微生物介导的稻田水土界面温室气体排放及其农事减排措施研究进展
2023-12-20韩燕云吴永红俞元春
韩燕云,吴永红,李 丹,4*,俞元春
1. 南京林业大学生态与环境学院,南京林业大学南方现代林业协同创新中心,江苏 南京 210037
2. 中国科学院南京土壤研究所,江苏 南京 210018
3. 中国科学院大学南京分院,江苏 南京 210018
4. 南京工业大学城市建设学院,江苏 南京 210018
全球气候变暖是当前人类社会亟待解决的关键生态环境问题,温室气体过度排放是导致全球变暖的主要根源. 作为重要的温室气体,二氧化碳(CO2)、甲烷(CH4)、一氧化二氮(N2O)每年正分别以1000×10—9、7×10—9和0.7×10—9容积比的速度增加,其温室气体效应贡献率分别高达60%、15%和5%[1]. 农业占全球人为温室气体排放量的近12%. 水稻是农业利用的重要形式,全球种植面积约为1.55×108hm2. 据报道,全球农田温室气体排放量约有48%来自稻田[2]. 闫琪等[3]基于DayCent 模型预测,相较于2001—2020 年,2021—2040 年我国暖温带区域农田N2O、CH4和CO2排放量分别增加22.8%、13.6%和6.7%,2041—2060 年将分别增加24.9%、13.4%和8.0%. 农田温室气体减排具有长期必要性,需要持续关注.
作为最大的稻米生产国,中国稻田面积约为3.0×108hm2,占全球水稻种植面积的23%,位居第二;水稻总产量占全球的30%,位居第一[4]. 据统计,我国稻田CH4排放量为7.7 Tg,约占我国农业源排放量的23%[5]. CH4的产生主要是产甲烷菌在厌氧状态下分解利用土壤有机碳的结果,CH4产生后有一部分将会被土壤中的甲烷氧化菌利用,未被利用的CH4将被排放到大气中. N2O 可在好氧和厌氧条件下通过硝化作用和反硝化作用而产生. 稻田土壤动植物、微生物呼吸作用及生物化学作用是稻田土壤排放温室气体的主要途径.
稻田既是重要的温室气体排放源,也是温室气体“源汇”调节器. 稻田生态系统中独特的水分管理模式使水稻土形成了具有独特湿地特征的水土交相界面[6]. 稻田水土界面作为水面和土面重要的界面区,具有最为复杂的物质和能量交换过程,其特殊的边界效应使其成为生物地球化学循环的“热区”. 一方面,稻田水土界面所提供的水环境聚集了高度密集的微生物聚落,促进地上和地下动态过程相互作用,同时稻田上覆水在土壤中的垂直运动和水平运动会影响碳排放和传输[7];另一方面,稻田水土界面对外界环境变化十分敏感,加之人为活动的干扰,这使得稻田水土界面温室气体的排放更复杂. 徐志波[8]研究发现,常年淹水区域与常年落干区域的CO2、CH4排放量存在显著差异. 因此水土交相界面的存在增加了水田和旱田温室气体排放通量的差异性,同时增加了稻田温室气体通量排放的复杂性. 土壤微生物是稻田温室气体的源和汇,稻田的复杂环境会影响微生物群落结构构建和物种组成. 同时,微生物群落对环境变化的响应又会在一定程度上影响温室气体产生. 因此,系统性地讨论环境因素在稻田土壤-微生物-植物共作系统中对水土界面温室气体排放的影响至关重要.
土壤微生物是稻田生态系统中最重要和最活跃的成分之一,作为土壤碳氮转化的重要媒介,直接制约温室气体的排放. 甲烷菌和产甲烷菌综合作用,共同决定稻田CH4的排放. N2O 的产生是由氨氧化细菌和亚硝酸盐氧化细菌介导的硝化作用以及由反硝化菌介导的反硝化作用共同支配、耦合发生的结果.通过改变土壤微生物的群落结构、酶活等可实现调控温室气体的排放. 例如,土壤中添加解淀粉芽孢杆菌(Bacillus amyloliquefaciens)可显著减少氨氧化细菌,抑制硝化过程,同时增加了N2O 还原菌丰度,反硝化过程增强,进而减少了50%的N2O 排放通量[9].此外,通过激活固氮根瘤菌的N2O 还原酶,可以促进N2O 还原为N2,从而减少N2O 的排放[10]. 通过反复富集高浓度CH4、激活低亲和力甲烷氧化菌来增强CH4消耗,可实现CH4的减排[10]. 目前,微生物群落介导的温室气体排放过程对环境因素、人为活动因素的响应机制,以及微生物调控温室气体的排放机制等尚不明确,亟待研究.
