盐田虾蛤混养模式下硬壳蛤的生长、水质及浮游植物特征分析
2023-12-19马东怡郭永军李永仁石八军张中华
马东怡,郭永军,李永仁,石八军,张中华
(1.天津农学院 水产学院 天津市水产生态与养殖重点实验室,天津 300392;2.天津市海升水产养殖有限公司,天津 300200)
我国沿海盐田总面积为33.7万hm2[1],为综合利用初级盐田资源,天津市于2001年在初级盐田进行对虾粗放试养,既稳定盐田生态平衡,提高制盐质量,又可以带来额外的经济效益[2]。经过几年的养殖筛选,发现南美白对虾具有广温广盐、抗病能力强及生长速度快等特性,已成为天津地区初级盐田的水产品主要养殖品种。初级盐田水体盐度较高、天然饵料较丰富,使所养殖的对虾(盐汪子虾)肉质紧实鲜美,深受消费者喜爱。但近年来为追求经济效益,养殖者盲目增加养殖密度和投饵量,导致水体富营养化,藻类大量繁殖,溶解氧含量下降,易造成对虾缺氧。初级盐田单池面积大,难以通过泼洒药物等措施改善养殖水环境,可引入滤食性贝类,开展虾贝生态立体混养模式,可有效防治水体富营养化。
生态混养即多营养层级复合养殖,通过搭配养殖不同生态位的水产种类,使物种之间互利共生,同时又可以维持养殖池内的生态平衡[3]。在对虾养殖水体中,饵料投喂会造成水质过肥,而滤食性贝类能够摄食养殖水体中的浮游植物和有机碎屑等,避免藻类大量繁殖,起到稳定水质的作用。
硬壳蛤(Mercenaria mercenaria)又称“美洲帘蛤”,原产于美国,1997年由中国科学院海洋研究所张福绥院士等首次引入中国[4],其肉质鲜美,营养丰富,畅销国际市场。研究表明,硬壳蛤对环境具有较强的适应能力,在沙质、泥沙质、沙泥质底质中均能存活和生长,耐受温度范围可达-2 ℃~35 ℃,适宜温度范围为18 ℃~28 ℃,耐受盐度范围为13~48,最适生长盐度范围为18~36[5],是一种适宜在初级盐田养殖的贝类。
