面包乳杆菌ZHG2-1 对荧光假单胞菌群体感应的淬灭作用
2023-12-16吕欣然王扬蕊白凤翎崔方超檀茜倩励建荣郭晓华
吕欣然,温 馨,王扬蕊,白凤翎,崔方超,檀茜倩,励建荣,2*,郭晓华
(1 渤海大学食品科学与工程学院 生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心辽宁省高等学校生鲜食品产业技术研究院 辽宁锦州121013 2 大连工业大学 海洋食品精深加工关键技术省部共建协同创新中心 辽宁大连116034 3 山东美佳集团有限公司 山东日照 276800)
荧光假单胞菌(Psdeuomnoda fluoerncnet)作为水产品中常见的优势腐败菌,其自身分泌的蛋白酶、脂肪酶、嗜铁素、生物膜等因子受N-酰基-高丝氨酸内酯(N-Acyl-homoserine lactones,AHLs)介导的LuxI/R 型QS 系统调控[1-2]。常通过使用抗生素等化学制剂控制腐败微生物对水产品的污染,然而长期使用不仅使细菌产生耐药性,还危害人体健康。以细菌QS 系统为作用靶点,干扰其相关致腐因子的表达,为控制由荧光假单胞菌引发的食品腐败提供新策略。
群体感应淬灭(Quorum quenching,QQ)是以QS 系统为靶点,通过抑制AHLs 合成、干扰AHLs与受体蛋白结合或酶促降解AHLs 等途径淬灭细菌群体感应效应的一种方式[3]。其中,降解AHLs信号分子,使其浓度低于临界值,抑制细菌相关毒力因子表达的酶称之为群体感应淬灭酶(Quorum quenching enzymes,QQE)。其具有专一性强、催化效率高、安全无毒等优点,是目前最为有效的群体感应淬灭途径。依据结构特性,QQE 主要包括AHLs 内酯酶、AHLs 酰基转移酶、AHLs 氧化还原酶[4],根据来源可分为植物源、动物源、微生物源[5-7]。乳酸菌(Lactic acid bacteria,LAB)作为国际公认的安全菌株,已被广泛应用在食品工业领域。研究表明,乳酸菌来源的苯乳酸、细菌素和青霉素酰化酶等代谢产物具有良好的群体感应淬灭活性[8-10],而同种或不同种微生物分泌的淬灭酶结构和催化特性的差异性,导致其淬灭机制也存在一定差异。本课题组前期获得的面包乳杆菌(Lactobacillus crustorum)ZHG2-1 具有QQ 活性,对C4-HSL、C8-HSL 和3-oxo-C12-HSL 等AHLs 信号分子均具有较强的降解作用,该活性物质初步鉴定为AHLs-酰基转移酶或氧化-还原酶[11],然而,其对荧光假单胞菌群体感应的淬灭机制尚不清楚。
