生态旅游型乡村的乡土景观植物遗传多样性*
2023-12-21吴秀忠陈大华范开放唐建路米小琴王义强
冯 致 ,姚 志 ,孙 全 ,李 萌 ,吴秀忠 ,陈大华 ,蒋 龙,范开放,唐建路,李 锴,米小琴,于 咏,王义强**
(1.中南林业科技大学林业生物技术湖南省重点实验室 长沙 410004;2.岳麓山实验室碳汇林品种创制中心 长沙 410004;3.湖南省湘西自治州林业局 吉首 416000;4.湖南省湘西自治州森林生态实验站 永顺 416700)
近年来,我国越来越关注乡村绿色和生态发展,强调人与自然和谐共生,各项政策的重点是生态环境保护和修复、生物多样性保护、生态旅游和森林康养绿色产业发展等领域[1]。国家农业农村部科教司发布了中国美丽乡村建设十大模式,分别为产业发展型、生态保护型、城郊集约型、社会综治型、文化传承型、渔业开发型、草原牧场型、环境整治型、休闲旅游型和高效农业型[2]。为更好地促进乡村可持续发展,美国、加拿大、日本等发达国家正大力推进乡村生态旅游产业的发展进而推动地区经济、生态和文化的发展。乡村生态旅游是由“乡村旅游”和“生态旅游”两个概念发展而来,两者之间既有联系又相互区别。乡村旅游是以乡村地区为界,通过乡村自然和人文景观特色,满足旅游者在此休闲、观光、娱乐及学习等多种需求[3-4]。生态旅游以学习、欣赏地区自然景色和野生动植物为目的,前往相对无干扰和无污染地区旅行[5]。乡村生态旅游是以乡村为背景、以农业产业为支撑、注重生态环境保护的旅游形式[4]。10 种美丽乡村建设模式中生态保护型和休闲旅游型农村建设模式与乡村生态旅游开发的理念相符合。两种乡村模式的营建能保护地方乡村生态与文化,同时保证经济可持续发展。在两种乡村模式建设过程中,具有本土地缘性、景观观赏性、环境保护性、文化承载性等特征的乡土景观植物是美丽乡村营造的关键因素之一,其与农田、溪流、祠堂、民居等乡村要素共同构成乡村生态景观。然而在人为经济生产活动和自然环境保护修复等多种因子影响下,乡土景观植物的种群数量与种群结构会发生变化,进而改变乡土景观植物群体的遗传多样性水平。遗传多样性水平低的乡土景观植物无法持续、稳定地在美丽乡村营建中提供支撑。
乡土景观植物是指生长在乡村地区自然生态环境中的本土景观植物或在本地经过长期引种驯化的景观植物[6]。相较于非乡土植物,乡土景观植物具有适应性强、地域代表性强、敏感度低、再生能力强且具有人文底蕴的优点[7]。因此,在乡村景观营造中,利用乡土植物进行景观设计不但可以达到良好的绿化和美化效果,也能更好地对本地生物多样性进行保护,同时更好地展现地方人文历史与文化特征。近几十年,国内外对于生物多样性的研究集中在自然区域,乡村生物多样性研究没有得到足够关注[8]。乡村作为人文自然共生地带,是生物多样性保护的基本场所[9]。生物多样性包含生态系统多样性、物种多样性和遗传多样性3 个层次[10]。在村镇区域,受人为干扰影响生态系统多样性和物种多样性的变化较为直观,而遗传多样性代表着一个群体内不同个体遗传变异的总和,在自然环境和人文影响下较难发现种群的变异差异。但种群遗传多样性的减少,会伴随着种群间分化增加、基因流减少而发生,易造成物种地域性濒危,最终对当地的生态环境产生严重影响[11]。乡土景观植物是乡村振兴和美丽乡村建设的重要载体,研究其群体遗传多样性水平,揭示物种在当地的基因资源分布,提出相对应的基因资源保护手段是极为必要的。乡土景观植物的遗传多样性研究方法从传统的形态学、细胞、生理生化水平逐渐深入到当前的DNA 分子标记水平[12]。形态标记测定操作较为方便、容易观察,通过对数量性状和质量性状的观测,可快速区别不同种、属[13];而DNA 分子标记测定较为精准,利用分子标记位点[随机扩增多态性、简单重复序列、扩增片段长度多态性、简单重复序列区间(ISSR)]进行引物设计开发,基于PCR 技术扩增基因组目标片段,从基因水平揭示群体之间、个体之间的变化差异[14]。在一系列分子标记中,ISSR 标记具有不需要基因组序列信息,能有效在种内水平区分形态高度相似的个体、技术相对简单、成本相对较低以及产生高度多态性等优势,适合对同一地区多个物种的遗传多样性进行研究[15]。