农事管理措施对温室气体排放具有显著影响. 通过采用可持续的农业实践,如改进水分管理、肥料管理等,可在保证水稻稳产的基础上实现温室气体的减排. 例如,在水稻生长季节实施间歇灌溉,可以引发周期性的好氧条件,从而有效减少CH4的排放[11]. 生物炭还田可通过改变土壤碳氮底物、氧化还原条件及微生物组分进而显著减少CH4排放[12]. 除常规农事管理外,微生物调控也是实现稻田温室气体减排的重要措施之一.
本文系统梳理了稻田土壤微生物介导的温室气体(CH4、N2O 和CO2)产生与排放机理;多角度分析了稻田生态系统中微生物、水分、温度、土壤氧化还原电位以及植物自身特性对稻田水土界面温室气体排放的影响;重点研究了土壤微生物调控及水分管理、养分管理等农事管理模式下土壤温室气体的减排效果及作用机理;最后,结合目前稻田温室气体研究现状,展望未来亟待深入探索的研究方向,以期为稻田温室气体减排提供新视角,为实现农田温室气体减排、减缓全球变暖提供了理论支持.
1 水土界面温室气体排放机制
稻田的碳循环系统主要涉及光合作用的碳固定和碳释放过程. 水稻通过光合作用固定大气中的CO2,将碳储存于植物体并转化为有机物. 与此同时,水稻通过呼吸作用将部分CO2释放回大气中. 土壤碳排放过程通过呼吸作用实现,主要包括植物根系呼吸、土壤动物、微生物呼吸以及由含碳矿物质的化学氧化作用. 其中,土壤中含碳物质的化学氧化作用和土壤动物的呼吸作用通常十分微弱,往往忽略不计[13],而植株根系呼吸和土壤微生物呼吸的贡献较大,根系呼吸的贡献为30%~70%[14]. 在稻田生态系统中,CO2的固定与排放是一个动态循环过程,其影响因素众多,如稻田土壤有机质及其组分、土壤水分含量、微生物群落以及温度、光照等环境因素,均可通过直接或间接影响土壤微生物的呼吸作用和光合作用等来影响稻田CO2的排放.
稻田CH4的排放是由土壤CH4的产生、氧化及传输三个过程相互作用的结果. 产CH4过程是CH4排放体系极其关键的一步,是土壤有机物在淹水厌氧条件下,经产甲烷菌分解形成的最终产物[15]. 如图1所示,CH4是由有机物通过以下步骤进行分解产生:复杂聚合物在水解酶的作用下转变为单体化合物,并进一步转变成脂肪酸、CO2和H2;脂肪酸在产氢产乙酸菌作用下氧化生成乙酸、CO2和H2;此后,乙酸、H2和CO2分别被乙酸营养型产甲烷菌、氢营养型产甲烷古菌利用,产生CH4. 此外,同型产乙酸菌也可将H2、CO2转变为乙酸,并经乙酸营养型产甲烷古菌转变成CH4[16]. 与稻田CH4产生相似,CH4的氧化过程是CH4排放的重要组成部分. 甲烷氧化菌作为一种好氧细菌,在甲烷氧化过程中发挥着极其关键的作用. 甲烷氧化菌可通过甲烷单氧合酶(Methane monooxygenase)将CH4等还原物转化为CO2、H2O和其他中间代谢物,如CH3OH、甲酸(HCOOH)和甲醇(CH4O). 稻田环境中产生的90%以上的CH4可被甲烷氧化菌消耗[17],剩余的CH4通过气泡传输、扩散传输和植物通气组织传输这三种途径完成从稻田土壤或水层到大气的传输.
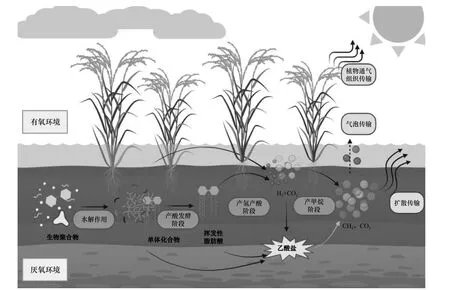
图1 稻田水土界面产甲烷示意Fig.1 Schematic diagram of CH4 production at the interface between soil and water in rice fields
水-土界面是微生物活动的关键场所,会直接影响微生物介导甲烷气体的排放过程. 稻田系统水土界面受干湿交替影响剧烈,历经好氧和厌氧环境的转换,影响水土界面微生物组成及其活动,最终影响CH4的排放过程.