鉴于上述问题,本文以面包乳杆菌ZHG2-1为研究对象,利用双层琼脂扩散法分析菌株ZHG2-1 对荧光假单胞菌AHLs 的降解效果。通过测定生物膜、胞外蛋白酶、胞外多糖、生物胺等腐败指标,探究菌株ZHG2-1 对荧光假单胞菌的细胞表型淬灭效应。采用qRT-PCR 技术研究菌株ZHG2-1 对荧光假单胞菌致腐因子基因水平的影响,阐明其淬灭机制,以期为开发水产品新型生物防腐剂提供理论基础。
1 材料与方法
1.1 材料与试剂
1.1.1 菌种及培养条件 面包乳杆菌(Lactobacillus crustorum)ZHG 2-1,分离自辽宁阜新彰武腌制酸黄瓜。荧光假单胞菌(Psdeuomnoda fluoerncnet)PF08,分离自腐败大菱鲆。
紫色杆菌(Chromobacterium violaceum)CV026 为紫色杆菌ATCC 31532 的mini-Tn5 突变体,菌株自身不产生AHLs,仅当外源AHLs 存在时,可产生特征性紫色菌素,检测酰基侧链长度为C4~C8 的信号分子。以上菌种均于本实验室保藏。
1.1.2 培养基与试剂 MRS 肉汤、LB 肉汤、LB 培养基,北京奥博星生物技术有限公司;乙酸乙酯(分析纯级),福晨(天津)化学试剂有限公司;AO/EB 双荧光染色试剂盒,北京雷根生物技术有限公司。
1.2 仪器与设备
Imark 酶标仪,美国BIO-RAD 公司;MS105UD 电子分析天平,瑞士梅特勒-托利多有限公司;SS-4800 场发射扫描电子显微镜,日本日立公司;徕卡激光共聚焦显微镜,德国徕卡仪器有限公司;Bioscerrn C 全自动生长曲线分析仪,上海谓载商贸发展有限公司5804R 高速冷冻离心机,德国Eppendorf 公司;DL-CJ-2N 超级洁净工作台,北京东联哈尔仪器制造有限公司;SPX-25 生化培养箱,宁波海曙赛福实验仪器厂;RE-2000A旋转蒸发器,上海亚荣生化仪器厂;Multifuge1L-R高速冷冻离心机,德国Heraeus 公司;SCIENTZ-10N/A 冷冻干燥机,宁波新芝生物科技股份有限公司;立式自动压力蒸汽灭菌器GI80DS,致微(厦门)仪器有限公司。
1.3 方法
1.3.1 菌株ZHG2-1 上清液与荧光假单胞菌AHLs 的制备
1.3.1.1 菌株ZHG2-1 上清液粗提物的制备 参照Lv 等[12]方法,将面包乳杆菌ZHG2-1 以2%的接种量在MRS 肉汤中活化至第3 代,离心过滤得乳酸菌无细胞上清液(Cell free supernatants,CFS),用乙酸乙酯萃取CFS 活性成分,经真空旋转蒸发、真空冷冻干燥制成粉末状固体,置于-80℃冰箱备用。
