富士和新红星苹果虎皮病抗病性与生理生化特性关系比较研究
2023-12-18王建锋谢欣蔚
王 燕 王建锋 谢欣蔚 王 颖 惠 伟
中国是世界上最大的苹果生产国和消费国,2021 年全国苹果总产量4 579.34 万t,占世界总产量的54.7%。 我国苹果90%以上用于鲜食,贮藏保鲜在缓解果实卖难,实现季产年销中起到了重要作用。 由于贮藏期长, 一些苹果品种在冷藏中、 后期易发生虎皮病, 造成严重的贮藏损失。 虎皮病发病初期, 病变部位呈不规则状淡褐色, 面积较小, 后随冷藏时间延长,病斑扩大, 严重时连接成片。 果实出库后,病斑面积迅速扩大,颜色加深,病果失去商品价值。前人研究表明,虎皮病的发生与品种、采收与处理、贮藏条件等有关。目前有关发病机理的研究主要集中在果实中α-法尼烯及代谢产物和抗氧化能力两个方面。Mditshwa 等发现,果实产生的乙烯诱导α-法尼烯合成,之后它自氧化成共轭三烯(CT)和6-甲基-5-庚烯-2-酮(MHO),诱发虎皮病。 α-法尼烯通过甲羟戊酸途径合成,其中α-法尼烯合成酶(AFS)是α-法尼烯合成过程中关键的限速酶, 而AFS1是AFS基因家族中与苹果虎皮病关系较为密切的基因。此外,苹果虎皮病的发生与果皮中氧化还原状态密切相关。果实衰老过程中形成的活性氧参与α-法尼烯自氧化过程, 降低了膜的完整性, 让处于细胞内不同区域的酚类物质与多酚氧化酶发生酶促反应, 形成醌, 从而产生褐斑, 导致虎皮病。 苹果多酚和类黄酮具有抗氧化作用, 通过清除自由基来保护果皮细胞, 花色素苷是类黄酮的主要组分, 是红色品种着色的物质基础,一般着色面具有良好的抗氧化性, 虎皮病发病率较低。 本文试图通过比较不同虎皮病发生程度的富士(抗病性强)和新红星(感病性强)两个品种在冷藏期间果皮中α-法尼烯代谢以及抗氧化能力等变化, 揭示红色苹果品种虎皮病发生的可能机理, 为培育抗虎皮病苹果品种提供依据。
1 材料与方法
1.1 材料和处理 在陕西白水县采收达到商业成熟度的富士和新红星苹果, 采后当天选取大小均匀、 无机械损伤和病虫害的果实作为试验材料, 次日装箱贮藏于冷库,3 天降温至0±0.5 ℃,相对湿度保持90%~95%。冷藏210 天后转移至25 ℃环境中进行货架贮藏。
1.2 测定指标及方法
1)测定硬度与可溶性固形物。每次取10 个果实,用FT327 型硬度计测硬度,用WYT-J 型手持折光仪测可溶性固形物含量。
2)测定乙烯释放速率与呼吸速率。 参照高俊凤方法, 每个处理从冷库中取10 个果实,在真空干燥器中密封2 小时,用注射器抽取1 mL气体,用安捷伦6890N 气相色谱仪测定乙烯释放速率(单位μL/kg·h),重复3 次。
3)统计虎皮病发病率和病情指数。