氮功能微生物介导稻田水土界面氮循环过程. 氮功能微生物的生长和分布与水土界面氧化还原生境密切相关. 如图2 所示,稻田N2O 主要是在微生物作用下通过硝化和反硝化作用产生的. 在通气良好的条件下,土壤中的硝化细菌将氨氧化为硝酸和亚硝酸的过程为硝化作用,其主要分为氨氧化和亚硝酸盐氧化两个步骤. 氨氧化过程是指在氨氧化细菌(AOB)和氨氧化古菌(AOA)的作用下,氨或铵盐转化为亚硝酸盐(NO2—) 的过程. 在这个过程中,中间产物羟胺(NH2OH)发生不完全氧化产生N2O. 亚硝酸盐氧化过程是指亚硝酸盐在硝化细菌的作用下转化为硝酸盐的过程. 其中,氨氧化是硝化过程中的限速步骤,即整个硝化作用的速率受氨氧化过程的影响. 反硝化作用作为N2O 产生的重要过程,是在反硝化细菌或化学还原剂的作用下,硝酸盐在硝酸盐还原酶(Nar)、亚硝酸盐还原酶(Nir)、一氧化氮还原酶(Nor) 以及氧化亚氮还原酶(Nos) 的作用下,最终被还原成N2的过程[18]. 硝化-反硝化作用通常耦合发生,二者共同支配N2O 的产生.
2 微生物介导的水土界面温室气体排放影响因素
稻田温室气体的排放不仅受土壤中不同微生物活动的影响,还与水稻自身植物特性介导的气体输送有关;此外,稻田温室气体的排放还受土壤氧化还原电位、含水量等环境因素的影响,具体如表1 所示.
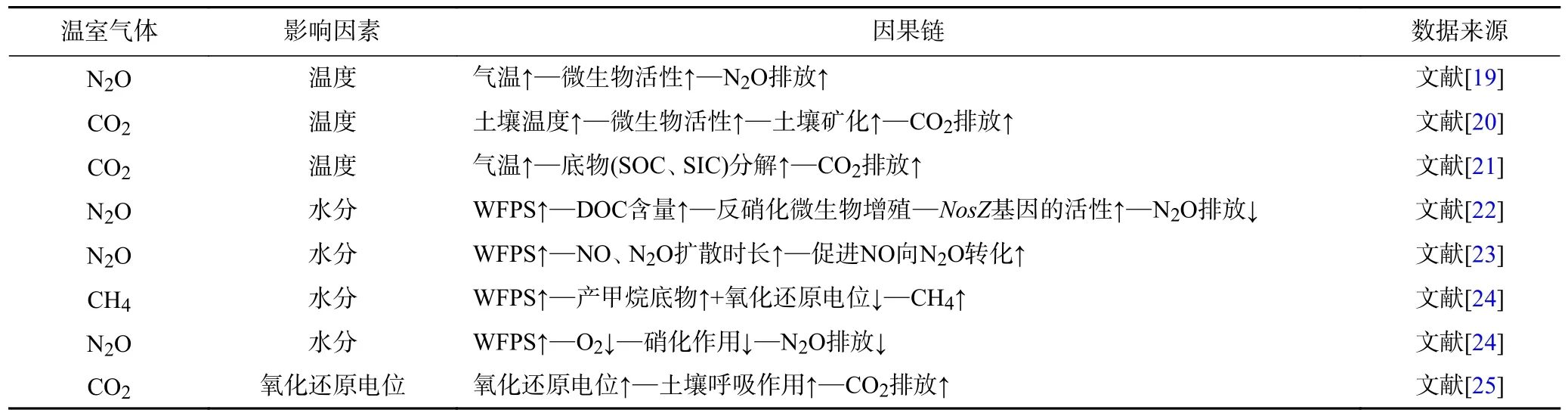
表1 稻田生态系统GHGs 的重要因果链Table 1 Important cause-effect chains for GHGs in paddy ecosystems
2.1 土壤微生物
土壤微生物作为稻田生态系统碳氮循环的关键驱动者,在陆地碳氮循环中发挥着重要作用. 一方面,自养微生物可以固定大气中的CO2并将其转移到土壤碳库中;另一方面,微生物的呼吸和分解作用可以将土壤中的有机碳转化为CO2并释放到大气中. 土壤微生物对土壤呼吸的贡献率约为20%[26]. 不同微生物群落组分的土壤呼吸速率各有差异. 屈冉等[27]发现,土壤呼吸速率与放线菌、土壤细菌数量呈显著正相关,与真菌数量则呈显著负相关. 另有研究[28]表明,土壤呼吸速率与微生物量碳含量呈正相关,与微生物量氮含量呈负相关. 产甲烷菌和甲烷氧化菌在CH4排放中起着关键作用. CH4的产生主要由氢营养型产甲烷菌和乙酸营养型产甲烷菌完成,其中2/3 的CH4产生于乙酸营养型甲烷菌,不到1/3 的CH4产生于氢营养型甲烷菌[29]. 除此之外,地杆菌(Bacillus subtilis)作为土壤最常见的电活性细菌之一,可与产甲烷菌形成的共生与竞争关系,参与中间代谢产物的转化,促进CH4产生[30]. CH4的氧化作用是由甲烷氧化菌完成. 不同微生物种类对N2O 排放的贡献存在差异[31].已有研究[32]表明,在酸性土壤中,相对于细菌,真菌在N2O 产生方面扮演更重要的角色. 真菌可通过反硝化作用生成N2O,因此减少酸性土壤中真菌数量可以降低N2O 的排放量. 此外,地杆菌作为氮循环的参与者,可以利用亚铁作为电子供体进行富集,利用厌氧环境中的硝酸盐完成硝酸盐异化还原成铵;同时地杆菌可以产生导电菌毛,用于碳源和远距离电子转移[33],从而诱导反硝化过程中硝酸盐的快速转化,减少N2O 的排放. 部分含有nosZII基因的微生物因缺乏亚硝酸盐还原酶基因而在反硝化过程中不产生N2O[34],因此这类微生物具有很大的N2O 还原潜力.
