茄子萼下果色遗传分析
2023-12-19乔军刘婧潘东方李素文王利英
乔军,刘婧,潘东方,李素文,王利英
(1. 天津市农业科学院蔬菜研究所,天津 300381;2. 蔬菜生物育种全国重点实验室,天津市蔬菜遗传育种重点实验室,天津 300381;3. 天津科润蔬菜研究所,天津 300381;4. 天津市农业广播电视学校,天津 300202)
茄子是一种重要的蔬菜作物,在我国各地均有种植。据FAO 统计,2021 年我国茄子栽培面积为803 000 hm2,占世界总栽培面积的40.9%。近年来,各地茄子种植效益稳定,面积逐年增加,特别是设施栽培面积迅猛增长,茄子在生产中占有越来越重要的位置,已成为我国北方设施栽培的主要蔬菜种类之一。茄子喜光,适温条件下光饱合点约为40 000 lx,光补偿点约为2 000 lx[1]。保护地内光照较弱,常常导致紫茄皮色发白,俗称“白脸”或“绿脑门”,降低了茄子营养品质和商品性。近年来,国内外主要围绕茄子果色调控相关基因等方面进行研究,侧重于果皮性状。随着模式植物花青素相关基因的深入研究,影响茄子果实着色的相关基因也相继克隆,为揭示茄子耐弱光分子机理奠定了基础。
茄子果皮颜色是重要的农艺性状,颜色表达与光照条件密切相关。弱光下着色易变浅、斑驳,甚至过早失去光泽,严重影响果实商品性。在茄子的育种改良中,茄子果实颜色是一个重要的数量性状,而花青素苷是构成茄子果实的主要色素,属类黄酮化合物[2]。花青素苷属于类黄酮化合物的分支途径,主要存在于植物液泡中,是植物界最大的一类水溶性色素。至今已知的花青素苷超过550 种,但92%种类是由天竺葵色素、矢车菊色素、飞燕草色素、芍药花色素、矮牵牛色素、锦葵色素中常见的花青素衍生而来[3]。花青素苷和叶绿素共同作用影响茄子果色,花青素苷含量高则呈现为紫色,反之为绿色,色素含量因基因型而异。不同茄子材料果皮着色也在不同光照强度下表现出很大差异,一些紫茄材料萼片覆盖下的果皮不能正常着色,一般比不被覆盖的果皮部分色浅,甚至完全不着色,即呈现出果肉色。