液态发酵工艺对丝状真菌脂质积累的影响研究进展
2023-12-18薛福婷时文悦王秀文李少琦逄淑娴宋元达
薛福婷,时文悦,吴 琛,王秀文,李少琦,逄淑娴,宋元达
山东理工大学 农业工程与食品科学学院 (淄博 255000)
产油微生物发酵生产的油脂称为微生物油脂,与动植物油相似,是生产多不饱和脂肪酸的重要来源[1]。微生物油脂在脂肪酸组成上多数是偶数碳,特别是棕榈酸(C16: 0, PA)、油酸(C18: 1, OA)等,有些产油微生物也可以产生γ-亚麻酸(C18: 3, GLA)、花生四烯酸(C20: 4, ARA)、二十碳五烯酸(C20: 5, EPA)和二十二碳六烯酸(C22: 6, DHA)等多不饱和脂肪酸,在缓解关节肿痛、减轻炎症、治疗心血管疾病、抗癌等方面具有重要的生理功能[2,3]。
丝状真菌俗称霉菌,广泛存在于土壤、水体、空气及植物根系[4]。在液态发酵中的丝状真菌形态通常呈现出絮状、团状和球状,其形态上的多样性远复杂于单细胞的细菌和酵母菌[5]。对于丝状真菌而言,液态发酵具有周期短、易操作的优势;而且在脂质代谢过程中具有简单、易调控的优势[6]。由于丝状真菌的油脂含量高并含有丰富的功能性多不饱和脂肪酸而被广泛研究。本文主要总结近年来丝状真菌利用液态发酵工艺生产脂质的研究进展,旨在为丝状真菌规模化生产脂质提供参考和依据。
1 丝状真菌脂质积累概况
目前,能够用于生产油脂的微生物种类有藻类、酵母菌、丝状真菌及细菌,尤其以酵母菌和丝状真菌的真核生物居多,且含有丰富的脂肪酸种类[7,8],如表1。一些典型的丝状真菌,包括地半乳霉菌、毛囊霉菌、被孢霉和环核霉,其积累脂肪酸的能力已经得到了广泛的研究。卷枝毛霉(Mucorcircinelloides)、雅致枝霉(Thamnidiumelegans)、刺孢小克银汉霉(Cunninghamellaechinulata)、葡萄酒色被孢霉(Mortierellaviticola)、矮被孢霉(Mortierellahumilis)等丝状真菌能够积累GLA[7],如拉曼被孢霉(MortierellaRaman)和深黄被孢霉(Mortierellaisabellina)以葡萄糖为碳源时积累的GLA产量为1.0~1.6 g/L[9,10]。最早发现含有ARA的丝状真菌为青霉菌(Penicilliumcyanuam),其碳源为烷烃时积累的ARA含量为0.2 mg/g[11],拉曼被孢霉、深黄被孢霉和高山被孢霉(Mortierellaalpina)也是生产ARA的优良菌株。在产油微生物中,藻类和酵母菌含有丰富的EPA,终极腐霉(Pythiumultimum)、高山被孢霉等丝状真菌也具有积累EPA的能力,且高山被孢霉积累的ARA能够在常温下转化为EPA[12]。
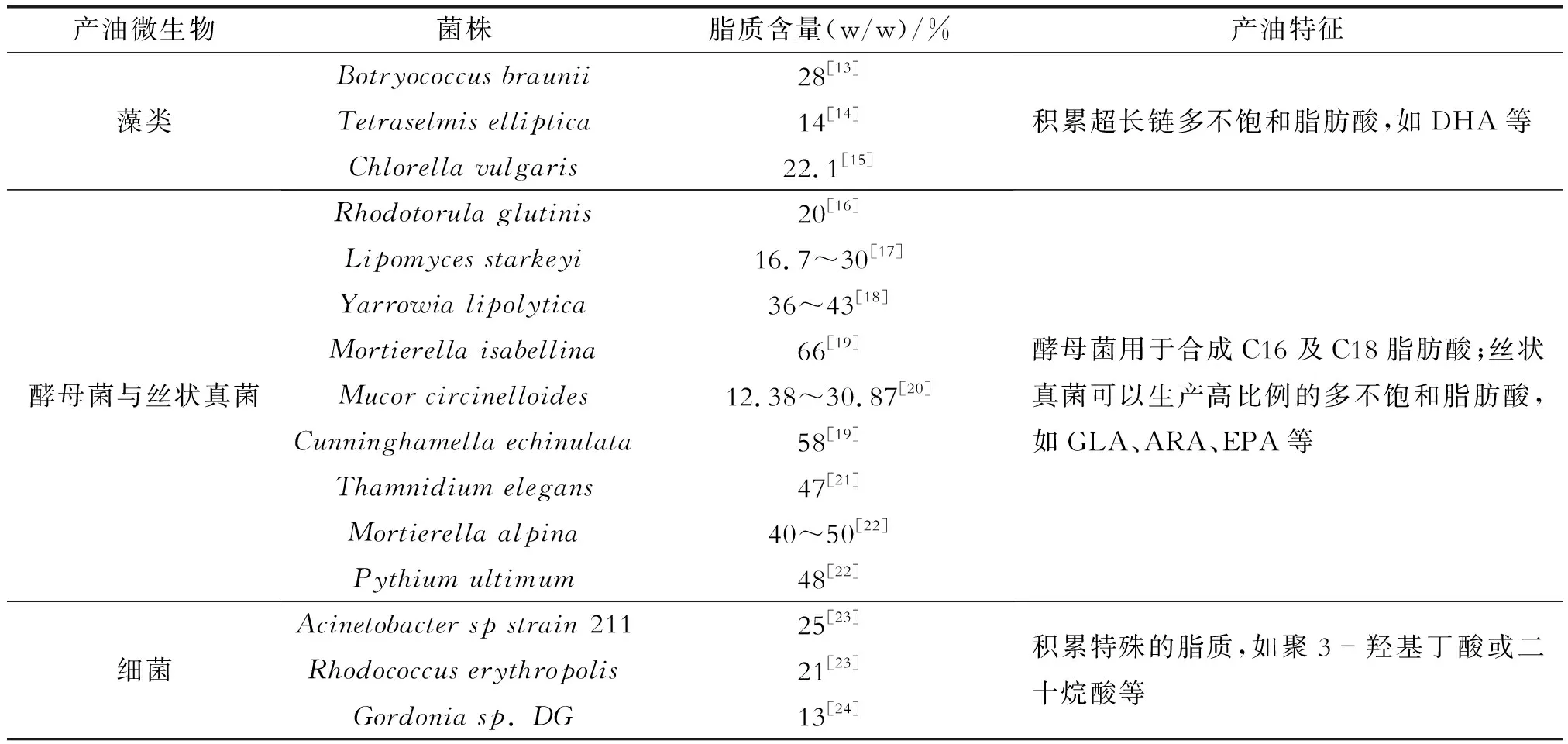
表1 丝状真菌和其他产油微生物在脂质含量和产油特征方面的比较
2 丝状真菌的脂肪酸合成过程