土壤介导微生物过程与功能菌群、功能基因密切相关,基因编辑、相关功能基因诱导调控均可不同程度地影响温室气体的排放. 但由于生物分子学技术的有限性,其相关机制尚不明确,限制了基于微生物技术调控温室气体减排的发展. 但尽管如此,微生物的调控技术在实验室的研究中已表现良好的温室气体减排潜势.
2.2 水分
土壤含水量通过影响土壤的通透性、底物浓度、微生物的组分和功能等影响土壤呼吸,进而影响土壤CO2的产生. 充足的水分可促进土壤中氧气、可溶性基质的扩散以及好氧微生物的活动,利于CO2的排放. 极端潮湿土壤环境的通透性差,导致氧气置换受阻,抑制稻田土壤动植物的呼吸而减少CO2排放;而极端干燥土壤环境中可溶性基质扩散有限,导致呼吸底物浓度降低,从而减少了土壤CO2的产生[35]. 土壤含水量作为影响CH4排放的关键因素,通过影响稻田土壤氧化还原电位、产CH4菌活性、CH4扩散速率等而影响稻田CH4的排放. 高含水环境下导致的低氧化还原电位是CH4产生的先决条件. 高含水量环境中所形成的厌氧区,促进产甲烷菌生长,同时又制约甲烷氧化菌的活性. 此外,土壤水分还可通过限制CH4在水土界面的扩散、增加有效水溶性碳基质等影响CH4排放[36]. 有研究发现,稻田CH4和N2O 的排放具消长(trade off)关系,这主要归因于水分变化,淹水的时间越长,CH4的排放量越大,N2O 的排放则会受限[37]. 水分通过影响土壤的硝化作用、反硝化作用以及土壤通气状况进而影响N2O 的排放与传输.Bateman 等[38]发现,在含水量为35%~60%的土壤中,较高的氧分压有利于微生物的硝化作用,硝化作用成为产生N2O 的主要过程;而在含水量大于70%的土壤,反硝化作用则占据主导地位. 通常在土壤孔隙水含量大于60%的土壤中N2O 排放最快[39]. 此外,水稻土在不同含水量下的N2O 吸收量不同,N2O 吸收量随土壤含水量(20%~70%)的增加呈指数增长.
综上,土壤水分通过改变土壤孔隙度、氧气利用率、土壤微生物活性、底物浓度及气体分子扩散能力来综合影响稻田水土界面温室气体的排放. 稻田水土界面的碳源是温室气体的主要前提,微生物介导碳源转化对CO2和CH4的排放存在底物竞争,水土界面微环境变化是影响二者排放的重要因素,而微环境改变也是制约N2O 排放的关键因素. 以往研究主要针对其中一种或两种气体研究,鲜少针对三种气体的研究,在复杂的稻田生态系统中,三种温室气体的总体排放是制约温室增温潜势的关键. 基于生态系统对三种温室气体的总体定量研究是控制稻田温室气体排放的关键.
