水竹秆形特征对竹林经营措施的可塑性响应
2023-12-13冯博杰潘永柱林康民朱强根
冯博杰,潘永柱,林康民,朱强根
(1. 浙江省林业技术推广总站,浙江 杭州,310014;2. 丽水市林业技术推广总站,浙江 丽水 323000;3. 丽水学院 生态学院,浙江 丽水 323000 )
水竹Phyllostachysheteroclada是禾本科Gramineae 刚竹属Phyllostachys散生竹种,为长江流域及以南地区最常见的野生竹种之一,也是黄河和长江流域广为栽培的一个竹种[1]。水竹适应性强,病虫害少,不仅笋味鲜美、营养价值高,而且人工栽培种可长成胸径达12 cm、高达9 m 以上的大径材,是重要的乡土优良笋、竹两用竹种[2]。目前,对于水竹的研究仅限于生物学特性[3]、化学成分[4]及栽培育苗[5]等方面,而就其立竹构件因子及对抚育经营或干扰措施(如施肥、遮阴)的响应未见有研究报道。因此,采用异速生长分析方法研究处理效应对不同功能构件之间的资源分配变化,有助于揭示竹类植物性状与环境的响应关系[6]。鉴于此,本文就以下几个方面开展研究:(1)从水竹地上构件因子角度出发,其地径与株高、枝下高、枝下节数关系是否为异速生长关系;(2)在不同营林条件下(施肥与不施肥、遮阴与不遮阴、灌溉与不灌溉),其构件因子的异速生长关系是否会发生变化。通过对上述问题的探讨,旨在揭示水竹秆形特征关系及其对经营条件和环境因素等的响应,有着重要的生产和生态学意义。
1 材料与方法
1.1 研究区概况
研究区位于浙江省松阳县望松街道翁村,地理位置为28°19' ~ 28°36' N,119°30' ~ 119° 45' E,属于典型的亚热带季风气候区,四季分明,雨量丰沛,年平均降水量为1 500 mm,空气湿润,年平均气温约为17 ℃,极端最高气温为40 ℃,极端最低气温为-9.7 ℃,无霜期约为236 d。区内地貌以山地丘陵为主,中部为地势较低的台地,四周为低山丘陵,土壤类型为红壤和黄壤两大土类,海拔在100 ~ 1 100 m,平均海拔为375 m,植被覆盖密集[7]。