1.3.1.2 荧光假单胞菌AHLs 粗提物的制备 将荧光假单胞菌在LB 肉汤中培养,传代2 次后于30 ℃培养16 h,离心过滤后用含0.01%冰醋酸的乙酸乙酯萃取,50 ℃下真空旋转蒸发以消除有机溶剂并用70%甲醇溶解。
1.3.2 菌株ZHG2-1 对荧光假单胞菌AHLs 淬灭活性验证 参照Cui 等[11]方法制备AHLs 检测平板,牛津杯打孔。将菌株ZHG2-1 CFS 调至中性,加入10 μL 荧光假单胞菌AHLs 粗提物,30 ℃培养24 h,离心后取上清,调pH 值至中性,取180 μL 加入打好的孔中,30 ℃培养24 h。
1.3.3 荧光假单胞菌与紫色杆菌CV026 的最小抑菌浓度与亚抑制质量浓度 将菌悬液浓度为106CFU/mL 的荧光假单胞菌与紫色杆菌CV026(100 μL)分别加入96 孔板中,依次加入10 μL 上清液粗提物至终质量浓度分别为0,2.0,4.0,8.0,16.0,32.0 mg/mL,以不添加提取物的LB 肉汤作为阴性对照,30 ℃静置培养24 h。在波长595 nm 处每隔2 h 测OD 值。
1.3.4 菌株ZHG2-1 上清液粗提物对荧光假单胞菌产生物膜量的影响
1.3.4.1 生物膜抑制的定量分析 取200 μL 荧光假单胞菌悬液到96 孔板中,添加菌株ZHG2-1粗提物至终质量浓度为1.0,2.0,3.0 mg/mL,以不添加提取物的LB 肉汤作为阴性对照,30 ℃培养12 h,吸去多余悬浮液,用无菌PBS 洗涤,参照Shikha 等[9]方法结晶紫染色定量检测生物模量。同时,参考Cui 等[11]方法测定浮游细胞存活率。
1.3.4.2 生物膜清除的定量分析 在96 孔板中加入荧光假单胞菌培养物(106CFU/mL),30 ℃培养12 h,移除悬浮液,PBS 洗涤后,向孔板中添加不同亚抑制质量浓度的菌株ZHG2-1 粗提物,以不添加粗提物的LB 肉汤作为阴性对照,30 ℃培养24 h,按上述方法定量检测生物膜残留量和浮游细胞存活率。
1.3.5 菌株ZHG2-1 上清液粗提物对荧光假单胞菌生物膜结构的影响 在24 孔板中放入无菌硅片(1 cm×1 cm),取1 mL 荧光假单胞菌菌液浸没,样品处理如1.3.4 节方法。PBS 洗涤硅片,2.5%戊二醛固定12 h,无菌水洗涤3 次,40%,70%,90%,100%乙醇依次脱水处理15 min,待硅片干燥后喷金,扫描电镜(SEM)观察生物膜结构。
1.3.6 菌株ZHG2-1 上清液粗提物对荧光假单胞菌致腐因子的影响
1.3.6.1 胞外多糖 参照Mohammed 等[13]方法,将荧光假单胞菌与不同亚抑制质量浓度的菌株ZHG2-1 粗提物30 ℃共培养16 h,离心,将沉淀重悬于0.85% NaCl 溶液中,离心30 min,将上清液与冷乙醇按体积比1∶3 混合,4 ℃过夜沉淀,移除上清液后,苯酚硫酸法定量检测胞外多糖含量。