乡土景观植物是反映本土人文和自然文化、打造美丽乡村的关键物种,本研究以华东地区典型生态保护型乡村和旅游休闲型乡村作为研究区域,在对乡村植物群体特征调查的基础上,结合当地自然景观和人文景观特点,筛选出具有乡土景观价值的8 种特色物种作为乡土景观关键种。利用表型标记和ISSR 分子标记计算并分析8 种乡土景观物种的遗传多样性。基于其他景观植物群体遗传多样性数据的统计,划分并评价乡土景观植物的遗传多样性水平;分析不同区域乡土景观植物群体的遗传多态性位点百分率、等位基因数和有效等位基因数,结合乡土景观植物自身特性,推测其遗传多样性变化的影响因素。研究结果将为乡村景观营造中乡土景观植物的选择、品种(品系)配置和保护提供理论依据。
1 研究区概况
华东地区位于我国东部沿海地带,该地区经济发达,自然条件优越,水资源和森林资源丰富,较早开展环境治理与保护且地域基础设施完善齐备、交通便捷,因此具有营造以生态保护和休闲旅游为模式的乡村的优势。本研究分别选择华东地区典型生态保护型乡村(常口村)和旅游休闲型乡村(牌坊社区)作为研究范例。
常口村位于福建省三明市将乐县(117.56°E,26.77°N),是典型的生态保护型乡村。常口村镶嵌在“美丽中国·深呼吸第一城”的将乐县东部,位于闽江支流金溪河畔,与省道204 线毗邻,距县城约15 km。村域面积约13.83 km2,其中耕地面积约80 hm2,林地面积约1266.67 hm2。拥有“山、水、林、田、湖、村”立地条件的常口村被评为“省级园林式村庄”和“省级生态村”等荣誉称号。将乐县是我国南方45 个重点林业县之一,地处武夷山支脉,位于福建省西部的低山丘陵地区;气候属中亚热带季风性气候,气候温润,年均温为18.7 ℃,雨季时间长,雨量充沛;以红土为主的土壤类型,适合杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)和银杏(Ginkgo biloba)等喜微酸性土壤的重要用材的乡土树种生长。森林覆盖率高达84.5%,混合分布有针阔叶林、针叶纯林、竹林等,丰富的森林资源更易于开展乡村景观植物遗传多样性的相关研究。
牌坊社区位于江苏省南京市江宁区(118.69°E,31.80°N),是典型的旅游休闲型乡村。牌坊社区位于江宁街道东南部,占地面积约8.2 km2,耕地面积约140 hm2,山林面积约333.33 hm2。从20 世纪60 年代开始,牌坊社区改良、培植优质茶园、竹林和杉木林。2013 年牌坊社区开始着力打造茶文化特色村,以成为金陵特色茶文化休闲旅游“第一村”为目标。江宁区属于亚热带季风气候,气候温和,年平均气温15.7 ℃,雨水充沛,适合多种乡土景观树种生长。江宁区素有“六山一水三平原”之称,有乡村旅游示范村24 个,是全国美丽乡村建设的典型示范区。乡村植物景观营造一直是美丽乡村建设中的重要课题,江宁区的旅游开发型美丽乡村景观建设总体已初见成效,逐渐由单纯绿化向彩化、美化、文化过渡。在乡村建设与人文情怀的双重驱动下打造的牌坊社区具有中国传统江南乡村的景观特点,该乡村关键种基因多样性的研究对于指导华东地区江南村镇的景观营造具有重要意义。
2 研究方法
2.1 乡村生境条件调查
于2021 年7-8 月,对乡村区域的地形、地貌、聚落类型(庭院、宅旁宅间、公共休闲、滨水、耕地、园地、林地)、人为干扰因素(焚烧、砍伐、采矿、耕种)进行调查。聚类区域具有1 项人为干扰因素认定为轻微干扰、2 项认定为一般干扰、3 项及以上认定为严重干扰。将村的地域管辖范围设定为整体村域。将干扰程度为一般及以上程度的聚落区域划定为人工区域,其他聚落区域划定为自然区域。
2.2 乡村景观植物种质资源调查
2.2.1 乡村景观植物确定
于2021 年7-8 月,对乡村植物群体特征进行调查,调查内容包括植物群体地理坐标、海拔高度、种群数量、乔木层主要优势种、灌木层主要优势种、草本层主要优势种等,结合当地村镇自然景观和人文景观特点,筛选出具有乡土景观价值(观赏、绿化、经济、文化和生态价值等)的特色物种作为乡土景观关键种。