4)AFS1基因的表达。 采用改良后的CTAB法, 提取-80 ℃苹果果皮冻样总RNA。 使用TaKaRa 反转录试剂盒对已提纯度较好的RNA进行cDNA 合成。 通过上海生工生物有限公司设计并合成引物,实时定量引物序列为Actin(F:5’-ATCGTGGTCATTGGCCATGT-3’R:5’-AGCCTGTGAGGTTCCAGTAATCAT-3’)AFS1(F:5’-CACAAGAATGAAGATCTTTTGTΑ -3’R:5’ -CAACAACGTTACAAACTGTAAAG-3’)。 将已转录成cDNA 的样品取出后,参照TaKaRa SYBR Premix ExTaq ll 试剂盒说明书进行实时定量PCR 分析。
5)测定α-法尼烯、共轭三烯和MHO。 参照Anet 方法稍作改进,用打孔器打取组织小圆片,随机取20 片,放入50 mL 离心管中,加入10 mL纯化的正己烷,在25 ℃,60 Hz 摇床上震荡提取2 小时,之后过滤定容至10 mL,迅速在232 nm波长下比色,测定α-法尼烯含量。
另取提取液3 mL,于281 nm 及290 nm 处比色,测定共轭三烯含量。
参照Ju 等方法稍作改进,测定MHO 含量。使用固相微萃取—气相色谱法(SPME-GC),用直径2 cm 的打孔器随机打20 个果皮组织圆片(厚度3 mm), 加入15 mL 样品瓶中, 加2 mL NaCl 溶液(200 g/L)密闭3 小时。 用老化后的萃取头萃取20 分钟后进行GC 分析。 进样口解析2 分钟。 GC 设置为不分流进样;柱初温40 ℃保持2 分钟后, 以50 ℃/min 的速率升至250 ℃,运行2 分钟;用氢离子火焰(FID)检测器,温度250 ℃。 用外标法定性,单位μL/kg·h。
6)测定总酚和类黄酮含量。参照陈玮琦等的方法,采用Folin-ciocalteu 法测定总酚。取研磨后的果皮冻样于1%HCL-甲醇溶液中避光浸提24小时,70 ℃超声波提取60 分钟,9 000 rpm/min 离心10 分钟, 取上清液用亚硝酸钠—硝酸铝比色法测定果皮类黄酮含量,每个样品3 次重复。
7)测定花青苷相对含量。 参照宋哲等的方法测定花青苷含量,稍作改进。取10 个样果,将果皮切碎充分混合, 取0.5 g 鲜样, 加入20 mL HCL-甲醇溶液,4 ℃下浸提24 小时, 用分光光度法测定提取液657 nm 和530 nm 处吸光度值, 两者吸光度值之差即为花青苷的相对含量。
8)测定DPPH 自由基清除率。 参照陈玮琦等的方法测定DPPH 自由基清除率,稍作改进。向0.5 mL 样品提取液中加入4.5 mL DPPH 溶液,摇匀,暗处室温放置30 分钟,于517 nm 处测定吸光度值,每个样品重复3 次。
9)数据处理与作图。 所有处理和测定指标均重复3 次, 采用SPSS 20.0 软件进行数据分析,用Excel 2003 软件进行数据统计、作图。