1.3.6.2 胞外蛋白酶 参照Vijayaraghavan 等[14]方法制备脱脂牛奶平板,牛津杯打孔。将不同质量浓度的菌株ZHG2-1 粗提物与荧光假单胞菌30 ℃共培养24 h,离心过滤后取180 μL 上清液加入打好的孔中,30 ℃培养24 h,测量孔径周围透明圈的大小,计算抑制率。
1.3.6.3 生物胺 根据张黎明等[15]的方法制备生物胺检测培养基,牛津杯打孔。取180 μL 菌株ZHG2-1 粗提物与荧光假单胞菌共培养物上清液加入孔中,30 ℃培养24 h,测量孔径周围蓝紫色圈的大小,计算抑制率。
1.3.6.4 脂肪酶 根据Leelatulasi 等[16]方法并稍作修改。50 mL 沸水将3 mL 三丁甘油酸酯和1 g聚乙烯醇-2000 混合乳化,调pH 值至中性,取4 mL 乳化液与100 mL LB 琼脂混合,制备脂肪酶琼脂平板。取180 μL 共培养物上清液加入孔中,30℃培养24 h,测量孔径周围透明圈的大小,计算抑制率。
1.3.7 菌株ZHG2-1 上清液粗提物对荧光假单胞菌QS、生物膜及致腐基因表达的影响 将过夜活化的荧光假单胞菌菌液稀释至106CFU/mL,加入3.0 mg/mL 菌株ZHG2-1 粗提物,30 ℃培养24 h,离心,弃上清,无菌PBS(pH 7.2)洗涤沉淀,再次离心收集菌体。将菌体送至北京华大蛋白质研发中心进行qRT-PCR 测试。qRT-PCR 用Gene 9600 定量PCR 仪进行,引物序列见表1,荧光假单胞菌的16S rRNA 为内参基因,引物由上海Sangon Biotech 公司设计并合成。
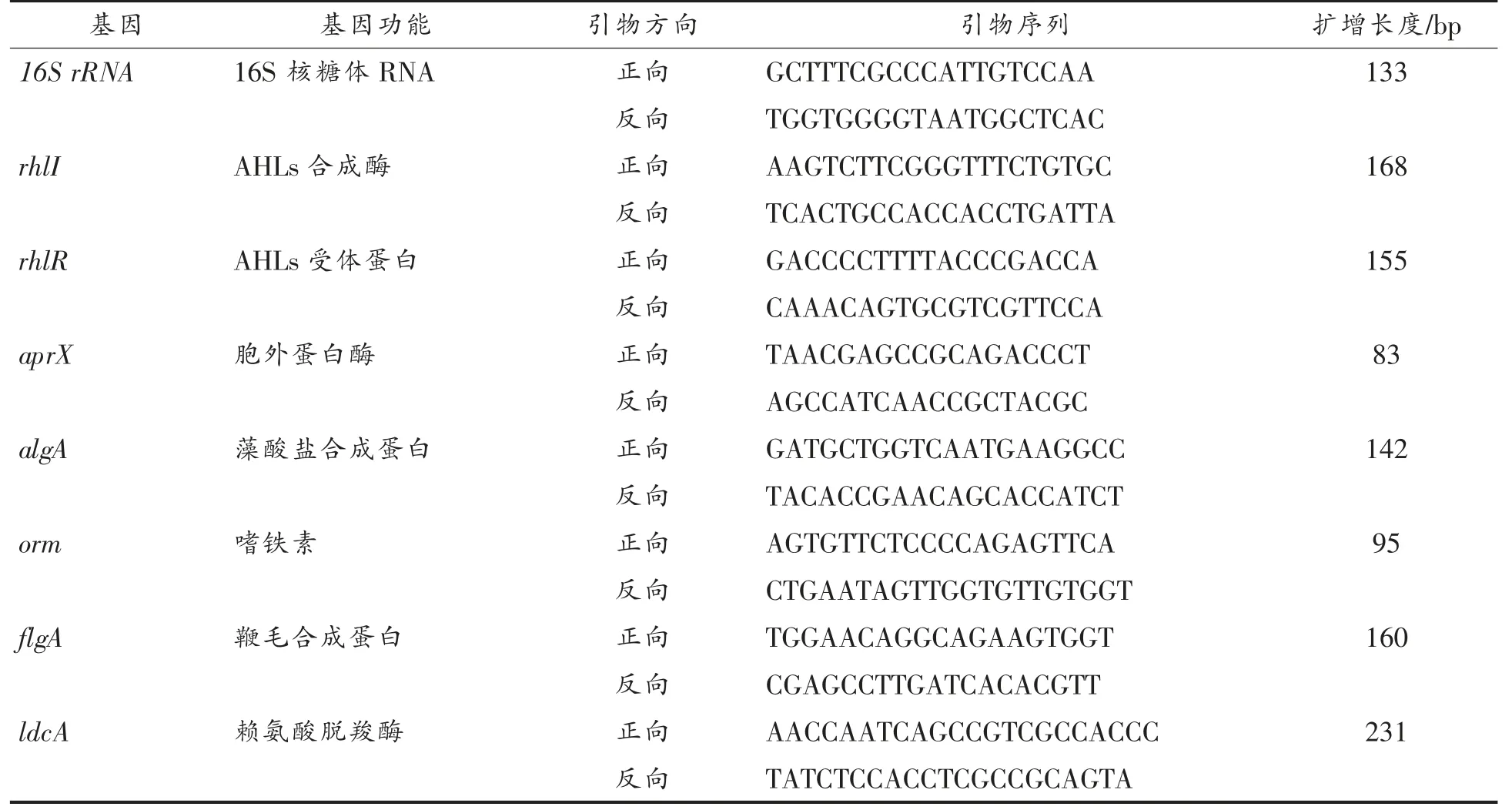
表1 荧光假单胞菌qRT-PCR 引物序列Table 1 The qRT-PCR primer sequences of Pseudomonas fluorescens
1.4 数据处理与分析
每组试验设3 个平行,用SPSS 19.0 软件进行数据的分析处理,Origin 8.5 软件进行绘图处理。
2 结果与分析
2.1 菌株ZHG2-1 对荧光假单胞菌AHLs 的降解活性
采用双层琼脂扩散法检测面包乳杆菌ZHG2-1 CFS 对荧光假单胞菌AHLs 的降解活性。由图1 可知,菌株ZHG2-1 CFS 对荧光假单胞菌AHLs 的降解活性为100%。

图1 菌株ZHG2-1 CFS 对荧光假单胞菌AHLs淬灭活性Fig.1 The quenching activity of strain ZHG2-1 CFS on Pseudomonas fluorescens AHLs