1.2 样地的设置与测定方法
水竹地径(diameter at ground level)测量点为从水竹基部向上20 cm 处,用电子游标卡尺读数;枝下高(under branch height,UBH)测定是从水竹出土的基部到现存枝条基部的长度;枝下节数(number of nodes under branches)是指从水竹出土的基部到现存枝条基部的节数。
2020 年7 月,在翁村不同海拔和不同坡度的水竹林样地设置遮阴、施肥、灌溉三个因素,遮阴采用遮阳网搭建,测得正午光照强度为全光照的(30±4.2)%;施肥使用螯合型笋竹专用肥(氮∶磷∶钾= 17∶8∶5),用量为750 kg·hm-2,沟施;灌溉为0 ~ 20 cm 土层的土壤相对含水量低于55%时及时灌溉;3 个因素分别设置对照,为不遮阴、不施肥、不灌溉。由于样地选取受实际条件所限,试验采用了不完全设计,形成6 个组合处理(见表1)。
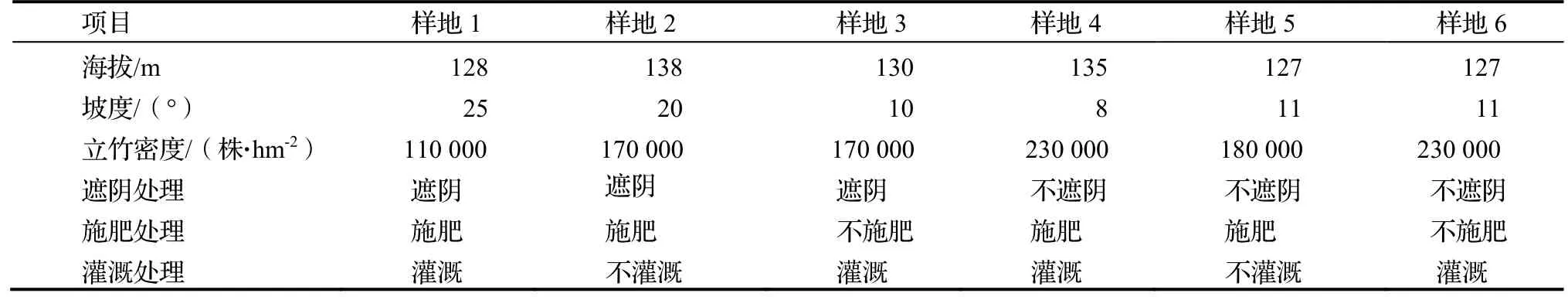
表1 水竹样地设置基本情况Tab. 1 Treatments and controls of sample plots of Ph. heteroclada
6 个处理分别安排在6 块20 m × 30 m 的野生多年生水竹纯林试验样地上,除试验所设计因素外,其他抚育措施均保持一致。2022 年3 月,在每块样地内划设5 m×8 m 的样方5 个,每个样方随机选取5 株标准水竹伐倒,测定每株地径、株高、枝下高、枝下节数。
1.3 数据处理
数据的图表制作采用Excel 2010 完成,地径与株高、枝下高、枝下节数的异速生长分析采用标准主轴(standardized major axis,SMA)估计法[8]。
2 结果与分析
2.1 水竹地径与株高、枝下高、枝下节数的异速生长关系