2.2.2 种质资源样品采集
按照7 种聚落类型(庭院、宅旁宅间、公共休闲、滨水、耕地、园地、林地)对整体村域进行划分,并依据聚落点的人工干扰程度划分出人工区域和自然区域,分别对2 个区域的乡村景观植物进行随机抽样。
乔木样品: 样品间至少相隔200 m,人工和自然区域采集20 株以上,若区域植株不足20 株则全部采集;灌木样品: 样品间至少相隔15 m,人工和自然区域采集25 株以上,若区域植株不足25 株则全部采集;草本样品: 样品间至少相隔5 m,人工和自然区域采集尽可能多的样品。
2.3 基于表型性状的遗传多样性分析
2.3.1 表型性状测量
种质资源形态特征调查: 对植株的性别、整株特征、叶形态特征和花形态特征等主要形态性状进行调查统计,调查内容包括: 性别、株龄、株高、胸径、冠幅、树姿、树形、叶色、叶形、花色和花形。
1)叶片形态性状测量: 以关键种树冠外围正常长枝基部向上第3~5 个叶为标准叶。野生种每株采集30 片叶观测;栽培种每份种质采集3 个单株,每个单株分别观测30 片叶。利用数显游标卡尺测量叶柄长度、叶柄粗度、叶长、叶宽、叶厚度,使用Photoshop 2018 软件测量叶基角,使用精密天平测量单叶鲜重、单叶干重,分别统计形态数据[16]。对不同区域、不同植株的叶片形态数据计算平均值和标准差。
2)花形态性状测量: 以完全盛开且未出现花瓣凋零的花为观测对象,乔木植物每株采集30 个以上花朵,草本植物以5 m 为直径采集样地内所有花朵。利用数显游标卡尺测量花序总梗长度、花梗长、裂瓣深度、花冠直径,使用精密天平测量百朵鲜花重[17]。对不同区域、不同植株的5 个花形态数据计算平均值和标准差。
2.3.2 表型性状聚类分析
统计乡土景观植物种群所有形态性状数据,利用R 语言非加权平均法聚类分析[18-19],将形态相似数值归为一类,以最小相似系数为阈值划分种质资源类型。
2.3.3 表型性状变异系数计算
变异系数(CV)又称“离散系数”“标准差率”,是恒量数据间离散程度的一个值,用标准差与平均差的比值表示[20]:
式中: AVG 为关键种种群某形态数据平均值,SD 为关键种种群某形态数据标准差。
种质资源形态测量分别统计了叶、花的形态数据,利用公式(1)对不同植物形态数据的平均值和标准差进行运算,获得各形态变异系数。一般变异系数值越大,种群形态变异程度越高,反之越小。计算不同区域不同植物的表型变异系数,展现同一物种在不同区域(整体村域、人工区域和自然区域)内每一种乡土景观植物的表型变异程度。
2.3.4 表型Shannon 多样性指数计算
统计关键种单株各形态数据和种群各形态数据,计算单形态在种群形态中所占比例并进行log 函数降维,计算Shannon 指数(I)[20-21]:
式中:ni为单株性状数据,N为种群性状数据。
对关键种各单形态Shannon 指数求平均值,得到关键种种群的Shannon 指数,该指数用来估计群体多样性水平的高低,一般其值越大,表示群体遗传多样性越丰富。本研究以表型Shannon 指数展现同一物种在不同区域(整体村域、自然区域和人工区域)内每一种乡土景观植物的表型遗传多样性水平。
2.4 基于ISSR 分子标记的遗传多样性分析
2.4.1 植物样品保存
采集新鲜、幼嫩、无病虫害的植物叶片样品,取适量样品放在玻璃皿中,用清水清洗去除表面灰尘,擦干后剪成3~4 mm2的碎片备用。DNA 保存液滴加到保存管(1.5 mL 级别) 1/5 处,将备用样品加入,用封口膜顺时针进行封口,4 ℃低温保存[22-23]。
2.4.2 植物基因组DNA 提取
参照申林静[24]的方法进行样品DNA 的提取和保存。
2.4.3 ISSR-PCR 反应和产物检测
1) ISSR-PCR 引物的筛选: 查找哥伦比亚大学ISSR 通用引物库(UBC primer set no.9,University of British Columbia,Canada),随机筛选备选引物进行扩增试验。