大部分丝状真菌主要以葡萄糖为碳源进行液态发酵,葡萄糖在胞液内经过糖酵解途径生成丙酮酸(Pyr),该途径中葡萄糖-6-磷酸脱氢酶(G6PDH)和6-磷酸葡萄糖酸脱氢酶(6PGDH)为脂质积累提供所需的还原力,部分丙酮酸进入线粒体(Mitochondrion)合成乙酰辅酶A(AcCoA),由于培养基中氮源耗尽使三羧酸循环(TCA)中柠檬酸(Cit)过度积累,从而被大量转运到细胞质,经柠檬酸裂解酶(ACL)催化后产生乙酰辅酶A,随后经乙酰辅酶A羧化酶(ACC)的作用生成丙二酰辅酶A(MaCoA),最终在脂肪酸合成酶(FAS)、延长酶及一系列脱饱和酶的作用下生成饱和或不饱和脂肪酸[25,26]。
3 培养基组分对丝状真菌脂质积累的影响
3.1 碳源
碳源在发酵过程中不仅为丝状真菌提供细胞骨架以及生长繁殖所需要的能量,还会影响其油脂积累能力,以葡萄糖为碳源使丝状真菌获得较高的油脂含量,且不同浓度的葡萄糖对油脂积累具有明显作用[27]。唐鑫等[28]研究20~80 g/L葡萄糖浓度对高山被孢霉发酵生产脂质的影响,在20~50 g/L浓度的菌体生物量和脂质含量逐渐增加,50 g/L的葡萄糖是其发酵的最佳碳源浓度,高于该浓度时脂质含量逐渐减少。同样地,严建平等[29]以不同浓度的葡萄糖为单一碳源探究卷枝毛霉EIM-10积累GLA的作用,发现卷枝毛霉在葡萄糖浓度为60 g/L时的生物量下降, GLA含量较低。以上研究说明,适宜的葡萄糖浓度有利于丝状真菌生长和提高脂质含量,但浓度过高可能使发酵液变得粘稠,导致菌丝体不能均匀生长,最后不利于脂质积累。
除葡萄糖外,许多其他碳源可用于丝状真菌的液态发酵。Li等[30]用海藻糖发酵刺孢小克银汉霉,GLA占总脂质含量为21%,高于使用葡萄糖作为碳源时的13%。刘尚杰等[31]采用5种碳源(果糖、葡萄糖、蔗糖、麦芽糖、乳糖)研究深黄被孢霉突变株A35-4的油脂含量,以果糖为碳源的油脂含量较高,达到73.84%。因此,不同碳源对丝状真菌的生长和脂质积累方面有重要作用,而木质纤维素、非谷物糖原料和商业废物等廉价底物也可以作为碳源,能够减少大量使用葡萄糖造成的高昂发酵成本,便于日后扩大生产脂质[32]。
3.2 氮源
氮源作为丝状真菌生长的基本营养物质,能够提供生长过程中所需的核苷酸、氨基酸和含氮代谢物等物质,氮水平的变化可能导致细胞代谢的巨大变化。顾晓颖等[33]研究有机氮源尿素和无机氮源硝酸钠对高山被孢霉的产脂的影响,该研究表明尿素更适合高山被孢霉生长和积累ARA。张天然等[34]采用蛋白胨、硫酸铵、酵母膏、尿素及硝酸铵这5种氮源发酵深黄被孢霉YZ-124,其中有机氮源酵母膏最适合菌体的生长和油脂积累。综合来看,有机氮源更利于丝状真菌积累脂质。
许多研究表明,氮限制和高碳氮比的培养基组分是脂质积累的有利条件,可以诱导丝状真菌产生脂质,抑制蛋白质和核酸合成[35]。Zhao等[36]改变卷枝毛霉发酵时的氮源含量,发酵50 h后发现卷枝毛霉在高氮培养基中的细胞干重明显高于低氮,但低氮中总脂肪酸含量是高氮的1倍多。常璐璐等[37]在高山被孢霉中也探究了碳氮比对脂质积累的影响,同样发现低氮内脂肪酸含量较高。因此,低氮有利于菌体积累油脂,而高氮则有利于菌体生长。