2021年4月至2022年8月,在天津地区初级盐田开展了南美白对虾与硬壳蛤等的生态混养试验,监测并分析养殖水质理化因子与浮游植物状况,以期为初级盐田的高值化综合开发提供参考。
1 材料和方法
1.1 试验材料
试验用初级盐田位于天津市滨海新区大港沿海天津恒仟水产养殖有限公司,水源来自于天津市滨海新区大港发电厂的海水冷却水,平均水深常年保持在1.5~1.8 m。年盐度变化范围为28~40,pH 8.48~8.85,盐田面积为66.67 hm2,硬壳蛤投放在沿岸6.67 hm2范围内。2021年4月4日投放硬壳蛤苗种,规格为2 400粒·kg-1,在6.67 hm2投放区内投放密度为66 700粒·667 m-2,折合整个养殖水域放养密度为10粒·m-2。分别于2021年4月和2022年4月底投放2万尾·kg-1南美白对虾苗种,投放密度为3万尾·667 m-2,投放时南美白对虾体长约1.5 cm。当南美白对虾体长长至约4 cm时,每日投喂对虾饲料2次,日投喂量为南美白对虾体质量的3%~7%。2021年共投喂对虾饲料61 500 kg,2022年共投喂对虾饲料66 000 kg。对虾饲料中粗蛋白质≥42.0%,粗灰分≤15.0%,水分≤12.0%,粗纤维≤6.0%,粗脂肪≥5.0%,赖氨酸≥2.2%,总磷≥0.8%,钙0.5%~3.0%,氯化钠0.5%~3.0%。养殖盐田中还有天然生长的脊尾白虾和鰕虎鱼。
1.2 试验方法
试验时间为2021年4月—12月和2022年4月—8月,每月于试验初级盐田固定位置采集硬壳蛤50粒,监测其生长情况,测定壳长与湿质量(因处于新冠病毒感染暴发期,2022年1—3月未采样)。每15 d采集表层海水(10 cm深处)样本测定水质理化指标,包括盐度、pH、溶解氧、化学需氧量、氨氮、亚硝酸盐氮、硝酸盐氮及总磷含量8个指标,检测方法见表1。镜检浮游植物丰度、种类。
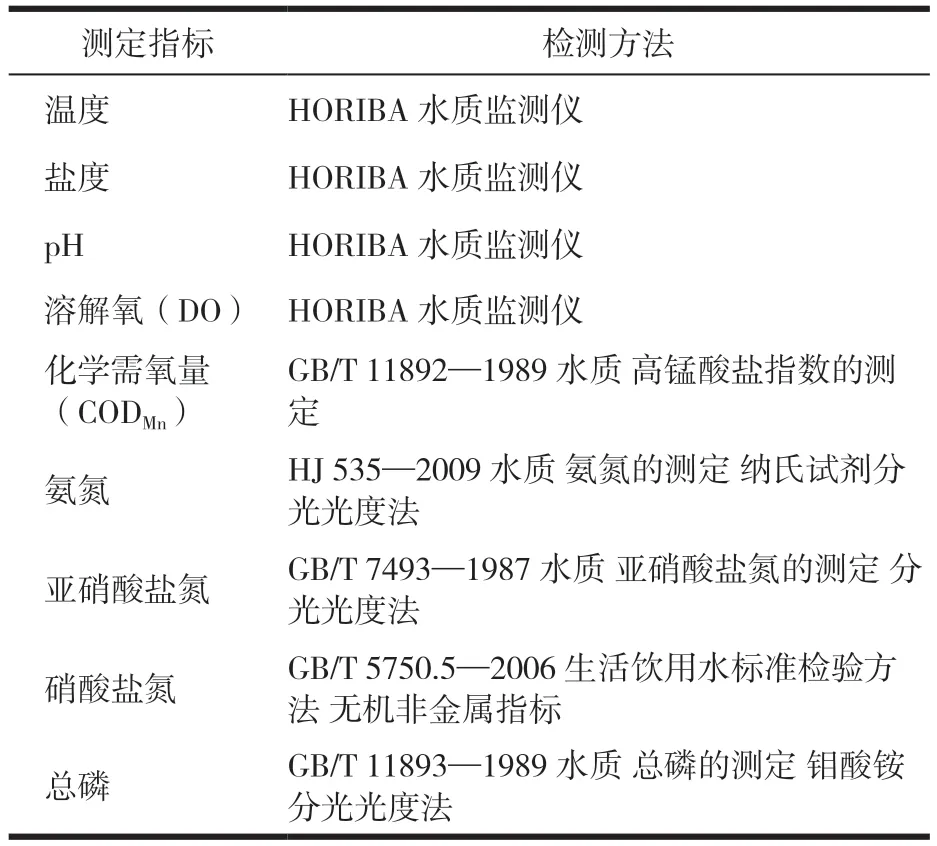