2.2 荧光假单胞菌和紫色杆菌CV026 的最小抑菌浓度与亚抑制质量浓度
面包乳杆菌ZHG2-1 上清液粗提物对荧光假单胞菌和紫色杆菌的最小抑菌浓度分别为8.0 mg/mL 和4.0 mg/mL。为确认菌株ZHG2-1 粗提物的降解活性是出自群体感应淬灭作用,还是抑菌作用,研究不同亚抑制质量浓度对荧光假单胞菌和紫色杆菌生长的影响。由图2 可知,1.0,2.0,3.0 mg/mL 的菌株ZHG2-1 粗提物对荧光假单胞菌和紫色杆菌的生长均无影响,选取这3 个质量浓度作后续研究。
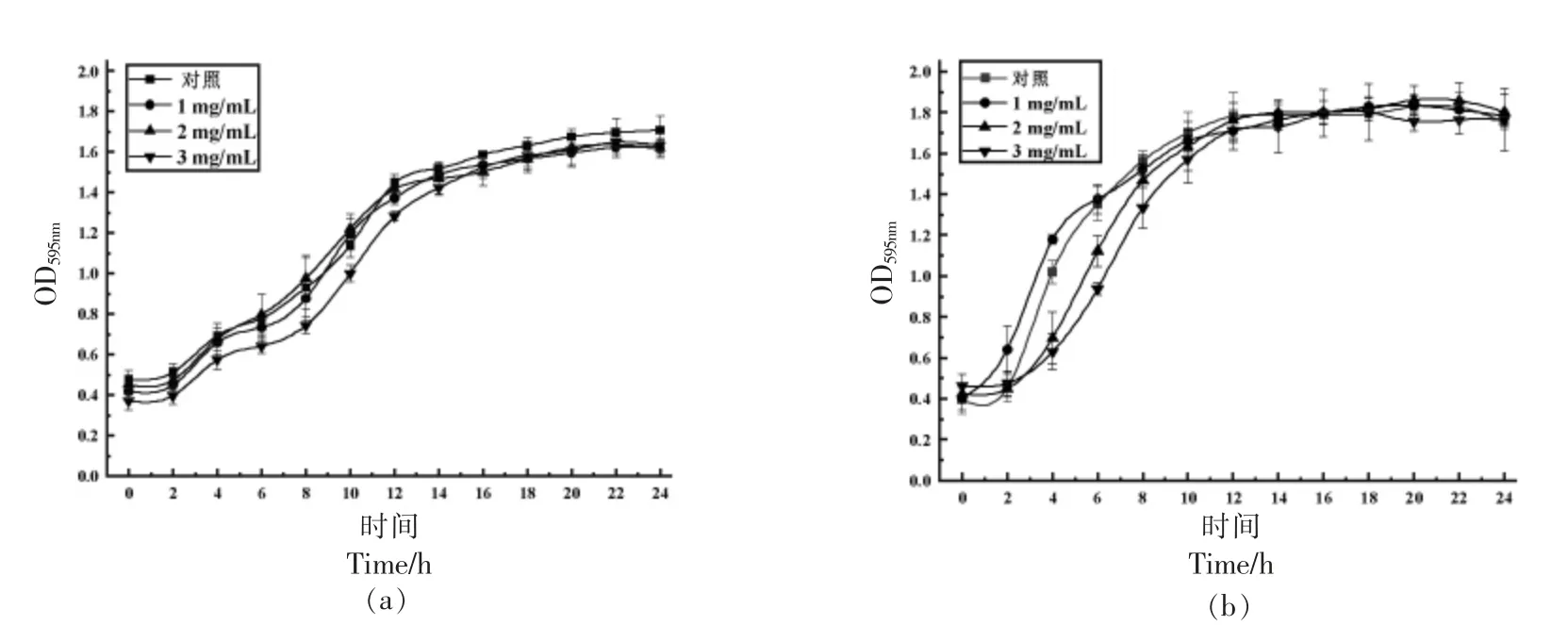
图2 亚抑制质量浓度下菌株ZHG2-1 粗提物对荧光假单胞菌(a)和紫色杆菌(b)生长的影响Fig.2 Effects of strain ZHG2-1 crude extract on the growth of Pseudomonas fluorescens(a)and Chromobacterium violaceum(b)at sub-inhibitory mass concentrations
2.3 面包乳杆菌ZHG2-1 对荧光假单胞菌产生物膜量的影响
生物膜指黏附在物体表面的细菌与其分泌的胞外基质聚集形成的群落组织,其对外界环境变化的抵抗力远高于浮游细胞[17]。在食品加工中,细菌能快速附着在设备及食品表面形成生物膜,并难以清除,给食品工业带来严峻考验[18]。图3 所示,在不影响荧光假单胞菌生长的前提下,质量浓度为1.0,2.0,3.0 mg/mL 的菌株ZHG2-1 粗提物对荧光假单胞菌生物膜抑制率分别为(9.66±2.92)%,(20.46±3.72)%,(31.32±3.39)%,清除率分别为(28.62±3.20)%,(37.07±3.82)%,(62.82±5.84)%,且呈质量浓度依赖性。Shikha 等[9]研究发现鼠李糖乳杆菌(Lactobacillus rhamnosus)上清液对铜绿假单胞菌生物膜形成的抑制率为35.01%,对成熟生物膜的清除率约为75%,本研究结果与其相近。上述结果说明菌株ZHG2-1 粗提物通过群体感应有效抑制和清除荧光假单胞菌生物膜。
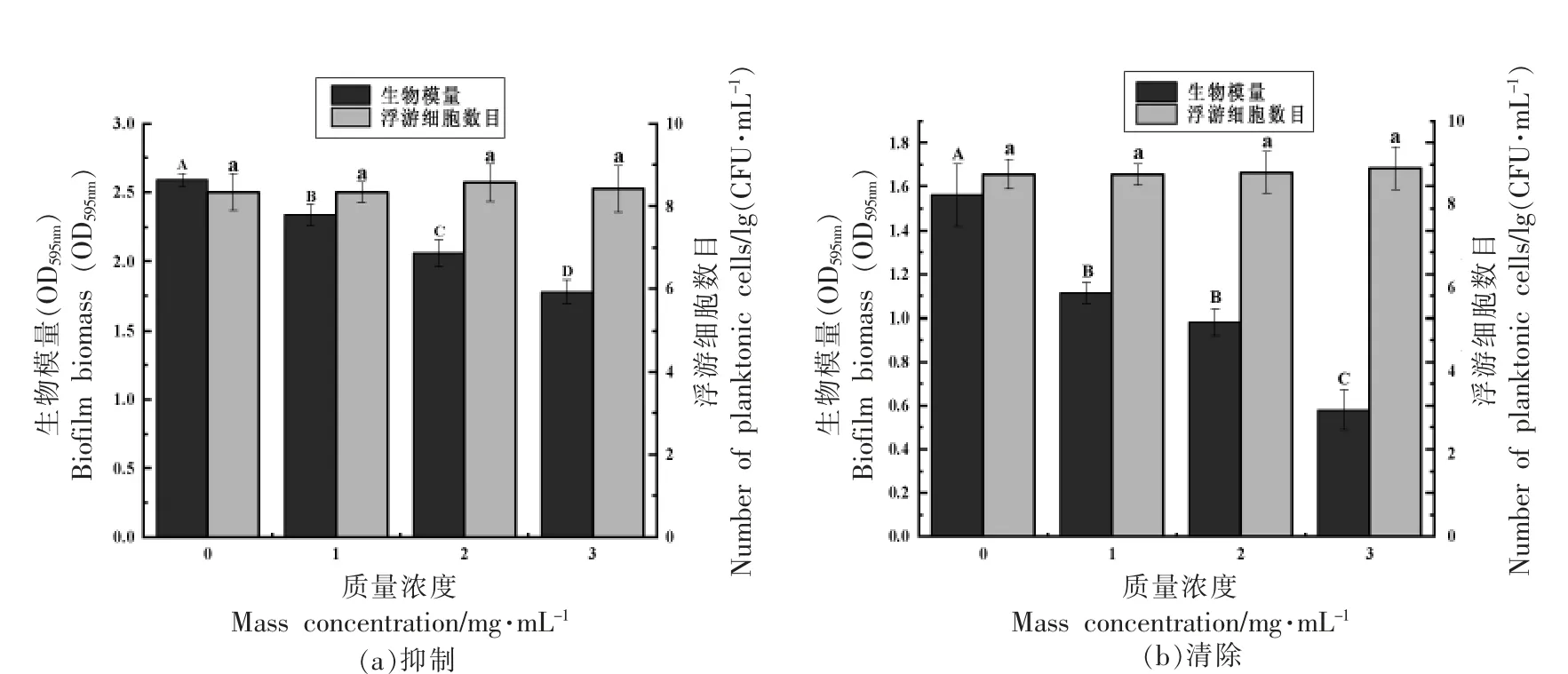
图3 菌株ZHG2-1 粗提物对荧光假单胞菌生物膜量及浮游细胞数目的影响Fig.3 The effects of strain ZHG2-1 crude extract on the Pseudomonas fluorescens biofilm biomass and number of planktonic cells