2) ISSR 反应体系的构建: ISSR-PCR 的10 μL 优化反应体系含2× Master Mix (包含Taq 热启动DNA聚合酶) 5 μL、2 μmol∙L-1ISSR 引物1 μL、<1 μg DNA 模板,加无菌去离子水至10 μL。反应程序为96 ℃预变性2 min;96 ℃变性10 s;50 ℃退火30 s;72 ℃延伸1 min,30~35 个循环;72 ℃延伸1 min;4 ℃保存[24]。
3) ISSR-PCR 反应物检测: PCR 产物用2.5%琼脂糖凝胶电泳进行检测,220 V 电压下电泳25~30 min,于紫外凝胶成像系统进行观察、拍照记录。
2.4.4 植物遗传多样性分析
1) ISSR 指纹图谱的构建: 通过ISSR-PCR 扩增结果,记录每一个样本、每一条引物的扩增条带。扩增出的每个条带视作一个位点,根据凝胶同一位置DNA 条带的存在与否估算扩增条带的分子量。有条带记作“1”,没有条带记作“0”,统计表格(0,1)矩阵,构建关键种指纹图谱[25-26]。
2)期望杂合度计算分析: 采用Popgene 32 软件对各关键种进行遗传多样性分析,利用构建的矩阵计算Nei’s 基因多样性指数[27-28]。期望杂合度(he)是1973 年著名遗传学家Nei 在观测杂合度的基础上提出的用来度量群体遗传多样性水平的参数[27-28],其假定一个随机交配种群中的位点个数为1,按照公式(3)进行计算:
式中:r为某个位点的等位基因数,qi为第i个等位基因频率。
当被检测种群中有n个位点时,采用公式(4)进行计算,得到Nei’s 基因多样性指数(He)。
采用He评估种群基因多样性水平,其取值范围为0~1,He越高,关键种遗传多样性水平越高。
3)多态性位点百分率: 多态性位点(np)是指具有两个以上等位基因的位点,多态性位点百分率(PPL)指群体中多态性位点数占检测总位点数(n)的比例。一般多态性位点百分率越大,群体遗传多样性水平越高。PPL 用公式(5)计算:
4)等位基因数: 等位基因数(Na)指所有检测位点上等位基因数量的平均值,当遗传多样性越高时,其值越大,通常用公式(6)计算:
式中:ai为第i个位点的等位基因数。
5)有效等位基因数: 有效等位基因数(Ne)指与现实群体中观测到的纯合体频率相等、具有相同等位基因频率的理想群体(具有相等的等位基因频率)中的基因数目,通常用公式(7)计算:
式中:np表示多态性位点数,Pij表示第i个位点上第j个等位基因数的频率。
当有效等位基因数越接近等位基因数时,等位基因位点在群体中的分布越均匀。因此,本文利用Ne/Na值来反映等位基因的分布均匀度,Ne/Na值范围为0~1,此值越接近1,等位基因的分布均匀度越高。
2.4.5 统计分析
采用IBM SPSS Statistics 23 软件对人工区域和自然区域的乡土景观植物群体He和PPL 进行单因素方差分析和t检验。
3 结果与分析
3.1 乡土景观植物的筛选与种质资源库构建
通过对目标村落环境、植被的调查,结合当地村镇自然景观和人文景观的特点,在常口村筛选出3 种景观植物: 枫香(Liquidambar formosana)、福建青冈(Quercus chungii)、青冈(Quercus glauca);在牌坊社区筛选出5 种景观植物: 榉树(Zelkova serrata)、香椿(Toona sinensis)、马兰(Aster indicus)、野菊(Chrysanthemum indicum)和蓬蘽(Rubus hirsutus)。收集了枫香131 份(67 份自然区域、64 份人工区域)、福建青冈89 份(84 份自然区域、5 份人工区域)、青冈53 份(41 份自然区域、12 份人工区域)、榉树37份(6 份自然区域、31 份人工区域)、香椿34 份(3份自然区域、31 份人工区域)、马兰28 份(5 份自然区域、23 份人工区域)、野菊37 份(8 份自然区域、29份人工区域)、蓬蘽55 份(15 份自然区域、40 份人工区域)种质资源,具体见附表1-附表8 (附表见官网www.ecoagri.ac.cn/cn/article/doi/10.12357/cjea.20230326此文的资源附件,各附图同)。