表1 水质样本的测定指标与检测方法
1.3 数据分析
对浮游植物丰度、种类数、主要类群、优势种属进行分析。
浮游植物丰度采用浓缩计数法,计算公式为:
物种优势度(Y)计算公式为:
其中,N′为浮游植物丰度(cell·L-1),n为取样计数所得的细胞数目(个),V′为水样浓缩体积(mL),V为水样浓缩前体积(mL),V″为取样体积(mL),N为样品的总个体数,ni为第i种的个体数量,fi为第i种个体在采样点中出现的频率。优势度Y≥0.02的种类,判定为优势种属。
采用Canoco 5.0软件对浮游植物优势种属与水质因子的关系进行RDA分析[6]。
2 结果与分析
2.1 硬壳蛤的生长与存活情况
硬壳蛤于2021年4月4日投苗,2022年9月1日开始为期15 d的收获。经过17个月的养殖,硬壳蛤平均壳长由13.95 mm长至51.83 mm,平均体质量从0.50 g增至45.07 g,成活率达89.99%。
2022年硬壳蛤在6.67 hm2投放区内单产为2 696.75 kg·667 m-2,折合整个盐田面积6.67 hm2单产为269.68 kg·667 m-2。2021年南美白对虾单产为51.3 kg·667 m-2,2022年南美白对虾单产为55.0 kg·667 m-2。2021年脊尾白虾单产约为10.0 kg·667 m-2,鰕虎鱼单产约为6.5 kg·667 m-2。2022年脊尾白虾单产约为7.50 kg·667 m-2,鰕虎鱼单产约为5.0 kg·667 m-2。
2.1.1 第一个快速生长阶段 2021年4月4日—7月中旬,养殖区水温为14.68 ℃~30.02 ℃,温度较适宜硬壳蛤的生长,其生长速度较快。
4月4日—5月中旬,硬壳蛤平均壳长从13.95 mm长至22.40 mm(图1),月相对增长率达到60.57%(图2);平均体质量从0.50 g增加至3.08 g,月相对体质量增加率达到515.00%。