2.4 面包乳杆菌ZHG2-1 对荧光假单胞菌生物膜结构的影响
利用扫描电镜观察亚抑制质量浓度菌株ZHG2-1 粗提物对荧光假单胞菌生物膜二维结构的影响,如图4 所示,未经处理的荧光假单胞菌生物膜厚实紧密,随着菌株ZHG2-1 粗提物质量浓度的不断增大,致密的膜结构逐渐被解体。当用3.0 mg/mL 的菌株ZHG2-1 粗提物处理时,细菌胞外聚合物显著减少,细菌菌落处于分散状态。结果表明,菌株ZHG2-1 粗提物可通过抑制荧光假单胞菌胞外基质的形成,减弱细菌生物膜形成能力。图4b 显示经菌株ZHG2-1 粗提取处理、预先形成生物膜后,生物膜结构疏松分散,抑制效果没有清除效果显著,表明菌株ZHG2-1 粗提物可破坏荧光假单胞菌成熟生物膜结构。Lv 等[12]研究发现植物乳杆菌CY 1-1 粗提物处理的温和气单胞菌(Aeromonas sobria)数量减少且黏附性差,3.0 mg/mL 粗提物处理几乎没有形成生物膜。1.0,2.0 mg/mL 和3.0 mg/mL CY1-1 粗提物处理后的成熟生物膜结构稀疏,形成量显著低于对照组,本研究结果与其一致。

图4 菌株ZHG2-1 粗提物对荧光假单胞菌生物膜结构的影响Fig.4 Effects of strain ZHG2-1 crude extract on Pseudomonas fluorescens biofilm structure
2.5 面包乳杆菌ZHG2-1 对荧光假单胞菌产胞外多糖的影响
胞外多糖是组成细菌生物膜的重要基质,主要由藻酸盐、生物被膜胞外基质多聚糖以及在菌膜和生物被膜的形成过程中起作用的胞外多糖(Pel)等构成[19]。与如图5a 所示,菌株ZHG2-1 粗提物可显著降低荧光假单胞菌菌胞外多糖分泌,并显示质量浓度依赖性。与对照组相比,当用3.0 mg/mL 菌株ZHG2-1 粗提物处理细菌时,抑制率达(64.08±4.47)%。Yeon 等[20]研究发现酿酒酵母无细胞上清液对单增李斯特菌胞外多糖的抑制率为65.1%~86.02%,本研究结果与其相似。
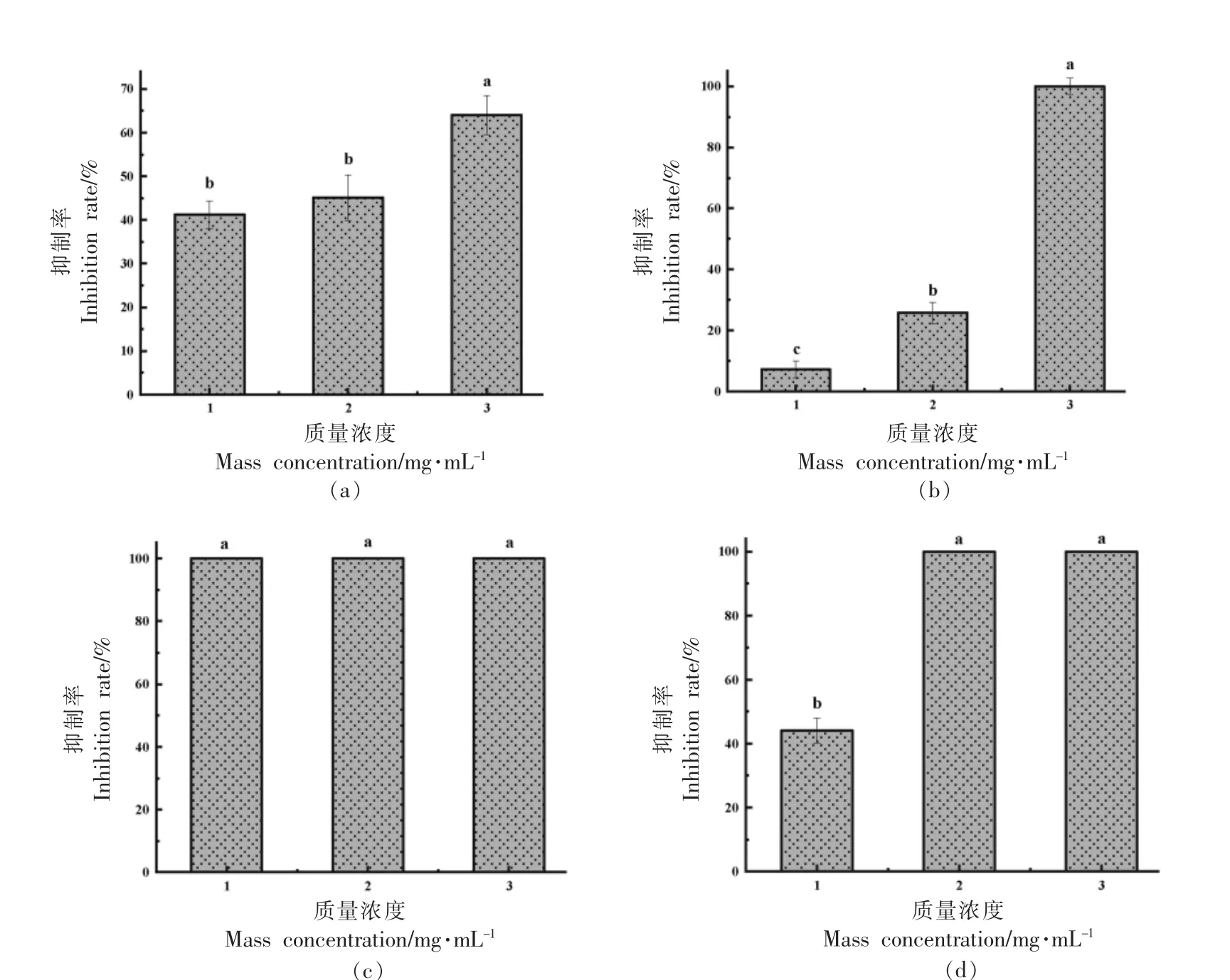
图5 菌株ZHG2-1 粗提物对荧光假单胞菌产胞外多糖(a)、胞外蛋白酶(b)、生物胺(c)、脂肪酶(d)影响Fig.5 Effects of strain ZHG2-1 crude extract on Pseudomonas fluorescens production of extracellular polysaccharides(a),extracellular proteases(b),biogenic amines(c),and lipases(d)