2.3 温度
稻田温室气体的排放量因环境温度的不同而表现出显著差异性. 研究表明,我国热带、亚热带和温带试验稻田(南部、中部和东北地区)的CH4累计排放量分别为63.4、161 和36.9 kg/hm2[40],N2O 累计排放量分别为1.3 kg/hm2[41]、0.91 kg/hm2[42]和0.30 kg/hm2[43].温度是影响稻田温室气体排放的关键因素. 较高的土壤温度可以增强作物的根系呼吸,促进土壤中有机质的分解,提高土壤中微生物的活性,加速土壤矿化,从而促进CO2排放[44]. 此外,气温的升高可促进土壤无机盐(SIC)和碳酸盐的溶解[21],由此增加了其对土壤CO2排放量的贡献. 温度对CH4排放的影响主要归因于温度改变了产甲烷菌和甲烷氧化菌的组分和活性. 甲烷产生菌的最适温度为35~40 ℃,而CH4氧化的最适温度为25~35 ℃[45]. Walker 等[46]通过原位自然增温试验探索土壤微生物对温度变化的响应机制,结果表明,微生物的温度敏感性和基质消耗程度共同影响土壤碳损失. 已有研究证实,土壤温度每升高1 °C,土壤将多排放0.091 g 碳[20]. Steinkamp 等[47]发现,在0~10 ℃的温度范围内,甲烷氧化对土壤温度具有积极的响应趋势,同时温度升高可通过增加气体扩散率,增加O2利用率以促进甲烷氧化. 土壤温度还可通过影响硝化作用与反硝化作用的相关微生物活性而影响N2O 的排放. 土壤中N2O 的产生有生物和非生物途径,高温会刺激微生物活动. Cui 等[48]通过对假单胞菌(Pseudomonas mandelii)进行培养,发现其反硝化活性与土壤温度成正比. 硝化作用的微生物活动适宜的土壤温度范围是15~35 ℃,而在低于5 ℃或高于40 ℃的土壤温度下,硝化作用会被抑制.反硝化微生物则需要适宜土壤温度为5~75 ℃. 土壤温度升高时,反硝化基因(nirS)的表达和N2O 的排放率都较高. 同时,土壤温度会强烈影响硝化速率和氨氧化细菌(AOB)的活性. 此外,高温对非生物途径产生N2O 的影响显著,50 ℃时的非生物反硝化效果最强,N2O 排放量最高[19].
温度的升高促进了土壤有机质的分解,进而影响土壤微生物的代谢活动和生理功能,如碳利用效率、微生物群落变化等来综合影响温室气体的排放. 目前,关于温度对稻田温室气体排放影响的研究中,绝大多数结果都表明温度的升高能够促进CO2、CH4排放,而N2O 则表现出较复杂的排放规律.
2.4 氧化还原电位
土壤氧化还原能力是产生CH4的关键因素之一.在土壤CH4产生过程中,土壤微生物还原过程依次使用NO3—、Mn4+、Fe3+、SO42—作为电子受体,还原程度不断增强,同时由于还原作用引起pH 的升高和氧化还原电位的降低,进而促进CH4释放. 通常CH4生成的起始氧化还原电位为—160~—150 mV[49]. 土壤氧化还原电位不仅影响CH4的生成,还影响CH4在植物中的气体传输. 较高的土壤氧化还原电位会降低通气组织的形成和产甲烷菌的活性[50]. 当土壤氧化还原电位由—200 mV 降至—300 mV 时,CH4的生成量可提高10 倍,其净排放量的提升幅高达17 倍[51]. 土壤呼吸作用与氧化还原电位呈显著正相关,呼吸作用受土壤氧化还原电位的间接影响而改变土壤CO2的排放[25]. 当氧化还原电位为—250~—150 mV 时,CO2排放明显增加[52]. 然而,土壤氧化还原电位与土壤CO2排放深层相互作用机制研究依然有限. 土壤氧化还原电位可以通过影响微生物介导的反硝化作用而影响土壤N2O 排放. 例如,在中度还原至弱氧化条件下(氧化还原电位大于250 mV),真菌反硝化占主导地位,而在强还原条件下(氧化还原电位小于—100 mV)细菌反硝化占主导地位[53]. 在土壤悬浮液中,400~500mV 的氧化还原电位值有利于氨化,而在较高的氧化还原电位值(500~650 mV)下,硝化作用占主导地位[54].
2.5 植物
植物在生长过程中可通过改变土壤养分组成及含量、植物自身生物活性及微生物群落组分等影响温室气体的排放. 在稻田中,植物为土壤和根际微生物提供基质,30%~60% 的净光合碳分配给根系,其中40%~90%的部分被转移到土壤中,这种地下碳流为温室气体的产生提供了反应底物. 不同植物的自身生物性状(如生物量、叶面积指数)存在较大差异,不仅影响着植物自身的生长和代谢活动,还影响着生态系统中凋落物和土壤有机质的输入,从而影响温室气体的排放. 不同品种水稻对应的土壤N2O、CH4排放量和全球增温潜势不同,且与水稻的地上、地下生物量显著相关[55]. Reichstein 等[56]发现,土壤呼吸与植被最大叶面积指数、叶片生物量、凋落物质量均呈正相关. 此外,土壤微生物作为温室气体排放的主要参与者,依据植株种类的不同而表现差异[57]. Hussain[58]研究发现,杂交稻与常规稻在根际微生物数量和组成方面存在差异,杂交稻拥有更多氨氧化细菌和甲烷氧化菌基因,而反硝化细菌基因拷贝数则相对较低.