育种实践发现,萼下果色为紫的品种耐弱光能力强,果实在弱光下基本可以正常着色,而萼下果色为绿的品种耐弱光能力差,光照不充分时,果皮易出现斑驳、发白的现象。研究萼下果色调控基因,有助于耐弱光育种。
与果实其他部位相比,萼片下果皮始终被萼片覆盖,形成局部天然遮光环境。因此,萼下果皮是研究茄子光照诱导花青素等色素合成的理想材料。陈雪平[4]检测到3 个与萼下果色相关的主效QTL,分布在茄子第6 连锁群上。相丛超等[5]研究了六世代遗传和连锁SSR 标记,茄子萼下果色由2 对主基因控制。Toppino 等[6]在第5 和第10 染色体上定位到了2 个主效QTL。乔军等[7]利用2 个萼下果色差异显著的自交系Y5 和Y73,通过基因组重测序BSA 定位到7 个显著关联区间,分布于茄子第4、10 和11 号染色体。本试验通过构建六世代遗传群体,性状精准测定,采用多世代联合的数量性状分离分析方法,系统研究萼下果色遗传规律,为耐弱光育种提供有效参考。
1 材料与方法
1.1 试验材料
试验在天津市农业科学院武清创新基地进行(116.956 4°E,39.431 4°N),种植耐弱光高代自交系P1、不耐弱光自交系P2、杂交F1、回交B1和B2,以及F2代群体。P1和P2均为绿肉紫黑圆茄,中熟,生长势旺盛,叶片深绿,茎秆粗壮,单果质量500 g 以上。如图1 所示,P1萼片下遮挡部分果色仍为紫色,耐弱光;而P2萼片遮挡部分为绿色,不耐弱光。2021 年3月20 日直播于塑料大棚,P1、P2和F1各种植32 株,B1、B2和F2分别种植145、137、185 株,株行距为45 cm×60 cm,覆盖黑色地膜,参照常规栽培管理。

图1 萼下果色试验材料
1.2 试验方法
1.2.1 萼下果色性状调查 当对茄商品成熟后,拍照采集萼下果色性状,按照茄子种质资源描述规范[8]中绿、紫绿和紫色3 类萼下果色进行划分。制定本试验萼下果色比色板,相近萼下果色性状归类并赋值,见图2。