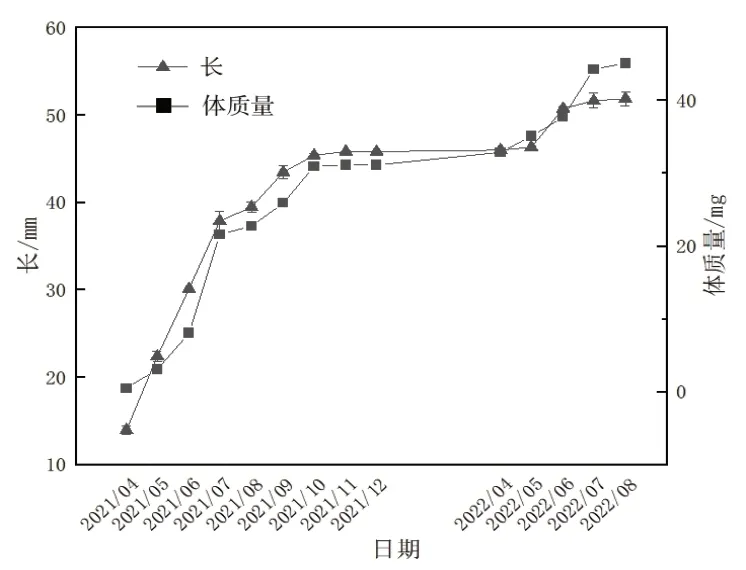
图1 硬壳蛤生长情况
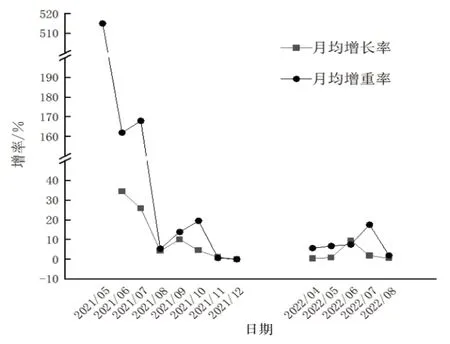
图2 硬壳蛤月均体质量增加率和生长率
5月中旬—6月中旬,硬壳蛤生长速度减慢,平均壳长从22.40 mm长至30.11 mm,月相对增长率降至34.42%;平均体质量从3.08 g增加至8.06 g,月相对体质量增加率下降至161.95%。
6月中旬—7月中旬,硬壳蛤壳长与体质量的增长表现不同步,此时进入硬壳蛤繁殖期,壳长增长缓慢,而随着性腺发育,硬壳蛤体质量增长很快[7];硬壳蛤平均壳长从30.11 mm长至37.86 mm,月相对增长率下降至25.72%;平均体质量从8.06 g增加至21.58 g,月相对体质量增加率升至167.91%。
2.1.2 第一个缓慢生长阶段 2021年7月中旬—8月中旬,由于养殖区水温达到32.19 ℃~33.48 ℃,温度较高不适宜硬壳蛤生长,导致硬壳蛤生长速度缓慢,平均壳长从37.86 mm长至39.45 mm,月相对增长率下降至4.20%;平均体质量从21.58 g增加至22.74 g,月相对体质量增加率下降至5.35%。
2.1.3 第二个快速生长阶段 2021年8月中旬—10月中旬,随着养殖区水温下降到硬壳蛤生长的适宜温度17.87 ℃~27.68 ℃,硬壳蛤生长速度再次加快。
8月中旬—9月中旬,硬壳蛤平均壳长从39.45 mm长至43.43 mm,月相对增长率升至10.10%;平均体质量从22.74 g增加至25.88 g,月相对体质量增加率上升至13.81%。
9月中旬—10月中旬,养殖区水温继续下降,硬壳蛤壳长与体质量的增长再次表现不同步,体质量增长很快,开始蓄积能量为越冬做准备。硬壳蛤平均壳长从43.43 mm长至45.37 mm,月相对增长率下降至4.46%;平均体质量从25.88 g增加至30.92 g,月相对体质量增加率上升至19.48%。