2.6 面包乳杆菌ZHG2-1 对荧光假单胞菌产胞外蛋白酶的影响
荧光假单胞菌可分泌胞外蛋白酶,能将水产品中富含的蛋白质、脂质、游离氨基酸等成分降解,产生致腐代谢物,使水产品丧失原有风味和品质[21]。由图5b 可知,随着菌株ZHG2-1 粗提物质量浓度的增大,荧光假单胞菌分解蛋白能力不断被减弱,3.0 mg/mL 时抑制率达100%,结果表明,菌株ZHG2-1 上清液提取物能显著抑制荧光假单胞菌的胞外蛋白酶的产生。Chu 等[22]研究发现芽孢杆菌上清液对嗜水气单胞菌YJ-1 蛋白酶抑制率为83.9%,本研究结果与其相似。
2.7 面包乳杆菌ZHG2-1 对荧光假单胞菌产生物胺影响
生物胺是一类低分子碱性含氮化合物,主要由微生物产生的氨基酸脱羧酶分解游离氨基酸形成。水产品腐败会产生大量生物胺,过量积累会对人体造成毒害作用[23]。生物胺平板的原理是细菌可分解培养基中游离氨基酸,溴甲酚紫可与产生的碱性生物胺呈蓝紫色反应,蓝紫色圈直径越大,细菌产生物胺能力越强。如图5c 所示,对照组中,荧光假单胞菌有很强的产胺能力,而1.0 mg/mL 菌株ZHG2-1 粗提物可完全抑制生物胺的产生,结果表明,较低质量浓度的菌株ZHG2-1 粗提物可显著抑制荧光假单胞菌生物胺的产生。冯杰等[24]研究发现2.0 mg/mL 蓝莓花色苷对波罗的海希瓦氏菌(Shewanella balticia)腐胺含量的抑制率为94.4%,本研究结果与其相似。
2.8 面包乳杆菌ZHG2-1 对荧光假单胞菌产生脂肪酶的影响
脂肪酶被定义为甘油三酯酰基水解酶。假单胞菌属通过分泌脂肪酶催化甘油三酯分解为脂肪酸、甘油等物质,会改变肉制品肌肉组织,从而使肉制品变黏、变色、产生异味,导致腐败发生[25-26]。本文采用三丁酸甘油酯平板透明圈法研究不同质量浓度菌株ZHG2-1 粗提物对荧光假单胞菌脂肪酶活性的影响。如图5d 所示,1.0 mg/mL 菌株ZHG2-1 粗提物抑制率为(44.09±3.95)%,随着质量浓度的增大,抑制活性可达100%。Shen 等[27]研究发现来源于荧光假单胞菌的AHLs-酰基转移酶PF-1240 可干扰蜂房哈夫尼亚菌脂肪酶的形成,本研究结果与其相似。
2.9 面包乳杆菌ZHG2-1 对荧光假单胞菌QS、生物膜及致腐基因表达的影响