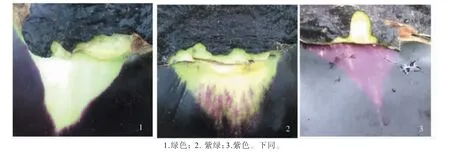
图2 萼下果色比色板
1.2.2 统计分析方法 采用盖钧益等[9]多世代联合的数量性状分离分析方法Ⅱ——P1、F1、P2、B1、B2和F2联合分析方法分析试验数据。通过极大似然估计法和IECM 算法对混合分布中的有关成份分布参数进行估计,根据AIC 准则和适合检验选择最适模型,通过最小二乘法估计相应一阶和二阶遗传参数。
2 结果与分析
2.1 亲本及杂交后代的萼下果色级值次数分布
2.1.1 P1、P2和F1萼下果色级值次数分布 由图3所示,级值为3,亲本P2表现为绿色,级值为1;F1萼下果色表现为紫绿,级值为2。组合亲本间萼下果色差异显著,F1萼下果色级值均介于亲本之间,符合六世代分离试验要求。

图3 F2 萼下果色分离
2.1.2 F2萼下果色级值次数分布 185 株F2代分离群体萼下果色的调查统计显示,55 株萼下绿,92株萼下紫绿,38 株萼下紫,符合数量性状遗传特点。B1代分离群体萼下绿的为17 株,萼下紫绿的为102株,萼下紫的为26 株。B2代分离群体萼下绿的为70 株,萼下紫绿的为49 株,萼下紫的为18 株。可以看出,F2代群体萼下果色出现明显分离,说明茄子萼下果色是数量性状,可能存在主基因和多基因的影响。
2.2 最优遗传模型的选择与检验
以六世代群体的萼下果色级值为基础,应用六世代分离分析软件计算24 个遗传模型的AIC 值。根据六世代联合分析原则,选择AIC 值较小的3 个遗传模型进行适合性检验,从中选出参数达到显著差异数量最少的模型作为最优模型。以萼下果色级值为基础,应用多世代分离模型软件计算AIC 值(表1),其中AIC值较小的3 个遗传模型分别是B-1、B-2 和E-4。
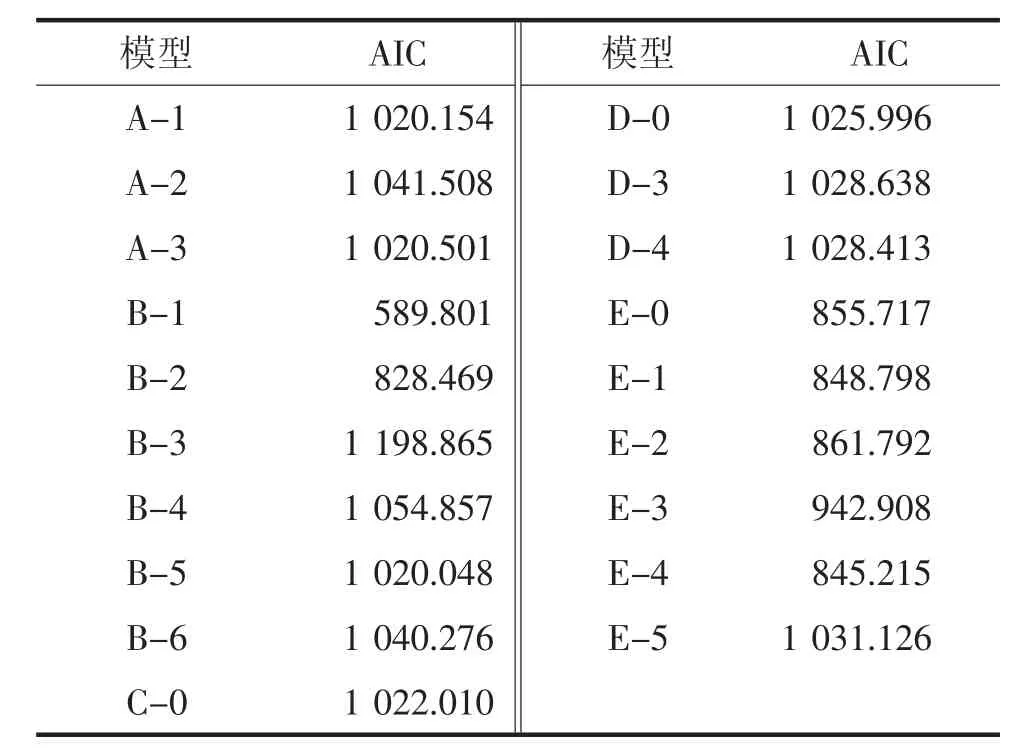