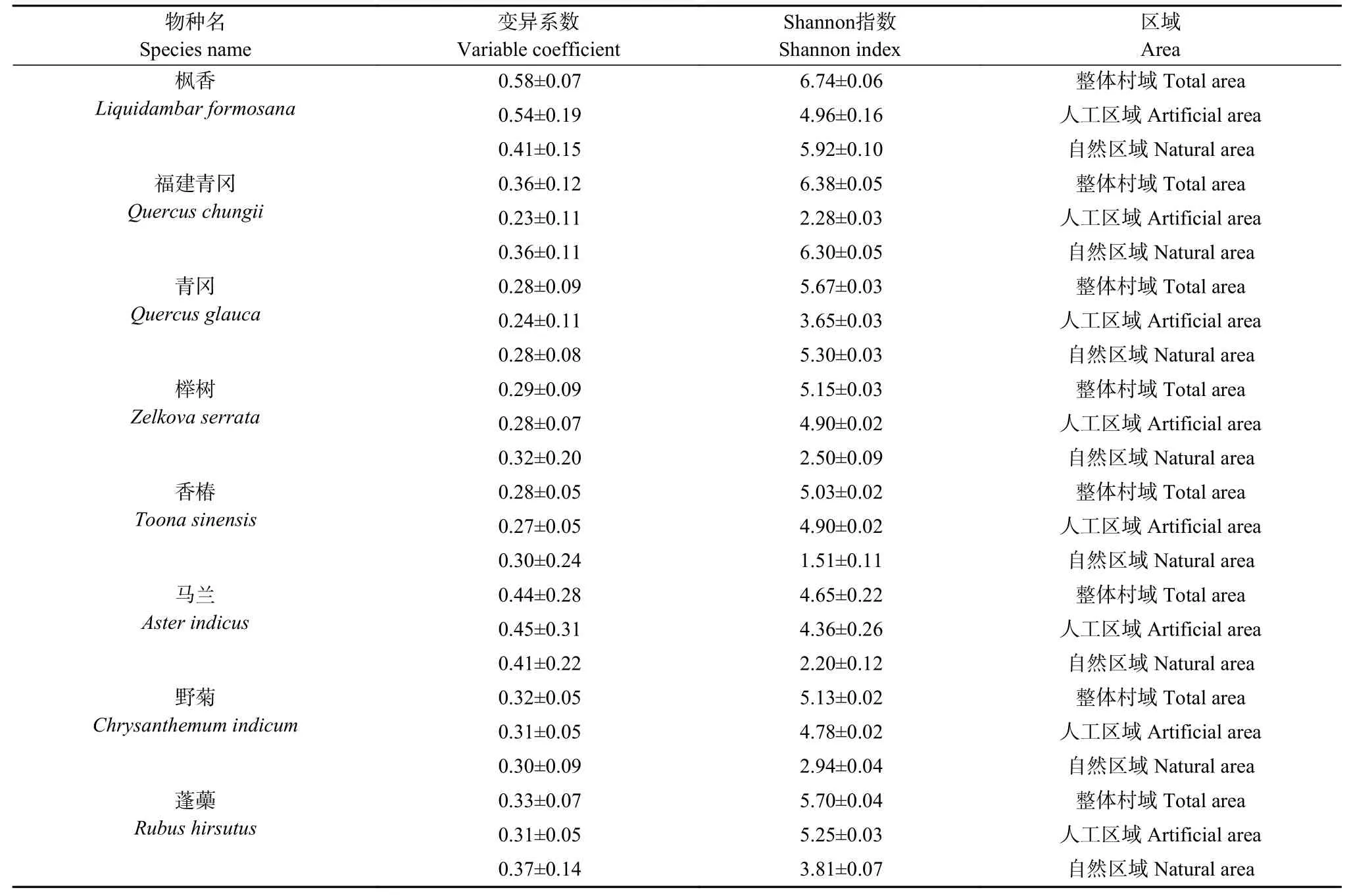
表1 乡土景观植物在不同乡村区域的表型变异系数和多样性指数Table 1 Phenotypic variation coefficients and Shannon indexes of rural landscape plants in different rural areas
3.2 乡土景观植物的表型性状聚类分析
以0.01 为阈值对常口村131 份枫香种质资源的表型性状进行聚类分析(附图1),划分为5 类品系资源(图1A);以0.01 为阈值对常口村89 份福建青冈种质资源的表型性状进行聚类分析(附图2),划分为6类品系资源(图1B);以0.012 为阈值对常口村53 份青冈种质资源的表型性状进行聚类分析(附图3),划分为4 类品系资源(图1C);以0.0028 为阈值对牌坊社区37 份榉树种质资源的表型性状进行聚类分析(附图4),划分为16 类品系资源(图1D);以0.001 为阈值对牌坊社区34 份香椿种质资源的表型性状进行聚类分析(附图5),划分为7 类品系资源(图1E);以0.0078 为阈值对牌坊社区28 份马兰种质资源的表型性状进行聚类分析(附图6),划分为10 类品系资源(图1F);以0.007 为阈值对牌坊社区37 份野菊种质资源的表型性状进行聚类分析(附图7),划分为5 类品系资源(图1G);以0.0023 为阈值对牌坊社区55 份蓬蘽种质资源的表型性状进行聚类分析(附图8),划分为6 类品系资源(图1H)。
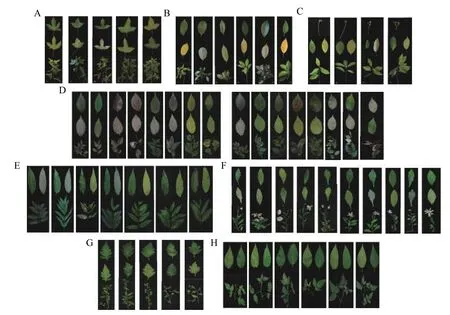
图1 乡土景观植物的品系资源Fig.1 Strain resources for rural landscape plants
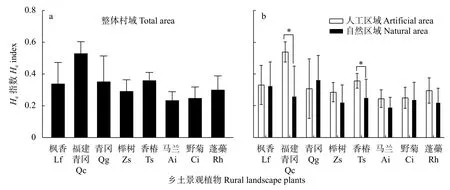
图2 不同乡村区域的乡土景观植物Nei’s 基因多样性(He)指数Fig.2 Nei’s genetic diversity (He) indexes of rural landscape plants in different rural areas
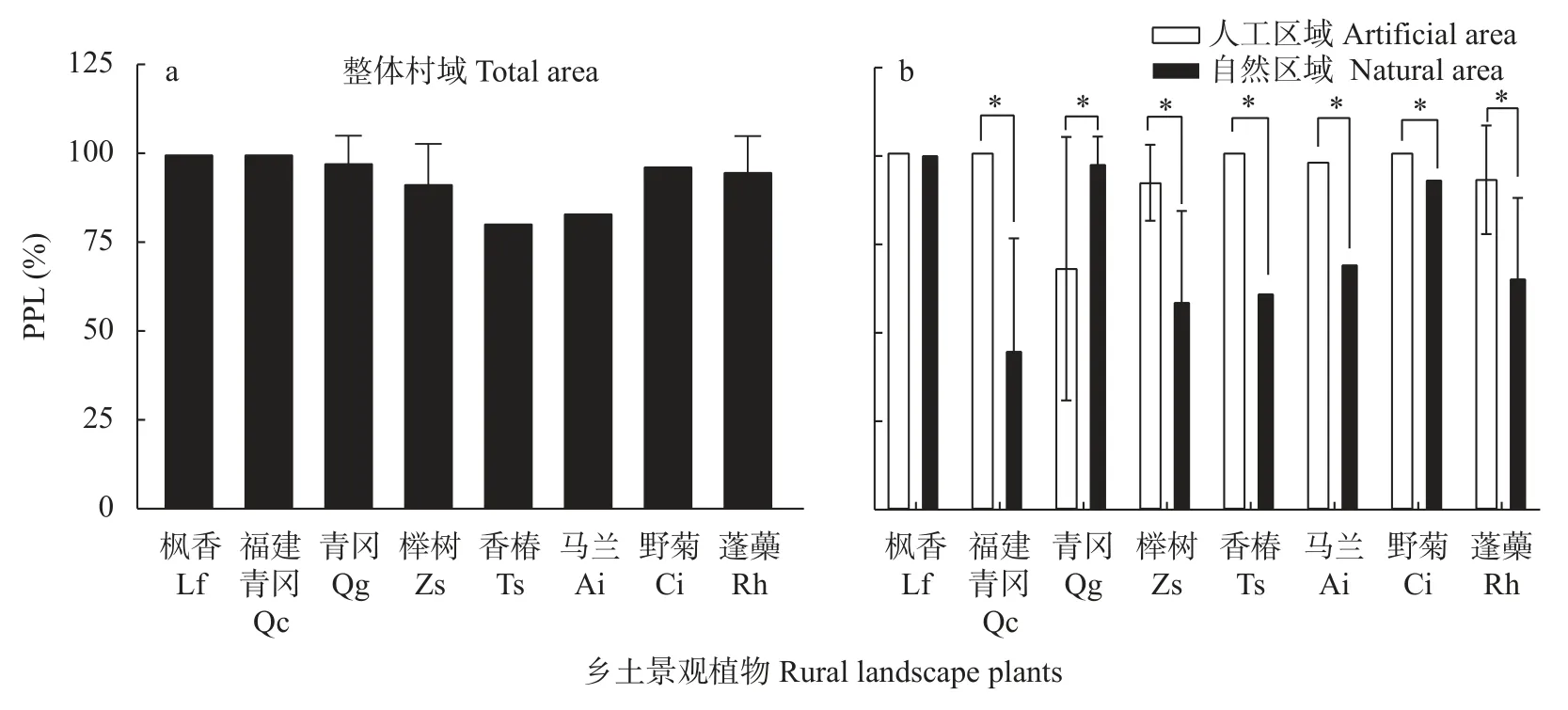
图3 不同乡村区域的乡土景观植物多态性位点百分率(PPL)Fig.3 Percentages of polymorphic loci (PPL) of rural landscape plants in different rural areas
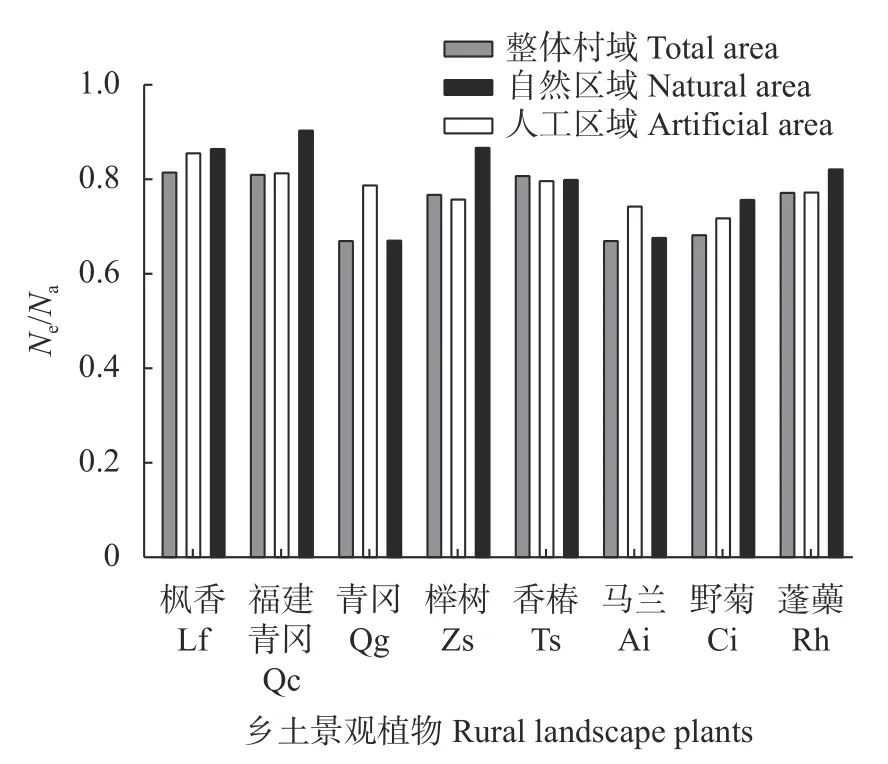
图4 不同乡村区域的乡土景观植物等位基因均匀分布程度[有效等位基因数量(Ne)与等位基因数(Na)差异水平(Ne/Na)]Fig.4 Degree of uniform distribution of allelles [ratio of number of effective alleles (Ne) to number of alleles (Na),Ne/Na] of rural landscape plants in different rural areas
3.3 乡土景观植物表型遗传多样性
基于变异系数和Shannon 指数,对乡村整体区域、人工区域和自然区域的景观植物表型遗传多样性进行分析。
3.3.1 表型变异系数
如表1 所示,8 个乡土景观植物的表型变异系数范围为0.23~0.58,其中枫香和马兰的表型变异系数值较高,枫香在整体村域、人工区域和自然区域变异系数分别为0.58、0.54 和0.41,马兰在整体村域、人工区域和自然区域变异系数分别为0.44、0.45 和0.41,其他植物的变异系数在0.23~0.37 区间。8 个植物中,枫香、马兰和野菊在人工区域的表型变异系数高于自然区域,而其他植物反之。较高的表型变异系数预示着植物种群个体表型性状差异较大,枫香和马兰在乡村分布区域或有着较为丰富的表型类型。