2 结果与分析
2.1 冷藏期间富士和新红星苹果硬度、可溶性固形物含量变化 由图1A 可知,冷藏期间,富士和新红星的硬度皆随冷藏时间延长呈逐渐降低趋势。 除在第120 天时两种苹果的硬度达到相同值6 kg/cm2外, 在整个冷藏过程中富士的硬度整体高于新红星,且差异显著(P<0.05)。由图1B 可知, 富士在整个冷藏期间的可溶性固形物含量始终高于新红星, 两者差异极显著(P<0.01)。冷藏7 个月时,它们的可溶性固形物含量分别由16.8%、12.4%降到14%、10%。
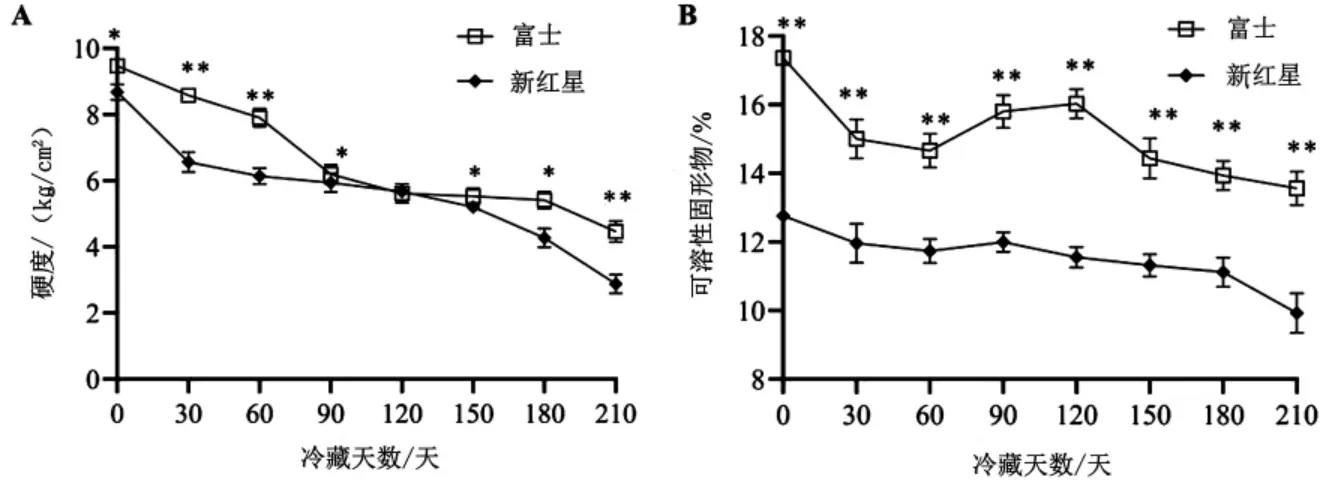
图1 冷藏期间富士和新红星苹果硬度、可溶性固形物含量变化
2.2 冷藏期间富士和新红星苹果乙烯释放速率和呼吸速率的变化 由图2A 可知,整个冷藏期间,富士在第120 天达到乙烯释放峰值,而新红星在冷藏第90 天即达到乙烯释放峰值, 比富士提前30 天,而且峰值极显著高于富士(P<0.01)。由图2B 可知,整个冷藏期间,富士和新红星呼吸速率皆于冷藏第90 天达到峰值,富士呼吸速率始终低于新红星,两者差异显著(P<0.05)。
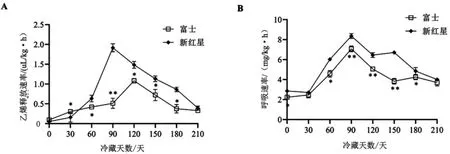
图2 冷藏期间富士和新红星苹果乙烯释放速率和呼吸速率变化
2.3 冷藏期间富士和新红星苹果虎皮病发病率和病情指数的变化 由图3A 可知, 冷藏结束(210 天)时,富士未发生虎皮病,出库室温下(25 ℃)放置10 天后,仍没有发生虎皮病。 冷藏结束(210 天)时,新红星虎皮病果率为73.3%,出库室温下放置10 天后, 新红星病果率增至100%, 相比冷藏期刚结束时, 发病率增加26.7%,差异显著(P<0.05)。 由图3B 可知,冷藏结束(210 天)时新红星虎皮病病情指数为51.1%,出库室温下放置10 天后,病情指数升高到87.8%,相比冷藏期刚结束时增加了36.7%,差异极显著(P<0.01)。

图3 富士和新红星苹果在210 天冷藏和10 天货架期后虎皮病发病率和病情指数指数
2.4 冷藏期间富士和新红星苹果果皮内AFS1基因相对表达量的变化 由图4 可知, 整个冷藏期间, 富士和新红星果皮内AFS1基因的表达量与乙烯的变化趋势相似, 富士比新红星推迟1 个月, 分别在第120 天和90 天达到峰值,且峰值差异显著(P<0.05)
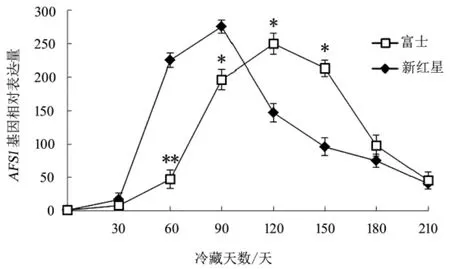
图4 冷藏期间富士和新红星苹果果皮内AFS1 基因相对表达量的变化