2.1.4 第二个缓慢生长阶段 2021年10月中旬—12月中旬,养殖区水温继续下降,温度范围为3.10 ℃~14.37 ℃,低于硬壳蛤生长的适宜温度,硬壳蛤生长速度减慢。
10月中旬—11月中旬,硬壳蛤平均壳长从45.37 mm长至45.79 mm,月相对增长率下降至0.93%;平均体质量从30.92 g增加至31.11 g,月相对体质量增加率下降至0.63%。直至12月中旬,养殖区水温降至9 ℃以下,硬壳蛤停止生长,进入休眠状态。
2.1.5 第三个快速生长阶段 2022年4月中旬—7月上旬,养殖区水温升高,温度范围为15.58 ℃~30.5 ℃,硬壳蛤进入到第二年度的快速生长阶段。
4月中旬—5月中旬,硬壳蛤平均壳长从45.96 mm长至46.30 mm,月相对增长率为0.73%;平均体质量从32.86 g增加至35.06 g,月相对体质量增加率为6.68%。
5月中旬—6月中旬,硬壳蛤平均壳长从46.30 mm长至50.70 mm,月相对增长率为9.52%;平均体质量从35.06 g增加至37.67 g,月相对体质量增加率为7.45%。
6月中旬—7月上旬,硬壳蛤平均壳长从50.70 mm长至51.62 mm,月相对增长率为1.80%;平均体质量从37.67 g增加至44.26 g,月相对体质量增加率为17.50%。
2.1.6 第三个缓慢生长阶段 2022年7上旬—8月中旬,养殖区水温高达32.7 ℃~34.9 ℃,抑制了硬壳蛤的生长,硬壳蛤生长速度减慢,平均壳长从51.62 mm长至51.83 mm,月相对增长率为0.42%;平均体质量从44.26 g增加至45.07 g,月相对体质量增加率为1.83%。
2.2 常规水质指标变化情况