3.3.2 表型Shannon 指数分析
如表1 所示,8 个乡土景观植物的整体村域表型Shannon 指数范围为4.65~6.74,枫香和福建青冈Shannon 指数较高,分别为6.74 和6.38。人工区域Shannon 指数范围为2.28~5.25,自然区域范围为1.51~6.30。Shannon 指数反映了植物表型多样性的丰富度,8 个植物在整体村域的表型多样性丰富度都较高,但是部分植物在人工或自然影响下表型多样性较低。
3.4 乡土景观植物分子遗传多样性分析
本研究从哥伦比亚大学ISSR 通用引物库中为8个植物物种分别筛选出10 个可高效扩增的ISSR 通用引物,引物序列详明如附表9 所示。物种的群体内Nei’s 基因多样性指数(He)、多态性位点百分率(PPL)和有效等位基因数(Ne) 3 项指数来体现。
3.4.1 乡土景观植物Nei’s 基因多样性指数分析
通过对扩增有效条带的测试与统计,计算不同区域Nei’s 基因多样性指数。如图2 所示,8 种乡土景观植物在整体村域的Nei’s 基因多样性指数为0.240~0.536,其中福建青冈最高,枫香、青冈、香椿、蓬蘽指数范围在0.3~0.4 区间,榉树、马兰、野菊指数范围在0.2~0.3 区间。
对人工区域和自然区域8 种乡土景观植物的Nei’s 基因多样性指数进行计算。如图2 所示,8 种植物的Nei’s 基因多样性指数在人工区域为0.244~0.540,福建青冈最高,枫香、青冈和香椿在0.3~0.4区间,榉树、马兰、野菊和蓬蘽在0.2~0.3 区间;在自然区域Nei’s 基因多样性指数为0.193~0.367,枫香和青冈在0.3~0.4 区间,福建青冈、榉树、香椿、野菊和蓬蘽在0.2~0.3 区间,马兰指数值最低。除青冈外,其他7 种植物在人工区域的Nei’s 基因多样性指数均高于自然区域,其中福建青冈及香椿在人工区域显著高于自然区域。
3.4.2 乡土景观植物多态性位点百分率分析
通过对扩增有效条带的测试与统计,计算8 个植物不同群体分子标记的多态性位点百分率。位点的多态性反映了分子标记在群体中的变异差异,多态性百分率越接近1 证明该群体的遗传多态性越高。如图3 所示,8 种植物在乡村整体区域的多态性位点百分率范围为80.56%~100.00%。其中枫香和福建青冈的遗传多态性极高,其分子标记多态性位点百分率为100.00%,其他植物的多态性位点百分率均超过了80%,证明8 种景观植物在乡村整体区域的遗传多态性水平较高。
从图3 可知,8 种植物在人工区域的多态性位点百分率范围为67.50%~100.00%,其中枫香、福建青冈、香椿和野菊的多态性位点百分率为100.00%,遗传多态性高。青冈多态性位点百分率仅为67.50%,与其他植物的遗传多态性水平差距较大。8 种植物在自然区域的多态性位点百分率范围为45.00%~100.00%,其中枫香、青冈和野菊的遗传多态性较为丰富,而其余植物的多态性位点百分率均未超过70%,其中福建青冈最低。
枫香在自然区域和人工区域的多态性位点百分率均为100.00%,不同区域遗传多态性水平均较高;另外7 种植物自然区域和人工区域的多态性位点百分率差异皆达到显著水平,其中青冈在自然区域的遗传多态性水平较高,在人工区域较低,另外6 种植物都是在人工区域有较高的遗传多态性水平,但在自然区域较低。8 个植物在自然区域和人工区域的多态性位点百分率变化趋势与Nei’s 基因多样性指数变化相似。不同区域中,一种乡村景观植物群体的遗传多样性指数越高,该群体的多态性位点百分率值越趋近1,群体的遗传多态性水平越高。
3.4.3 乡土景观植物有效等位基因数分析
有效等位基因数是反映植物群体遗传变异大小的指标,其数值越接近所检测到的等位基因绝对数,表明等位基因在群体中分布越均匀。如表2 所示,8个植物在整体区域的等位基因数范围为1.92~2.00、有效等位基因数范围为1.33~1.64,人工区域等位基因数范围为1.68~2.00、有效等位基因数范围为1.33~1.64,自然区域等位基因数范围为1.45~1.98、有效等位基因数范围为1.30~1.57。