各种氨基酸作为氮源对丝状真菌的生长也具有差异影响,Tang等[38]用20种氨基酸为单一碳源研究卷枝毛霉的脂质变化,发现以酪氨酸为氮源发酵后的生物量和脂质含量最高,GLA含量较高,分别比酒石酸铵为氮源时高出36%、25%和72%。因此,氨基酸也成为理想的氮源。此外,一些含氮原料黄豆饼粉、玉米浆、花生饼粉等也可替代昂贵的氮源[39]。这些原料不仅成本较低,而且还能促进生物量积累和脂质产量,大大提高了丝状真菌生产多不饱和脂肪酸的经济效益。
3.3 外源添加物
由于脂肪酶的存在,一些外源油脂也可以促进丝状真菌脂质积累,如大豆油、大豆磷脂、植物甾醇、橄榄油等。李利军等[40]在发酵液中外源添加大豆油、菜籽油、亚麻籽油培养终极腐霉生产EPA,结果发现添加大豆油效果最佳,积累的EPA产量为429.89 mg/L;王艳霞等[41]研究了橄榄油添加量对卷枝毛霉油脂含量的变化,当发酵液中橄榄油浓度为15 g/L时,其生物量和油脂含量达到最高,因此外源油脂可以促进多不饱和脂肪酸的合成。
此外,添加外源底物也可以对脂质积累产生影响。Zhang等[42]在卷枝毛霉Mc-MT-2的发酵培养基中添加苹果酸后获得的脂质产率为5.12 g/L,与未添加苹果酸的对照菌株相比,提高了2.56倍。另外,在培养基中添加酶类激活剂也能刺激菌体内细胞大量合成油脂。所以,丝状真菌能够吸收不同的外源物质,其中外源油脂能够有效提高其脂质含量并增加脂肪酸种类。
4 液态发酵条件对丝状真菌脂质积累的影响
4.1 pH值
pH值是影响丝状真菌细胞生长和细胞内代谢物稳定性的最重要参数之一。细胞膜的电荷随着培养基中pH水平的变化而变化,进一步影响脂肪酸延长酶和脱饱和酶的活性,从而改变丝状真菌吸收利用营养物质的能力[43]。Mohamed等[44]研究pH值为4~8范围内对卷枝毛霉的影响,在次优级pH环境中卷枝毛霉生长较差,pH值为6更适合其生长和产脂,此时总脂肪酸含量和GLA含量分别为23%和15%。宋遥遥等[45]研究不同发酵条件对深黄被孢霉积累多不饱和脂肪酸的作用,其中pH值的增加使菌体生物量和油脂含量呈现先上升后降低的趋势,pH为6的发酵环境更适合深黄被孢霉的生长以及油脂的合成。梁凇等[46]探索了pH对刺孢小克银霉积累GLA的影响,当pH为5时,菌丝分散性较好且均匀分布,此时 GLA含量达到最高,为16.05%。总体来看,丝状真菌适合于偏酸性环境生长,pH值和培养基组分之间的相互作用可以影响丝状真菌利用有效营养物质的产脂能力。
4.2 温度
许多研究表明,温度是影响丝状真菌脂肪酸组成的主要因素,低温发酵可能提高脂肪酸的不饱和度,如合成ARA,EPA,DHA等长链多不饱和脂肪酸[47]。Zhang等[48]在深黄被孢霉M6-22中研究低温对其生产PUFAs的影响,温度从30 ℃降到15 ℃时,PUFAs含量从20.85%增加到31.04%,维持低温能有效促进PUFAs合成。夏继蓉等[49]也发现温度对高山被孢霉的菌体生物量及对菌体中PUFAs合成具有显著影响,25 ℃发酵获得的生物量最高,但15 ℃发酵获得的脂质含量最高。张玲等[50]对雅致枝霉发酵生产GLA进行探究,结果发现12 ℃有利于菌体的生长和GLA的积累。所以,不同菌株有着不同的最适生长温度,培养温度会影响细胞膜的流动性,低温促进大部分丝状真菌积累多不饱和脂肪酸,并影响脂肪酸不饱和双键的生成。