硬壳蛤养殖过程中,pH变化范围为8.46~8.85,春夏季升高,冬季降低;溶解氧含量变化范围为3.67~9.86 mg·L-1,春夏季降低,冬季升高;化学需氧量含量变化范围较大,为1.80~18.15 mg·L-1,春夏季升高,冬季降低;氨氮含量变化范围为0~0.14 mg·L-1,春夏季升高,冬季降低;亚硝酸盐氮含量在整个养殖过程中浓度都比较低,变化范围为0~0.04 mg·L-1,春夏季升高,冬季降低;硝酸盐氮含量变化范围为0~0.15 mg·L-1,春夏季升高,冬季降低;总磷含量变化范围为0.27~0.49 mg·L-1,无规律性变化(图3)。
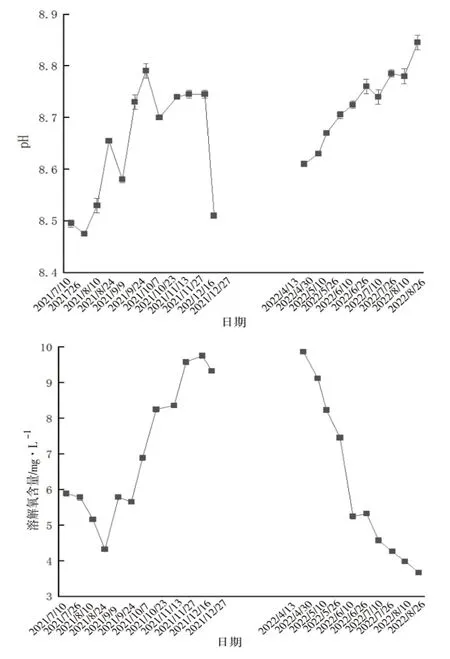
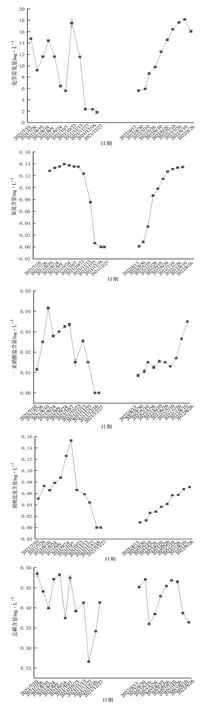
图3 常规水质因子变化情况
2.3 浮游植物群落特征分析
2021年4月—12月和2022年4月—8月,养殖区共鉴定出浮游植物7门29属(表2),其中硅藻门13属,占调查总种类的45%;绿藻门6属,占调查总种类的21%;甲藻门2属,蓝藻3门属,裸藻门2属,金藻门2属,隐藻门1属(图4)。
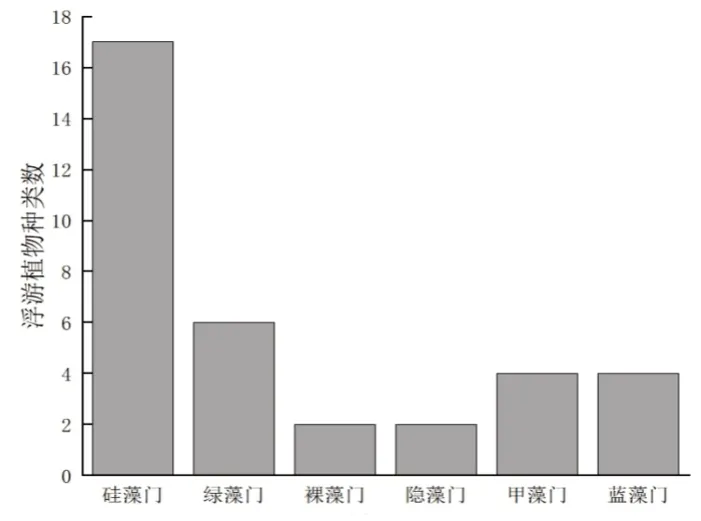
图4 浮游植物种类组成
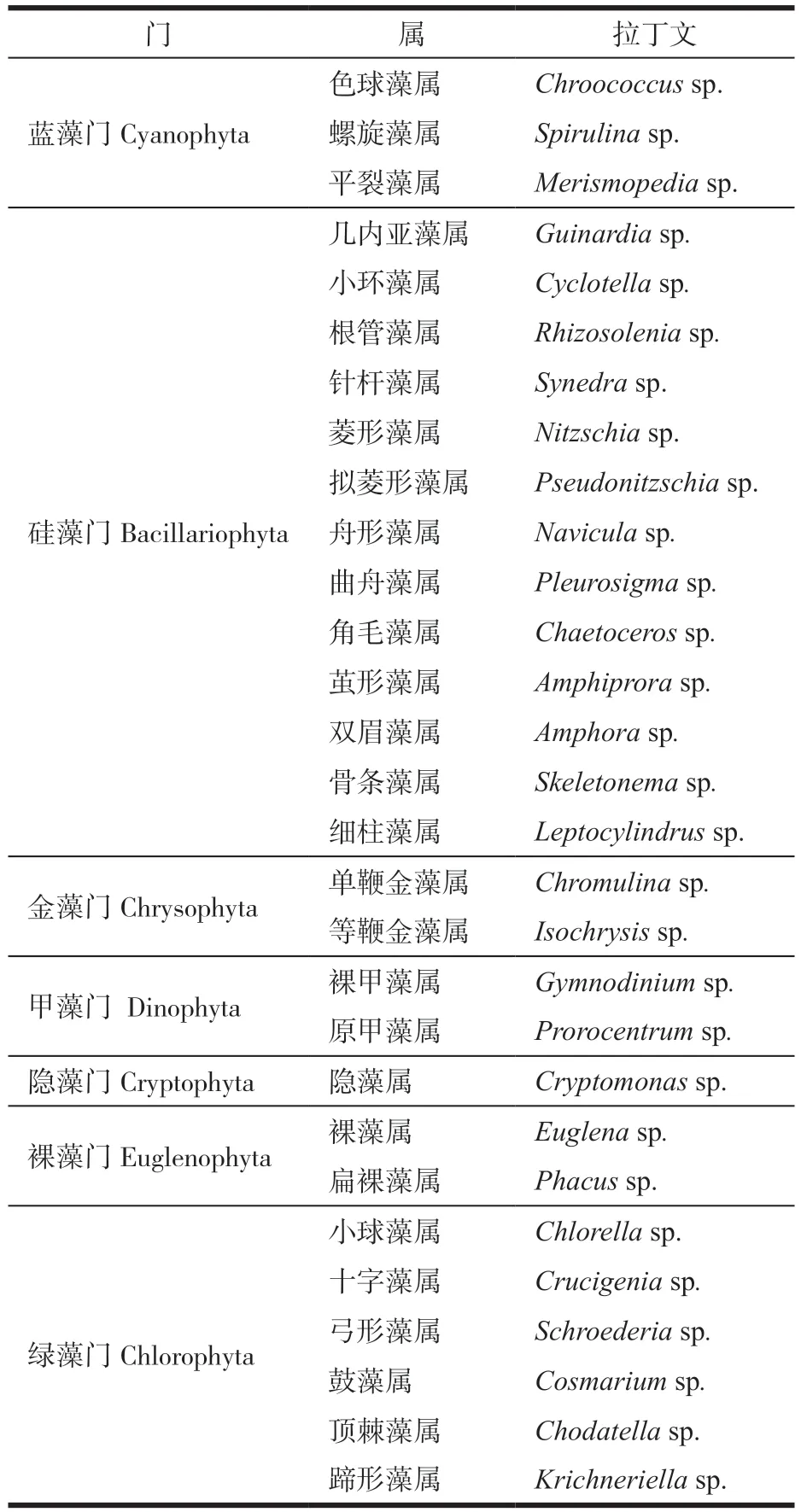
表2 浮游植物种类组成及拉丁文名
养殖区浮游植物丰度变化明显,变化范围为55.73 ×105~191.8×105cell·L-1(图5),2021年8月底浮游植物丰度最低,10月底浮游植物丰度最高,平均值为113.97×105cell·L-1。
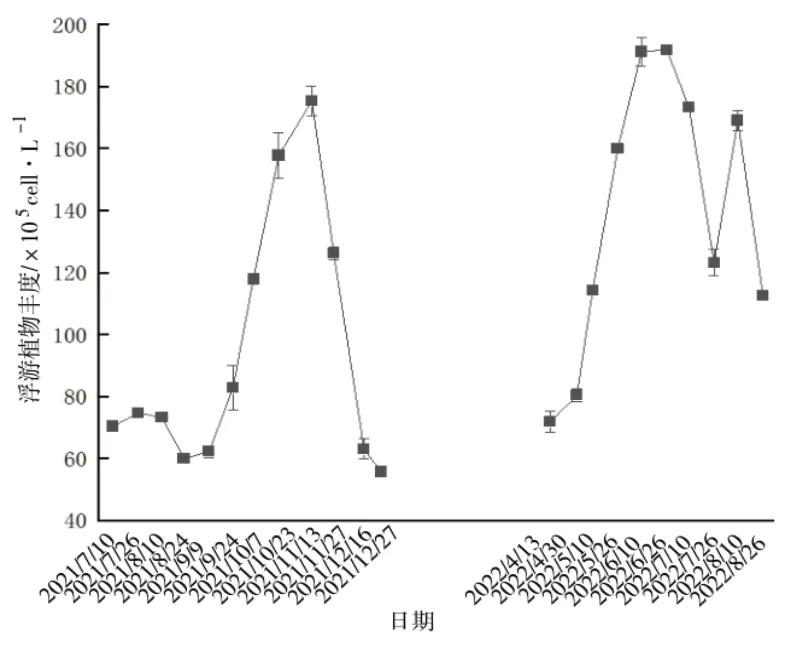
图5 浮游植物丰度时间分布
养殖区浮游植物优势种属(属级)共计7种(Y>0.02),其中硅藻门4种,蓝藻门、裸藻门、隐藻门各1种。每个月份的优势种属数量为1~3种(表3)。

表3 浮游植物优势种属变化情况