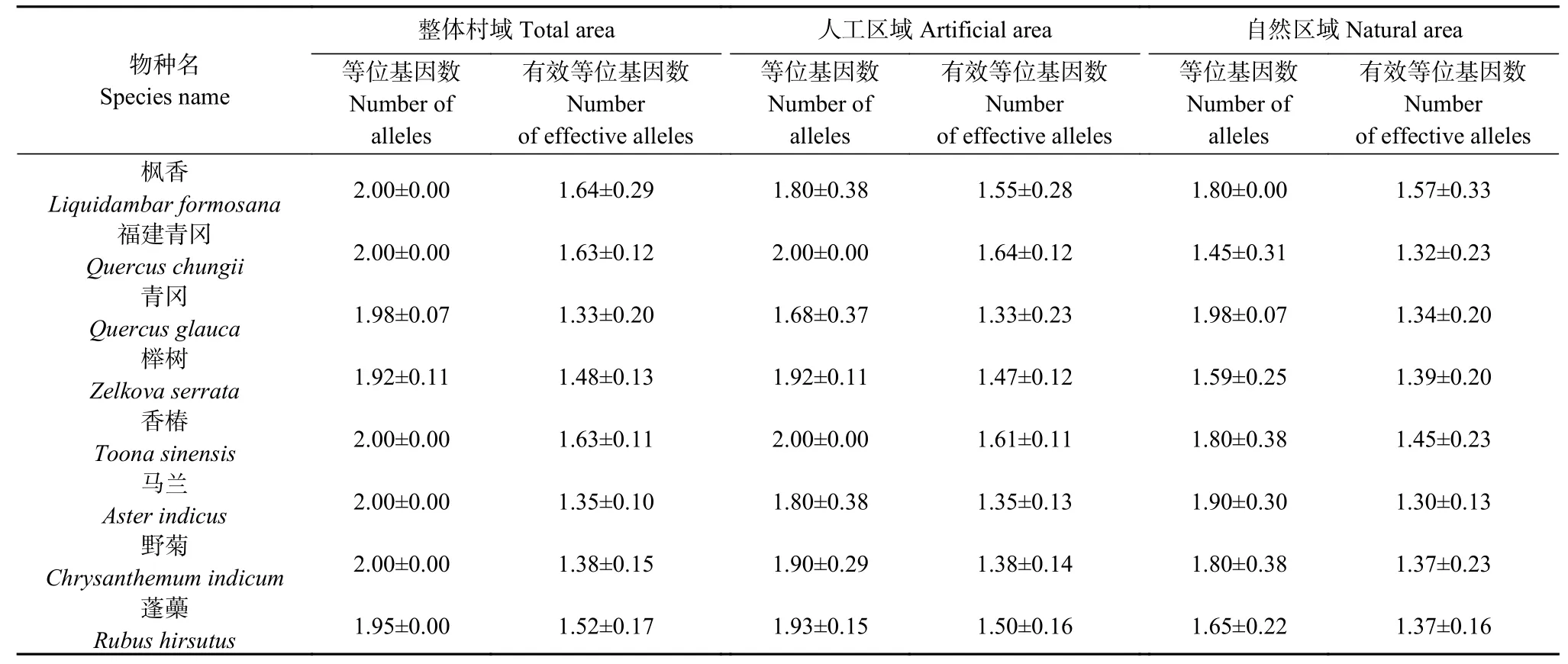
表2 不同乡村区域的乡土景观植物等位基因数和有效等位基因数Table 2 Number of alleles and effective number of alleles of rural landscape plants in different rural areas
将不同区域各植物的有效等位基因数(Ne)除以等位基因数(Na),反映有效等位基因数与等位基因数差异水平,以此体现景观植物的等位基因均匀分布程度。如图4 所示,枫香、福建青冈、香椿在整体村域、人工区域和自然区域的Ne/Na值均大于0.8,表明3 个物种的有效等位基因数与等位基因数较接近,预示相关等位基因在各群体分布均匀度较高。榉树和蓬蘽在自然区域的Ne/Na值大于0.8,表明二者在该区域群体等位基因分布均匀度较高;在整体村域和人工区域的Ne/Na值范围为0.7~0.8,等位基因分布均匀度中等。野菊在人工和自然区域的Ne/Na值为0.7~0.8,等位基因分布均匀度一般;在总体区域Ne/Na值小于0.7,等位基因分布均匀程度较低。青冈和马兰在人工区域的Ne/Na值为0.7~0.8,等位基因分布均匀度中等;在总体和自然区域的Ne/Na值小于0.7,等位基因分布均匀度较低。当群体基因Ne/Na值较低时会影响群体遗传多样性水平,需采取相关措施进行维护和提高。
4 讨论与结论
遗传多样性能够反映植物在不同环境条件下为了生存而进行的变异,极大地反映目标物种未来的生存发展[29]。本研究以华东地区典型乡村福建省常口村和南京市牌坊社区为对象,利用表型性状和分子标记遗传多样性分析技术,分别以表型Shannon 指数和Nei’s 基因多样性指数来评价乡土景观植物的表型多样性和基因多样性,旨在揭示村镇乡土景观植物遗传多样性的水平及变化规律,同时结合乡村实际情况,讨论影响遗传多样性变化的因素,为村镇乡土景观植物遗传多样性维护技术的提出提供科学依据。
乡村乡土景观植物需要与当地建筑、地形、河流等组成具有地域独特性的景观风景,以达到美化人居环境的目的[30]。景观植物常以叶、花、果等表型性状来体现其观赏性,研究乡村景观植物的表型遗传多样性能揭示群体形态特征差异性,并直观、快速地筛选出观赏价值高的优异种质资源[31]。本研究中,8 个乡土景观植物的整体村域表型Shannon 指数大于1.90,高于其他景观植物,如月季(Rosa hybrida,1.70)[32]、秋海棠(Begonia grandis,0.93)[33]、杉木(1.74)[34]、云南松(Pinus yunnanensis,1.40)[35]、观赏桃(Prunus persica,1.73)[36]、南方红豆杉[Taxus wallichianavar.mairei,1.08)[37]、观赏石榴(Punica granatum,0.60~1.16)[38],表明本研究选择的乡土景观植物的表型遗传多样性高,表型特征丰富,种质资源类型多样。虽然在不同区域受人为因素影响,不同景观植物的表型Shannon 指数有所区别,但各区域Shannon 指数基本高于平均数1.90 (自然区域的香椿群体除外),表型多样性较为丰富。由于表型是基因与环境互作表达的结果,性状的表达易受环境影响。因此,需要进一步利用分子标记遗传多样性分析,精准揭示乡村景观植物的基因多样性。
