较高,这也可以进一步解释基于非度量多维尺度分析得出的结论,即不同距离样带的群落之间的相似度较低[41]。由此可见,减小农田面积也有利于生物多样性的提高[42]。
3.3 边界地表覆盖植被对节肢动物发生的影响
本研究中,天敌的丰富度和多度以及害虫的多度均与稻田边界植被盖度呈显著正相关。这再次证明了稻田边界保留自然植被有利于形成多样化的生境,从而增加食物链的层次,有助于建立更稳定的食物网,进而促进稳定的、广谱的捕食者种群的形成,尤其可以为天敌提供更加舒适的栖息环境和丰富的食物资源[43],促进天敌的生物多样性。但多样的植物生境也可以同时为害虫提供生存环境[44],因此可能在促进天敌的同时,也增加了害虫多样性。有研究表明,不同植物物种对天敌、害虫的吸引或者趋避作用是不同的,这取决于植物的特性和生命周期。利用香根草(Chrysopogon zizanioides)对害虫的诱集作用及糖蜜草(Melinis minutifora)的趋避作用,可以实现螟蛾科(Pyralidae)害虫的防治[45]。鸭茅(Dactylis glomerata)、绒毛草(Holcus lanatus)、梯牧草(Phleum pratense)等一些禾本科的丛生禾草能够为甲虫提供潮湿荫蔽的理想环境,让它们在稻田中建立栖息地,这能实现天敌对蚜虫等害虫更好的控制作用[46]。夏季开花的霍香蓟(Ageratum conyzoides)、一点红(Emilia sonchifolia)等植物进入盛花期,可富集大量瓢虫(Curinus coeruleus)、蜘蛛目(Araneae)、赤眼蜂(Trichogrammatid)等天敌,它们有利于稻纵卷叶螟(Cnaphalocrocis medinalis)、稻飞虱的控制[46]。并非所有开花植物种类和其高密度的种植都能确保生物多样性的显著提升,如一年蓬(Erigeron annuus)是一种对害虫吸引力大于天敌的植物[47]。另一方面,本研究中,天敌和害虫的多样性指数与稻田边界植被丰富度无显著关系。这可能是因为本研究调查的稻田边界植被多是一些自然演替的杂草,这些植物物种对于天敌并没有针对性的多样性提升。综上,稻田边界的植被状况与天敌、害虫的数量之间存在着复杂的关系。因此,在农田边界的植被建设管理中,从多样性保护的角度应适当保留一定量的田埂和杂草[48],并且应当更好地了解不同植物和有益生物之间的相互关系,根据具体的环境和植物特性综合利用生物的种间关系,发挥不同植物的趋避、诱集效果,建立复杂的食物网,促进天敌和寄生虫种群的建立,才能更好地控制害虫种群,制定更有效的农田生态系统管理策略。
4 结论
稻田节肢动物多样性和群落组成受稻田边界和生产管理的影响,农田边界建设花带有利于稻田节肢动物中天敌多样性的提升,稻田边界植被盖度增加也有利于稻田节肢动物中天敌多样性维持;随与农田边界距离的增加,稻田中天敌和害虫多度比降低。综上,地块内进行有机生产管理,同时在农田边界种植功能植物带有利于稻田生物害虫控制服务的提升,促进稻田绿色生产。同时,距离边界5 m 处稻田天敌生物多样性最佳,因此保持较小的农田地块及较高密度的农田边界带,更利于提高稻田生态系统对捕食性天敌维持的潜力,最终增加天敌多样性,促进自然害虫控制。
致谢感谢昆山悦丰岛有机农场朱秀妹、汤耀中等人在实验场地、取样调查方面给予的支持和帮助!