利用Canoco 5.0软件对7种浮游植物优势种属与常规水质因子进行RDA分析(图6)。结果表明,细柱藻属、小环藻属、小球藻属与水温、盐度、氨氮、总磷呈负相关关系;舟形藻属、色球藻属均与水温、氨氮、亚硝酸盐氮、硝酸盐氮、总磷呈正相关关系,并且受硝酸盐氮影响较小,与盐度呈负相关关系;裸藻属、隐藻属与盐度呈正相关关系,并且受盐度影响较小,与氨氮、亚硝酸盐氮、硝酸盐氮呈负相关关系。
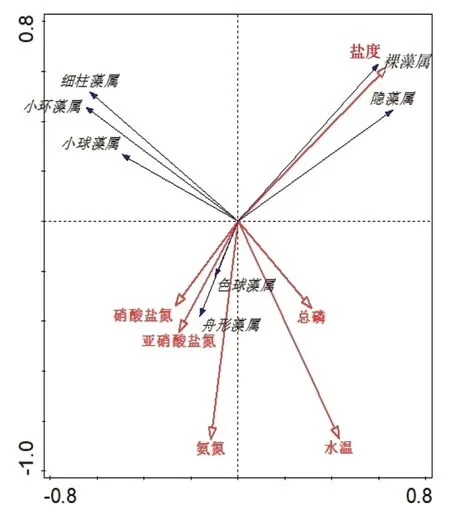
图6 浮游植物优势种属与常规水质因子的RAD分析
3 结论与讨论
3.1 水质理化因子的变化特征
初级盐田养殖池是一个小型半封闭生态系统,其面积大、水质理化因子会受气候、投饵、养殖密度等的影响,溶解氧含量、化学需氧量、无机氮(氨氮、亚硝酸盐氮和硝酸盐氮)含量的变化均与养殖进程相关。陈金玲等[8]在研究精养虾池主要生态因子变化特点时,也得出了类似结论。春季受温度和光照影响,浮游植物光合作用较强,水体中溶解氧含量较高,化学需氧量、无机氮含量较低;但随着夏季养殖的进行,生物耗氧量日渐增加,残饵粪便等有机质不断累积,使得养殖池内物质循环受阻,在微生物的分解下,溶解氧含量下降,化学需氧量、无机氮含量升高;秋季气温下降,养殖生物摄食代谢逐渐减少,溶解氧含量升高,化学需氧量、无机氮含量降低;冬季对虾养殖结束,硬壳蛤进入休眠状态,水体中溶解氧含量升至最高,化学需氧量、无机氮含量降至最低。
从整体来看,初级盐田养殖池无机氮含量均较低,氨氮、亚硝酸盐氮、硝酸盐氮含量均值分别为0.090、0.019、0.055 mg·L-1。杨世平和邱德全[9]在对虾高密度养殖试验中得出结论,即氨氮含量在养殖40~50d时出现峰值,亚硝酸盐含量在60~70d时才出现峰值,而本试验测定结果显示,氨氮、亚硝酸盐氮、硝酸盐氮含量的峰值基本出现在同一时间,这也说明混养滤食性贝类可以大量摄食藻类,可在一定程度上加快水体中的氮循环速度,从而保持良好的水质状况。