基于分子标记的基因多样性研究在于揭示群体内的遗传变异水平,而各物种基因多样性不易受到环境和发育引起的短期变化的影响[39]。为易于划分和评价乡村景观植物ISSR 分子遗传多样性,我们统计分析了女贞(Ligustrum lucidum,0.144)[40]、凤仙花(Impatiens balsamina,0.252)[41]、白木香奇楠(Aquilaria sinensis,0~0.141)[42]、枫香(0.178~0.372)[43]、黄杨叶栒子(Cotoneaster buxifolius,0.156~0.414)[44]、地黄(Rehmannia glutinosa,0.175~0.256)[45]、白花兜兰(Paphiopedilum emersonii,0.187~0.352)[46]、赤芍(Paeoniae lactiflora,0.090~0.190)[47]等景观植物ISSR 的Nei’s 基因多样性指数,发现上述景观植物的Nei’s 基因多样性指数范围为0~0.414,平均值为0.208。在乡村整体村域,福建青冈的Nei’s 基因多样性指数大于0.414,表明其在整个区域的遗传多样性水平极高;其他7 个植物的Nei’s 基因多样性指数大于0.208,表明本研究所选择的8 种乡土景观植物在整体村域的遗传多样性水平较高,乡村植物遗传多样性维护较好。在人工区域,福建青冈的Nei’s 基因多样性指数大于0.414,其他7 个植物的Nei’s 基因多样性指数范围在0.208~0.414 区间,表明人工区域的乡村景观植物的遗传多样性较高。而自然区域中,除马兰外其他7 种植物的Nei’s 基因多样性指数均高于0.208,遗传多样性水平高。物种遗传多样性越高或遗传变异越丰富,其环境适应能力越强,研究结果表明,8 种乡土景观植物具有较强的环境适应性与抗逆性,适合生长在当前区域内。
枫香在各区域(整体村域、人工区域、自然区域)表型和分子遗传多样性均较高,是群体遗传多样性较为稳定的乡土景观植物。作为我国优良观赏树种,枫香主要分布于黄河流域及其以南地区,主产于福建、浙江、江苏等省份[48]。枫香适应能力强、叶色叶形美丽,是维护改善生态景观的先锋树种[49]。福建省常口村作为生态保护型示范村,封山育林长达30 年,生态修复维护地带大量引种了枫香优良品系,同时作为乡村风水林,村民禁止随意砍伐枫香树,保护了本土枫香种源(特别是枫香古树)。引种枫香资源与本地资源一同维护了村域内枫香群体遗传多样性的水平,改善了乡村生态景观。因此,研究发现枫香的Nei’s 基因多样性指数、多态性位点百分率和Ne/Na值高,表明常口村的枫香基因资源丰富,群体内个体基因交流频繁,已与本土其他景观植物构成良好的生态景观。
福建青冈在整体村域和人工区域分子遗传多样性极高,Nei’s 基因多样性指数分别达到0.536 和0.540,高于统计的景观植物Nei’s 基因多样性指数范围0~0.414,而自然区域Nei’s基因多样性指数较低(0.263)。福建青冈又名黄楮、黄丝稠木,其木材具有坚重、耐磨、耐水湿的特性,是重要的工业原材和烧制上等白炭的原材料,在人工区域曾有过大量砍伐的记录。从20 世纪80 年代开始,福建省多地建立楮林保护区,种群得到了修复和保护[50-53]。在许多人为活动被限制的人工区域(滨水河谷、乡村道路、耕地园地),较少受到人为干扰,群落中出现大量的福建青冈小树和幼苗,种群分子遗传多样性指数和遗传多样性水平得到提高[54-55]。自然区域的福建青冈群体结构保持长期的稳定,具有较高的遗传变异水平,但是等位基因多态性位点百分率低于人工区域。说明自然区域福建青冈群体的遗传多态性程度低于人工区域,可以通过从其他区域引种补种优异种质资源,采用人工授粉方式促进群体内基因交流,提高自然区域福建青冈的遗传多样性水平。与福建青冈类似,榉树、香椿、马兰、野菊、蓬蘽在总体村域和人工区域的Nei’s 基因多样性指数高,而在自然区域有所下降,更证明人工维护措施在提高村镇乡土景观植物遗传多样性水平方面的重要作用。
通过表型性状与ISSR 分子标记所提供的DNA片段信息相结合,可以揭示乡村景观植物遗传的本质特征,对于制定科学的保护策略有重要作用。本研究利用表型和分子标记遗传多样性检测技术,对8 个乡土景观植物的遗传多样性进行了测定与评价。结果表明,8 个景观植物在乡村中的表型遗传多样性(Shannon 指数评价)和分子遗传多样性(Nei’s 基因多样性指数评价)水平都较高,各群体遗传变异丰富、结构稳定,适宜的遗传多样性维护措施在乡村生态景观建设中发挥着稳定而重要的作用。