不同植物物种间的养分吸收、生物性状、生理代谢活动等特性都存在着较大的差异,这些差异影响着稻田生态系统的氮碳循环,进而影响土壤中养分的分布. 此外,水稻通过影响水土界面局部生境而改变微生物群落分布,最终影响稻田温室气体排放.
3 温室气体减排措施建议
稻田生态系统中土壤特性因素(如理化结构、化学组分和养分含量、微生物群落)受农艺管理措施影响显著,其农艺过程对水土界面的影响为稻田生态系统中温室气体的减排提供了思路. 据报道,调整水分管理、肥料管理、有机添加剂的管理等农艺管理措施可以减少水稻生产碳排放[6],为实现稻田增汇减排提供了理论依据.
3.1 土壤微生物调控
通过了解酶学和微生物学,设计调控土壤生化性质方案,从而控制与温室气体相关微生物的生理学,可以达到有效减缓温室气体排放的效果. 农作管理措施可以在一定程度上通过控制土壤酶活和微生物种群等以调控温室气体的排放,如添加生物炭、秸秆管理、控释肥料、硝化抑制剂和除草剂等. Das 等[59]研究发现,施用除草剂(即甲基苄磺隆和甲草胺)可降低土壤反硝化酶活性、微生物生物量碳和易矿化碳含量,并显著减少N2O 和CH4的排放. 尿素的深埋施用可以通过在作物生长早期显著降低可矿化碳浓度和反硝化酶活性,从而有效减少8%~46% 的CH4和N2O 排放[60]. 秸秆还田配施化肥可以提高二磷酸核酮糖羧化酶活性,同时提高稻田土壤的固碳效率[61]. 调控微生物的种类和数量也是有效控制温室气体排放的手段. 土壤中添加解淀粉芽孢杆菌可显著减少氨氧化细菌丰度,抑制硝化过程,同时增加N2O 还原菌丰度,反硝化过程增强,进而减少了50%的N2O 排放通量[9]. 通过可溶性有机物料与氮肥联合施用,可以降低土壤中革兰氏阳性菌和真菌的数量,从而减少了光合碳在有机质和微生物量中的分配. 这种施肥联合措施有助于调整土壤微生物群落结构,优化碳的分配方式,对土壤碳循环和养分利用具有积极的影响[62].Yuan 等[63]发现,通过改变管理制度,可以实现调整CO2同化细菌和藻类的丰度和多样性. 鉴于地杆菌在土壤氮循环的参与地位,可以通过施用地杆菌药剂诱导硝酸盐在反硝化过程中的快速转化,以实现水稻土N2O 的减排[30].
作为微生物聚集体的周丛生物,广泛分布于稻田水土界面,主要由藻类、细菌、真菌等生物部分与胞外聚合物、生物碎屑等非生物部分组成. 在稻田生态系统中,一方面,周丛生物的存在改变了水土界面的营养盐含量;另一方面,增加了产甲烷菌的群落大小,上调了产甲烷过程的关键代谢产物辅酶F420,促进了CH4的排放. 周丛生物增加了表层土mcrA和pmoA基因丰度,并增加乙酰辅酶A、丙二酰辅酶A 和3-羟基丙酰辅酶A 等3-羟基丙酸(3HP)代谢途径的中间代谢物[40],促进了稻田CO2的固定. 综上,周丛生物可以通过改变土壤表层营养物质的含量,土壤酶活和微生物群落而影响稻田温室气体的排放. 因此,调控周丛生物的生长对稻田温室气体减排具有良好的调控作用. 通过农事管理可有效改善水土界面组成和组分,有助于水土界面生物群落形成,可实现水土界面微生物群落、功能微生物结构和组成的调控,进而介导温室气体的排放. 然而,微生物群落结构复杂多变,其如何驱动温室气体排放依然有待进一步研究和明确,为科学农事管理、微生物精准调控提供参考依据.
3.2 土壤水分管理
水分管理是对稻田温室气体排放影响最显著的因素之一. 据统计,亚热带双季稻田CH4年均排放量(以C 计)为(42±2) g/m2[64],而旱作稻田CH4的年排放通量为—26.0~—29.7 g/m2[65]. 根据Chen 等[66]的Meta数据分析,旱作稻田和水稻田的年均N2O 通量(以N 计)分别为(4.69±4.62)和(5.89±3.23) kg/(hm2·a). 水分管理通过改变土壤通气状况来影响土壤微生物种类、数量及其活性,进而影响有机质的分解速率以及温室气体的生成速率. 与常规水分管理相比,一方面,有效排水使土壤通透性增加,氧化还原程度提高,土壤有机质分解速率加快,CO2排放增加[18];另一方面,有效排水可直接影响CO2在土壤水中的溶解量以及在土壤孔隙中的扩散速率,从而对CO2的排放量产生影响.