3.2 浮游植物的变化特征
盐田浮游植物群落结构简单,优势种单一。研究表明,海水虾池中硅藻占主要优势[10],这与本试验的研究结果类似。研究显示,虾池浮游植物丰度在9月初达到峰值[11],而本试验监测结果显示,9月份气温开始下降后,浮游植物丰度上升并在11月初达到峰值。秋季是硬壳蛤的快速生长阶段,会滤食大量浮游植物,11月开始水温逐渐降低到硬壳蛤的生长温度以下,硬壳蛤进入休眠状态、停止摄食,而水体及底泥中氮磷等营养物质还在释放,浮游植物持续繁殖,硬壳蛤滤食量却大幅减少,这可能是浮游植物丰度在11月初达到峰值的原因。上述两方面或证明在虾池中混养硬壳蛤确实可以起到滤食大量浮游植物、控制藻类数量、稳定水质的作用。
3.3 水质因子与浮游植物优势种的关系
浮游植物优势种属的组成易受环境因子的影响。根据RDA分析结果综合来看,水温和总磷是影响该盐田养殖池浮游植物分布的关键环境因子。陈宇炜等[12]在调研太湖梅梁湾藻类及相关环境影响因子时,也得出水温和总磷是影响藻类总生物量最为显著的环境因子的类似结论。磷是影响浮游植物生长最主要的营养元素,根据李夜光等[13]调查显示,浮游植物吸收利用的磷占全部磷元素的85%,同时磷也影响着微藻种类的交替。微藻对磷源利用存在差异,对磷源的利用特性不仅与磷源形态(正磷酸盐、有机磷等)相关,而且受藻类细胞磷酸酶活性的影响显著[14]。梁英等[15]在不同磷源对3种海洋微藻生长与种属间竞争影响的研究中得出,三角褐指藻对有机磷源的适应程度和利用效率远远高于盐生杜氏藻和米氏凯伦藻,其改变了3种海洋微藻种群的生物量比例,从而在种间竞争中获胜。水温在一定程度上可以影响浮游植物的种类组成[16]。水温从冬春季低温进入到夏季高温再逐渐回降的过程,往往也伴随着浮游植物群落结构的演替[17]。本试验还发现盐田浮游植物优势种属从冬春季节的细柱藻属、小环藻属、隐藻属转为夏季的舟形藻属、色球藻属。