表1 各遗传模型的AIC 值计较
适合性检验结果表明,B-2 和E-4 模型分别有20、21 个统计量达到显著差异水平,而B-1 模型仅有19 个统计量达到显著差异水平,数量最少,可以用B-1 模型作为茄子萼下果色遗传的最佳遗传模型,即符合两对加性-显性-上位性主基因遗传模型(表2)。
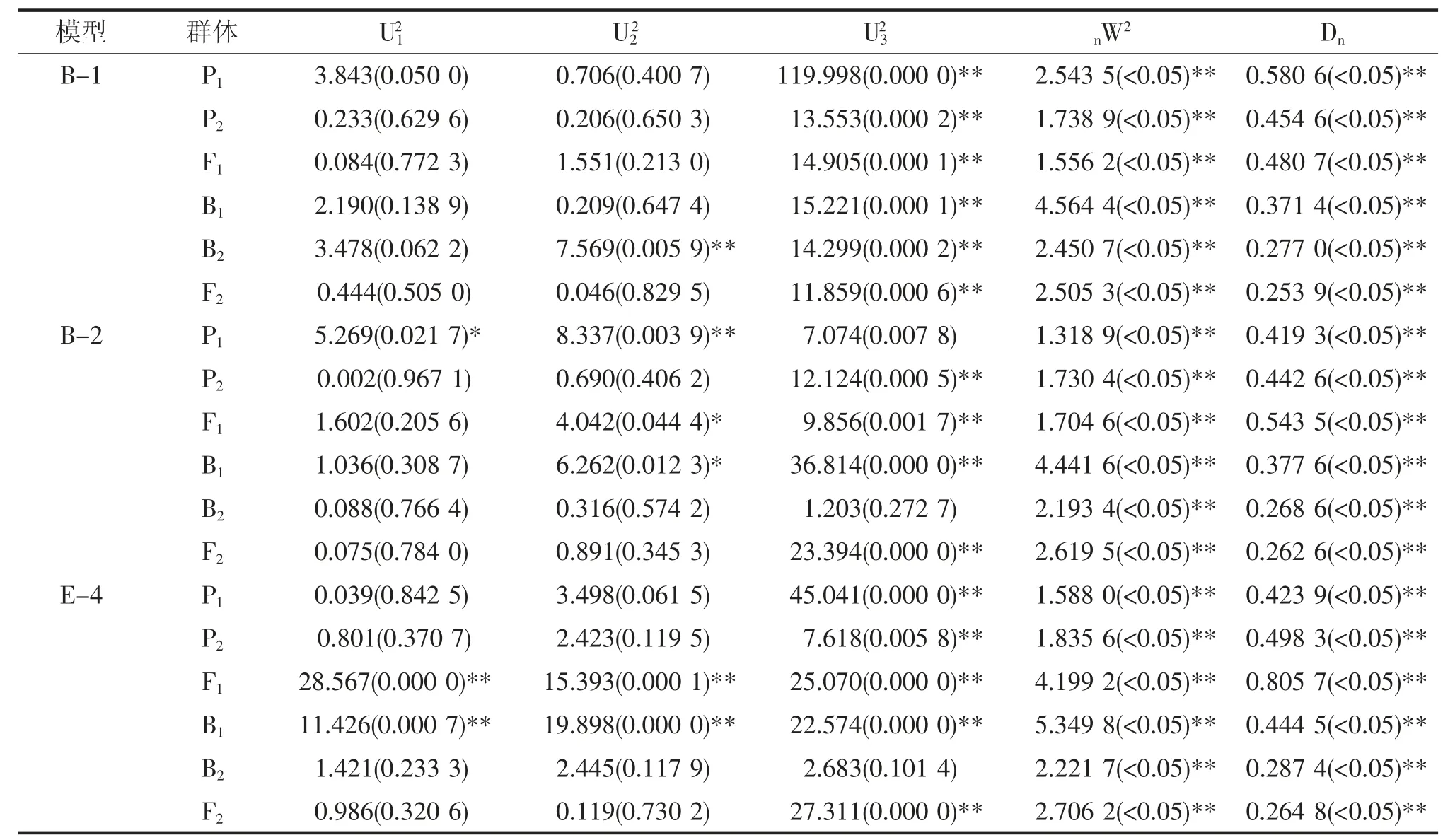
表2 遗传模型的适合性检验
2.3 最适模型的遗传参数估计
表3 是茄子萼下果色遗传的最佳遗传模型——B-1 模型的一阶遗传参数和二阶遗传参数。控制茄子萼下果色遗传的2 对主基因加性效应(da、db)分别为0.92 和-0.08,分别表现为正向和负向效应,2 对主基因均呈部分显性。第1 对主基因的显性效应(ha)为-0.93,显性度(ha/da)为-1.01,呈负向完全显性;第2 对主基因的显性效应(hb)和显性度(hb/db)分别为0.57 和-7.125,呈负向超显性。分析结果表明,第1 对主基因的加性效应和显性效应并重,第2对主基因以显性效应为主。
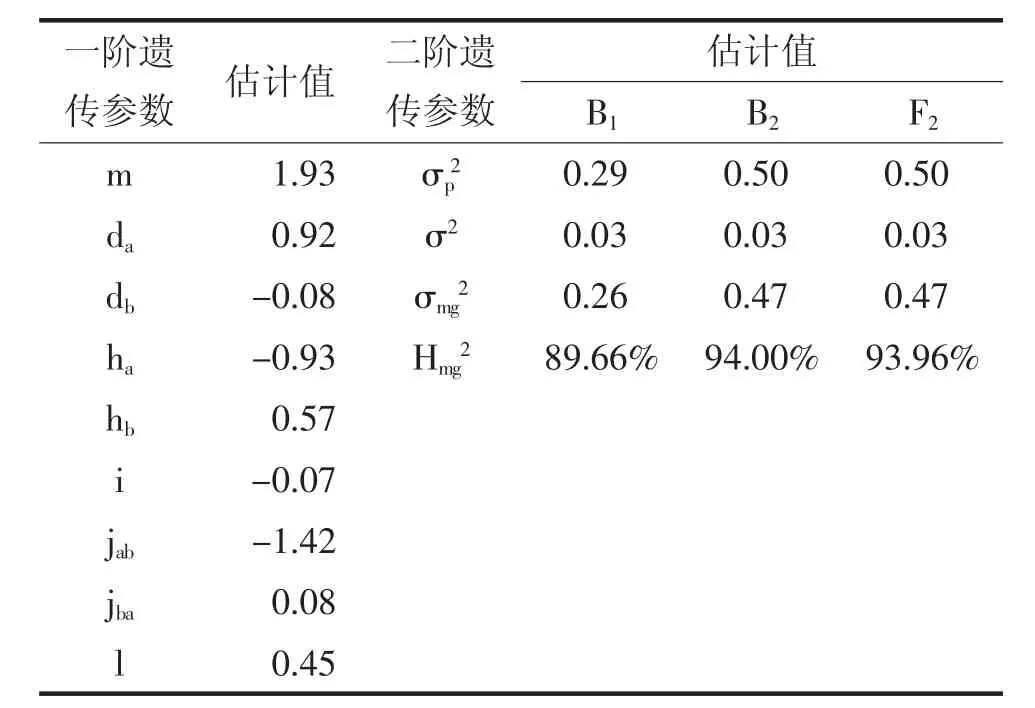
表3 六世代联合分析的萼下果色性状的遗传参数估计
在上位性作用中,2 对主基因的加性×显性(jab)和显性×显性(l)互作效应较大,显性×加性(jba)和加性×加性(i)互作效应一般。主基因遗传率(hmg2)在B1、B2和F2世代中分别是89.66%、94.00%和93.96%,选择效率较高。
结果表明:F2分离群体萼下果色级值呈正态分布,说明茄子萼下果色为主基因控制的数量性状;茄子萼下果色性状遗传符合2 对加性-显性-上位性主基因遗传模型(B-1 模型);主基因遗传率较高,为93.96%,可在育种早期进行选种。
3 讨论与结论