2.5 冷藏期间富士和新红星苹果果皮中α-法尼烯、 共轭三烯和MHO 含量的变化 由图5A可知, 整个冷藏期间, 富士和新红星果皮内α-法尼烯含量变化类似AFS1基因表达量变化, 富士于150 天达到峰值, 新红星于120 天达到峰值, 富士峰值低于新红星, 二者差异显著(P<0.05)。由图5B 可知,整个冷藏期间,富士和新红星果皮内共轭三烯含量皆呈增加趋势,富士共轭三烯含量始终低于新红星, 二者差异显著(P<0.05)。 由图5C 可知,冷藏90 天后新红星果皮内MHO 含量呈快速增加趋势, 而富士果皮中MHO 的含量保持相对稳定水平,且冷藏第120 天后均低于新红星, 二者差异显著(P<0.05)。
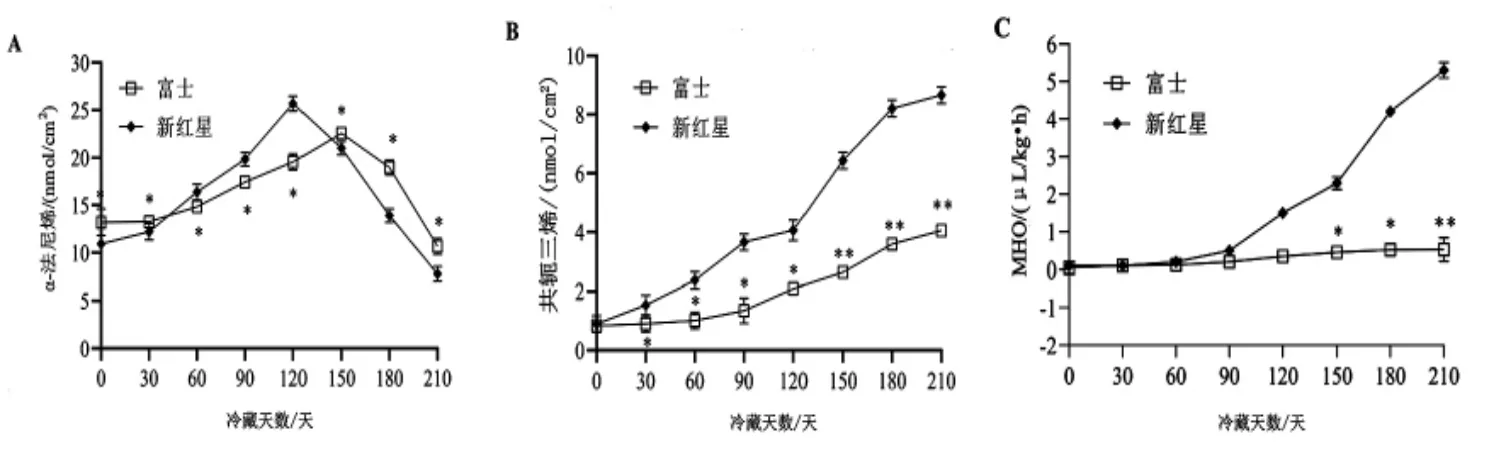
图5 冷藏期间富士和新红星苹果果皮内α-法尼烯、共轭三烯、MHO 含量的变化
2.6 冷藏期间富士和新红星苹果抗氧化能力的变化 由图6A 可知,整个冷藏期间,富士果皮内类黄酮含量波动较大, 在150 天时出现峰值达到0.34 mg/g,而新红星果皮内类黄酮含量较富士变化平稳,二者差异不显著(P>0.05)。
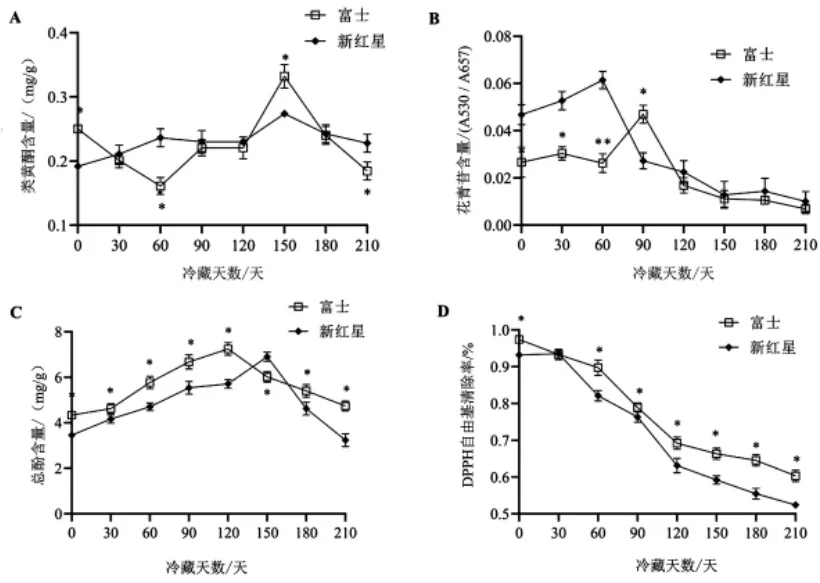
图6 冷藏期间富士和新红星苹果果皮中类黄酮、花青苷、总酚和DPPH 自由基清除率变化
由图6B 可知,整个冷藏期间,两个品种果实花青苷含量都呈现先短暂升高后降低的变化趋势。 在冷藏初期(0~60 天),富士花青苷含量变化不明显,第90 天前有过短暂增加,此后开始降低;果皮内花青苷含量0~60 天时较高,峰值出现在冷藏第60 天, 此后开始降低。 冷藏120 天后, 富士和新红星果皮内花青苷含量差异不显著(P>0.05)。
如图6C 所示,整个冷藏期间,富士和新红星果皮内总酚含量皆呈现先升高后降低的变化趋势,富士于冷藏120 天达到峰值(6.98 mg/g),新红星于150 天达到峰值(6.6 mg/g),整个冷藏期间富士果皮内总酚含量整体高于新红星,二者差异显著(P<0.05)。
自由基清除能力是苹果总抗氧化能力的一个指标,由图6D 可知,整个冷藏期间,两个品种果皮内DPPH 自由基清除能力都呈现逐渐降低趋势,但富士的下降速度相对缓慢。 在冷藏初期(30 天),两个品种DPPH 自由基清除能力差异不显著(P>0.05),30 天后,富士DPPH 自由基清除能力始终高于新红星, 二者差异显著(P<0.05)。由此说明,冷藏期间富士果皮抗氧化能力高于新红星。
3 讨论
虎皮病是一种生理性病害, 不影响果肉颜色,但会造成果实风味下降,失去商品价值。 富士和新红星都是红色苹果品种, 新红星冷藏4个月就开始发生虎皮病, 富士发病期在冷藏7个月以后,程度较轻。
Ding 等研究发现, 青香蕉苹果冷藏90 天虎皮病随乙烯释放速率上升而加重。 本研究发现,冷藏过程中新红星乙烯释放速率、呼吸速率整体高于富士(图2);两个品种冷藏后期虎皮病发病差异极显著(P<0.01),富士始终未发病而新红星发病率极高(图3A),这与品种耐贮性差异有关,新红星果肉容易发绵,而富士则能够一直保持较高的硬度(图1A)。