水分管理是影响稻田生态系统CH4排放的主要控制因素之一,为CH4的产生创造了必要的厌氧环境. 常规灌溉会减小表层土壤总孔隙度[67],破坏土壤团聚体结构[68],恶化土壤通气状况. 与常规水分管理相比,干湿交替[69-70]、季中排水[71]、控制灌溉[69]在不影响水稻产量的前提下,可减少温室气体的排放[6],其减排原理本质上是一致的,均通过改善土壤通气状况,升高氧化还原电位,促进CH4氧化菌活性,从而抑制了土壤中CH4的产生[72]. Zhang 等[73]利用同位素示踪试验发现,间歇性灌溉使稻田的季节性CH4生产潜力降低了45%,季节性CH4排放量减少了71%.
N2O 的排放与稻田土壤水分状况密切相关,改善水分管理措施是减少N2O 的潜在策略. 与常规灌溉相比,滴灌处理的N2O 排放量减少了55%~67%[74],应用小容量的灌溉措施可减少N2O 排放,改善水管理可以作为减少N2O 排放的有效工具. 因此,选择合适的灌溉方式对温室气体减排至关重要. 在施氮期间保持淹水,其余时期进行干湿交替灌溉可能有助于减少CH4和N2O 的排放[75].
3.3 土壤化学组成和养分管理
3.3.1 肥料管理
肥料管理是影响稻田生态系统温室气体排放的关键. 为提升肥料利用效率,我国已采取众多肥料管理策略,如测土配方、精准优化施肥、施用绿色有机肥、施用缓释肥料等. 减量优化施氮是一种可行的氮肥调控措施:一方面,减量优化施氮,减少氮肥施用量,即减少了氮流失损耗的基数;另一方面,较低的氮输入导致水稻与微生物之间的生存竞争,这有利于水稻对氮素的同化,从而降低N2O 的排放[76]. 研究[77]表明,优化施肥处理在减氮21.4%的条件下,产量与常规施肥处理持平,同时减少了水稻季12.6%的CH4排放和12.5%的N2O 排放. 相较于化肥而言,有机肥对土壤速效氮含量的增加速率相对滞后,抑制了反硝化酶的活性,降低了硝化电位,从而抑制了N2O排放. 有机肥和化肥的配合施用可通过影响土壤生化性状来改变参与CH4排放相关土壤微生物的结构.例如,施用高比例有机肥改善了产CH4土壤古细菌的丰度[78];Zhang 等[79]通过长期野外试验发现,施用有机肥显著减少了32.16%的N2O 排放;Linquist 等[80]报道指出,不同类型的缓控肥料可以显著减少N2O的排放.
肥料管理是稻田温室气体减排的有效措施之一.一方面,可以通过测土配方、减量优化肥料等,降低肥料损耗基数,从而实现减排;另一方面,延长肥料释放时间,使之与作物养分需求匹配,进而实现减少温室气体排放. 除此之外,肥料的施用改变了土壤微生物群落结构与组分进而影响温室气体的排放.
3.3.2 有机改良剂管理
将秸秆、生物炭等有机添加剂应用于水稻生态系统是保障粮食安全、减缓气候变化的潜在解决方案. 秸秆还田是水稻生产最为常见的农艺措施之一,可以有效增加土壤有机质含量,改良土壤结构,降低农业生产成本. 秸秆作为一种高C/N 资源,提供了大量产甲烷基质,导致土壤氧化还原电位显著降低,进而刺激CH4产生[81]. 与CH4相比,秸秆掺入有助于减少稻田N2O 的排放. 邹建文等[82]发现,稻季麦秸还田减少了19%的N2O 排放,这归因于高C/N 的秸秆使得土壤微生物固持土壤中大部分无机氮,从而使得硝化与反硝化作用所需的氮反应底物减少,削弱N2O的产生[83]. 相较于常规秸秆还田,秸秆接种微生物菌剂还田不仅可实现温室气体减排,还可促进水稻生长. 微生物接种剂覆盖秸秆是实现稻田增产减排有效管理措施[84].
生物炭作为重要的可再生资源,具有富含碳的细颗粒和多孔结构以及持久耐分解的特性,有助于改善土壤结构,促进土壤碳固存[85],被广泛应用于农业生产[86]. 生物炭的施用可增加22% 的碳封存量,减少35%的N2O 排放[87]. 生物炭的具有较高的减排潜力:一方面,这归因于生物质炭较强的吸附性能可能会改变土壤中碳氮底物含量,从而降低N2O 排放;另一方面,高C/N 也可能是生物质炭降低氧化亚氮排放的原因之一. 生物炭还田可改变与CH4、N2O 相关的土壤微生物群落组成、功能基因(mcrA、pmoA、nosZ基因)丰度,进而显著影响温室气体排放[12]. 在酸性土壤中添加生物炭和氮肥会使土壤pH 升高,抑制真菌的反硝化作用,显著降低真菌的丰度,增加nosZ基因的丰度,增强N2O 还原酶的活性,促进细菌将N2O 还原为N2[88]. 生物炭还田改变土壤碳氮底物、氧化还原条件及微生物组分而显著减少CH4排放[12]. Liu 等[89]通过Meta 分析发现,生物炭显著降低了29%的基于产量尺度计算的温室气体排放强度(GHGI).