为了探究ZHG2-1 的群体感应淬灭机制,通过qRT-PCR 检测ZHG2-1 对荧光假单胞菌QS、生物膜及致腐基因的表达水平。在荧光假单胞菌QS 系统中,rhlI 编码的是AHLs 合成酶,rhlR 编码的是AHLs 受体蛋白,两者组成的rhlI/R 系统共同调控荧光假单胞菌QS 现象[28]。aprX 是荧光假单胞菌胞外蛋白酶基因[29]。algA 编码的藻酸盐合成蛋白是构成胞外多糖的重要组分,不仅利于细菌的黏附和定殖,在生物膜成熟后期还可以帮助细菌摄取和利用环境中的微量元素[30]。orm 是嗜铁素合成基因,有助于细菌络合食品基质中的Fe3+[31]。flgA编码与生物膜形成以及细菌运动能力相关的鞭毛合成蛋白[32]。ldcA 编码赖氨酸脱羧酶,细菌通过酶解食品中的游离氨基酸,进而形成生物胺[33]。
由图6 可知,3.0 mg/mL ZHG2-1 处理下,rhlI和rhlR 的基因表达被显著下调0.93 和0.99,与荧光假单胞菌生物膜及腐败特性相关的aprX、algA、orm、flgA、ldcA 等基因的表达水平也被显著抑制,分别下调0.91,0.79,0.90,0.94,0.93,这与前期腐败表型的结果一致。Toushik 等[34]研究发现植物乳杆菌J.27 和M.21 可以下调致病菌生物膜形成及QS 相关基因表达,qRT-PCR 结果与其相似。乳酸菌分泌的群体感应猝灭活性物质可以下调QS(luxS、lasB、rhlR、pfs)相关基因的表达,从而使食源性致病菌致病性下降[35]。由此推测面包乳杆菌ZHG2-1 可通过干扰QS 基因表达,进而影响荧光假单胞菌生物膜和腐败因子的产生。

图6 3.0 mg/mL 菌株ZHG2-1 粗提物对荧光假单胞菌QS、生物膜及致腐基因表达的影响Fig.6 Effects of 3.0 mg/mL strain ZHG2-1 crude extract on the expression of QS,biofilm and putrefaction genes of Pseudomonas fluorescens
3 结论
面包乳杆菌ZHG2-1 具有较强的群体感应淬灭活性,其乙酸乙酯提取物可显著抑制荧光假单胞菌生物膜及相关致腐因子的产生。菌株ZHG2-1 粗提物对荧光假单胞菌生物膜形成量及预先形成的成熟生物模量均有显著的抑制作用。扫描电镜进一步显示菌株ZHG2-1 粗提物处理使其生物膜量显著减少,细菌菌落处于分散状态。在亚抑制质量浓度的菌株ZHG2-1 粗提物处理下,荧光假单胞菌胞外多糖、生物胺、蛋白酶、脂肪酶等致腐因子均被显著抑制,与细菌QS、生物膜、腐败因子相关的基因表达水平也被显著下调。综上所述,面包乳杆菌ZHG2-1 作为天然安全的生物防腐剂,通过干扰细菌QS 系统,调控生物膜和致腐基因表达,进而影响生物膜及相关腐败因子的产生,其具体机制可通过转录组学进一步研究。