4.3 溶氧(DO)
溶氧是细胞生长和代谢的关键因素之一,改善发酵过程中的溶氧环境会对菌体生长和产物代谢水平造成极大的影响。 Hussain等[51]以150~300 r/min的搅拌速度控制卷枝毛霉发酵时的溶氧,在150~220 r/min的总脂质含量逐渐增加,而在220 r/min时的总脂质含量最高,超过该搅拌速度时的总脂质含量降低。因此,适宜的溶氧浓度将促进细胞内油脂积累,在提高溶氧的同时需避免剪切力作用,防止菌丝损伤。
氧气供应对于富含多不饱和脂肪酸的脂质积累至关重要,多不饱和脂肪酸合成过程是由需氧脂肪酸脱饱和酶及延长酶途径催化而成,该过程中氧消耗极高, 过量DO浓度已被证明在提高其脂肪酸不饱和程度方面发挥重要作用[52]。此外,增加输入气体混合物中的氧气分压也可以提高PUFAs的产量,Higashiyama等[53]通过增压和增氧控制高山被孢霉发酵中的溶氧浓度,结果表明,最佳DO浓度范围为10~15 mg/L,与在7 mg/L DO下获得的结果相比,该浓度范围内的ARA产率提高了约1.6倍。
4.4 液态发酵模式
丝状真菌的液态发酵模式主要有连续发酵、分批发酵、补料发酵和重复分批发酵等,不同发酵模式为丝状真菌积累脂质提供了优化策略。分批发酵可以获得大量菌体细胞、延长稳定期和提高脂质含量,研究者通过建立发酵参数的数学模型验证分批培养的发酵方式适合深黄被孢霉生产脂质[54]。而补料发酵可以再次激活产脂代谢,Shuib等[55]发现班尼小克银汉霉发酵48 h后的培养基中酶活性的降低以及金属离子的消耗导致其脂质积累停止,经过补料酒石酸铵、葡萄糖及金属离子后,酶活性成功恢复,其脂质含量从32%增加到50%,并增强了GLA的积累。另外,重复分批发酵能够良好的利用营养物质,Dashti等[56]使用不同浓度的氮(1~4 g/L)和葡萄糖(20~40 g/L)对班尼小克银汉霉2A1进行重复分批发酵,结果表明:在20.2 g/L葡萄糖、2.12 g/L酒石酸铵和收获时间48 h的最佳条件下,脂质含量和GLA含量达到最高,分别为6.2 g/L和0.4 g/L,该模式在后续发酵过程中可以重复使用微生物细胞,也是脂质液态发酵中应用最广泛的一种。
5 展望
丝状真菌作为一类生产脂质的微生物,在脂质生产中具有较大的潜力,而且富含的多不饱和脂肪酸对人体健康具有重要意义。为了充分利用这一功能,本文总结了培养基组分对丝状真菌积累脂质的影响,其中改变碳源和氮源的种类或比例对脂质积累产生明显作用,但昂贵的发酵成本减缓了其产业化进程,因此在保持较高的生物量和脂质含量下使用廉价底物替代碳氮源被认为是一种可行的策略。控制液态发酵条件在脂质积累方面也尤为重要,使得丝状真菌不仅拥有适宜的生长环境,还改善了脂质积累量,但通过改变发酵条件不能定向生产多不饱和脂肪酸,因此需要借助诱变或基因工程等手段实现定向生产多不饱和脂肪酸的生产。在未来的发展中,需要加强丝状真菌的基础研究,清晰认识不同丝状真菌脂质合成所涉及的代谢网络以及液态发酵调控机制,特别是,对菌株进行改造并耦合液态发酵过程以强化脂肪酸的生产效率,进一步探究液态发酵与丝状真菌之间的联系,为丝状真菌工业化生产脂质创造更广阔的前景。