3.4 耦合技术联用
在实际的水稻生产中,通常采用水肥管理和其他农业工程技术相结合的方法来稳定产量和减少温室气体排放,其中水肥耦合技术是关键所在. 研究发现,中国稻田在减少15.7%的施氮量条件下不会降低水稻产量,而浅水灌溉和适当施肥相结合可以减少34.3%的温室气体排放,同时提高1.7%的水稻产量[90].此外,滴灌结合滴灌管埋深通过影响土壤理化特性和CH4氧化基因拷贝数而增加土壤CH4累积吸收量[91]. 徐丹[92]通过研究不同水肥模式下的寒地黑土稻田温室气体排放强度发现,控灌模式下的平衡施肥处理的水稻产量最高,且综合温室效应和温室气体排放强度最低. Tian 等[93]对中国水稻种植条件下温室气体排放与产量的平衡关系进行了评估,结果表明,通过采取集中排水和平衡施肥的双重管理措施,可以在保证产量的情况下显著减少CH4和N2O 的排放.这种综合管理方法有助于实现水稻种植的可持续发展,同时对气候变化的缓解也有重要意义[94].
4 结论与展望
4.1 结论
a) 稻田水土界面温室气体排放的影响因素众多,物理因素、化学因素及生物因素三者相互作用,并共同调控稻田温室气体的排放. 在水土界面微环境下,界面碳源是温室气体产生的物质基础,土壤微生物介导碳源转化受水土界面微环境变化(土壤氧化还原电位、土壤水分、温度等)的调控而直接或间接影响稻田温室气体的排放.
b) 通过调整水分管理、肥料管理,添加有机添加剂,调控土壤微生物及耦合技术联用,可有效实现温室气体减排. 其中,土壤微生物调控通过调控土壤微生物群落组分、活性及代谢产物等影响温室气体排放,微生物有助于水土界面物质组成改善,且可形成好氧-缺氧的微生态系统,在减排方面具有良好的潜势,是一种具有未来发展潜力的减排措施.
c) 土壤微生物是稻田温室气体的源和汇. 为减少稻田温室气体排放,了解土壤微生物介导的稻田温室气体产生过程非常重要. 土壤呼吸、甲烷生成、甲烷氧化、硝化和反硝化等微生物过程参与了稻田温室气体循环. 通过添加解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、促进甲烷氧化菌活性、探索含有N2O 还原酶基因的固氮菌等微生物群落,可以减少温室气体的排放. 因此,土壤微生物调控是减缓稻田温室气体排放的有效手段之一.
4.2 展望
稻田土壤温室气体排放的内因与外因是当前研究热点,其研究逐渐由宏观环境转向微观条件影响、由单因素影响向多因素交互作用转变,尽管如此,依然需要更多的研究进一步深入解析土壤微生物介导稻田温室气体产生及减排机理等系列问题. 根据当前研究现状的不足进行综合分析,对未来的土壤微生物与碳氮循环研究提出以下展望:
a) 由于微生物驱动温室气体排放的理论机制复杂性、科学技术手段的有限性,稻田温室气体排放微生物介导机制尚不明确,且众多过程局限于实验室研究,野外实地长期监测依然不足,野外大尺度稻田微生物群落结构组成研究较少,导致稻田生态系统水土界面微生物群落与稻田生态系统功能关联性较为薄弱,限制了稻田温室气体的定量研究. 未来研究应依托分子生物技术,从室内试验拓展到区域尺度,利用土壤微生物生物地理学理论和方法,深入解析稻田土壤微生物时空分布特征及微生物群落构建机制,揭示稻田土壤微生物分布与生态功能之间的关联机制.
b) 农事管理介导微生物调控相关研究及其量化模型尚处于起步阶段,相关稻田温室气体排放模型与实际情况存在显著差异,如何通过农事管理实现微生物调控其响应关系有待明晰,且管理措施如何定量影响微生物过程亟待明确. 未来研究应倾向于关注稻田水土界面土壤微生物对温室气体排放的影响及其在农事管理下的响应过程,明确农事管理与土壤微生物的交互关系,这对推动稻田温室气体的微生物介导和调控机制具有显著意义.