有关苹果虎皮病发病机理的研究表明,α-法尼烯及代谢产物诱导了虎皮病的发生。 苹果冷藏期间产生大量α-法尼烯, 之后极易被氧化生成共轭三烯, 共轭三烯又氧化成MHO, 这些挥发性物质是诱导虎皮病发生的关键。 索江涛等认为, α-法尼烯和共轭三烯的积累与红富士苹果虎皮病的发生有显著正相关性。 Sabban 等证实了澳洲青苹虎皮病的发生与其内源MHO含量有关。 Farneti 等对澳洲青苹果皮中非靶向代谢物进行研究, 共发现3 种代谢产物, 其中MHO 与虎皮病关系极为密切。 苹果的α-法尼烯合成酶基因AFS1对α-法尼烯合成起着至关重要的作用, 而且AFS1基因受到乙烯调控,感病品种的AFS1基因表达水平更高。 本试验研究发现, 虽然冷藏过程中两个品种果皮的α-法尼烯含量呈现先升高后降低趋势(图5A),但富士AFS1基因表达量和α-法尼烯含量的峰值都比新红星推迟1 个月; 新红星果皮共轭三烯和MHO 含量在冷藏过程中均保持上升趋势, 而富士共轭三烯和MHO 含量一直低于新红星(图5), 这可能与富士苹果较强的抗病性能有关。
虎皮病的发生与果实抗氧化能力有关,富士和新红星都是红色果皮品种, 且果皮中都富含类黄酮等多酚类物质, 它们对虎皮病的抗病性差异是否与这些物质含量有关, 目前未见报道。 伴随α-法尼烯大量氧化, 细胞膜透性增大, 果皮细胞膜系统的完整性及多酚氧化酶(PPO)与底物的区域分布遭到破坏, 最终导致虎皮病发生。 索江涛等研究发现, 1-MCP 处理显著抑制了红富士苹果乙烯形成, 并降低了多酚氧化酶(PPO)活性,进而保持了多酚等抗氧化物质的含量, 最终降低了苹果虎皮病发病率。 苹果富含多酚类物质, 其中黄酮类和花青素类物质是其重要组成, 具有较强的抗氧化、抗脂质过氧化活性, 并能延缓果实衰老褐变。本研究发现, 整体上富士和新红星的类黄酮含量没有显著差异(图6A); 富士前期花青苷比新红星还要低一些(图6B),可能与新红星乙烯和呼吸高峰更早出现有关。 宋哲等人发现, 伴随着苹果果实的衰老, 花青苷迅速降解; 杨小幸等研究发现,DPPH 自由基清除能力减弱可能是酚类物质减少的重要原因。 所以, 本研究中富士DPPH 自由基清除能力高于新红星(图6D)。富士苹果的抗病性与它总的抗氧化能力高有关。
4 结论
新红星苹果的感病性与其乙烯释放速率高有关。 富士对虎皮病的抗性与其低乙烯释放速率和低α-法尼烯及代谢产物含量有关,尤其是共轭三烯、MHO 一直维持低水平, 并且在冷藏过程中保持了较高的抗氧化能力。
针对不同苹果品种的果实虎皮病与乙烯释放速率、抗氧化能力和α-法尼烯及代谢产物的相关性,仍需进一步研究。