通过遗传分析发现,茄子萼下果色遗传受主基因影响严重,分离世代遗传率非常高。主基因遗传率在BC1、BC2和F2世代中分别是89.66%、94.00%和93.96%,非常有利于性状选择,在育种早期就可以快速固定该性状。
茄子的果皮颜色是最直观的园艺性状,主要由花青素和叶绿素共同决定,同时也受到各种环境因子的影响。Tatebe[10]研究推测了茄子果色相关的遗传因子,认为有6 个独立的遗传因子(C、P、D、G、Gv和Puc),其中C、P 和D 3 个主效基因共同调控花青素的形成,决定紫果皮颜色,G基因则和绿果皮颜色相关。Janick[11]报道,茄子果紫色性状是由2 对互相连锁且有不完全显性关系的基因支配的;Sambandam[12]研究发现,茄子果皮紫色对其他颜色都为显性;刘进生等[13]进一步研究发现,茄子果皮紫色性状是由2 对互相连锁且有不完全显性关系的基因控制,与本研究结果一致。
研究表明,茄子果实颜色属于主基因和多基因共同控制的数量性状[14]。目前已经在1、5、7、10 号染色体,以及多个连锁群上定位到了一些与果色相关的QTL 位点,表型贡献率最高达到了86%~93%[15-17]。相丛超等[5]研究萼片下果皮为浅紫色和绿色的亲本杂交后代的遗传分析,在F2群体中萼片下果皮颜色遗传符合2 对具有加性-显性-上位性作用的主基因和多基因模型,与本研究结果近似。
王世界[18]通过对紫茄和白茄果皮进行转录组测序,筛选到与茄子花青素生物合成相关MYB 转录因子3 个和结构基因8 个。孙保娟等[19]在茄子育种实践中发现,2 对上位性效应基因控制茄子果色遗传现象,应用SLAF-Seq 与BSA 关联分析技术,将控制茄子果色上位性遗传的2 个基因(D和P)分别定位到相应的2 个较小的区域。以上果色相关基因研究进一步证实,茄子果色控制基因的多样化和复杂性,遗传机制不是很清晰。
茄子萼下果色研究较少,本研究通过六世代遗传群体,借助数量遗传分析软件系统研究,深入揭示茄子萼下果色遗传规律,为品种选育提供理论参考和数据支撑,具有重要意义。