水竹地径与株高、枝下高、枝下节数的关系采用SMA 估计法,在不考虑遮阴、施肥和水分处理下,水竹地径与株高、枝下高、枝下节数的关系均呈显著的正相关关系(R2= 0.433 1,P< 0.001,R2= 1.559 7,P< 0.001;R2= 0.433 1,P< 0.001),异速生长斜率分别为0.710 4(95%CI= 0.628 ~ 0.803 6)、1.559 7(95%CI= 1.3443~ 1.809 6)、0.896 5(95%CI= 0.782 1 ~ 1.027 7),地径与株高、地径与枝下节数的SMA 斜率均显著小于1(P< 0.05),地径与枝下高的SMA 斜率极显著大于1(P< 0.001)。以上结果表明,随着水竹地径的增大,株高和枝下节数的相对增长速度下降,枝下高的相对增长速度上升。
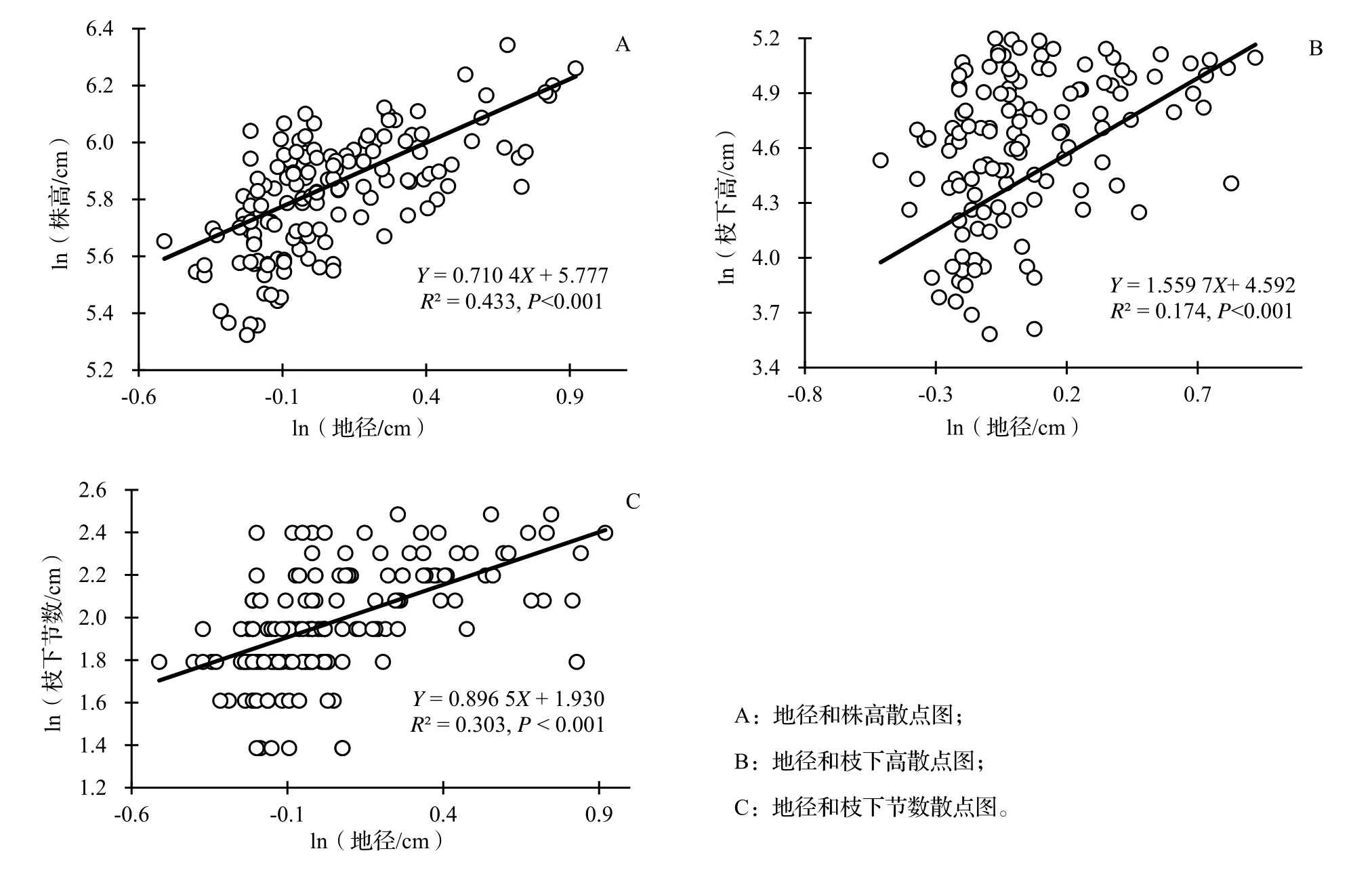
图1 水竹地径与株高、枝下高、枝下节数的异速生长关系Fig. 1 Allometric relationship of ground diameter with height, clear bole height and clear bole nodes of Ph. heteroclada
2.2 遮阴和施肥对水竹秆形特征异速生长关系的影响
2.2.1 不同遮阴处理下施肥对水竹地径、株高异速生长关系的影响 在不遮阴处理中,施肥与不施肥处理间水竹地径、株高的SMA 斜率之间差异不显著(P= 0.086,图2A),表明施肥没有改变水竹地径和株高的异速生长关系,共同斜率为1.007 1(95%CI= 0.761 2 ~ 1.336 0,P< 0.001),与1.0 没有显著差异(P> 0.05),二者呈等速生长关系。与不施肥处理相比,施肥处理水竹的SMA 截距沿着Y 轴出现了显著的正向移动(移动量0.185,Wald 检验,P< 0.001),表明在同等地径下,施肥处理水竹的株高大于不施肥处理。在沿共同斜率方向上,施肥处理和不施肥处理的SMA 主轴移动量也达到显著水平(P< 0.05),表明整体上施肥处理水竹的地径与株高比不施肥处理的更大。
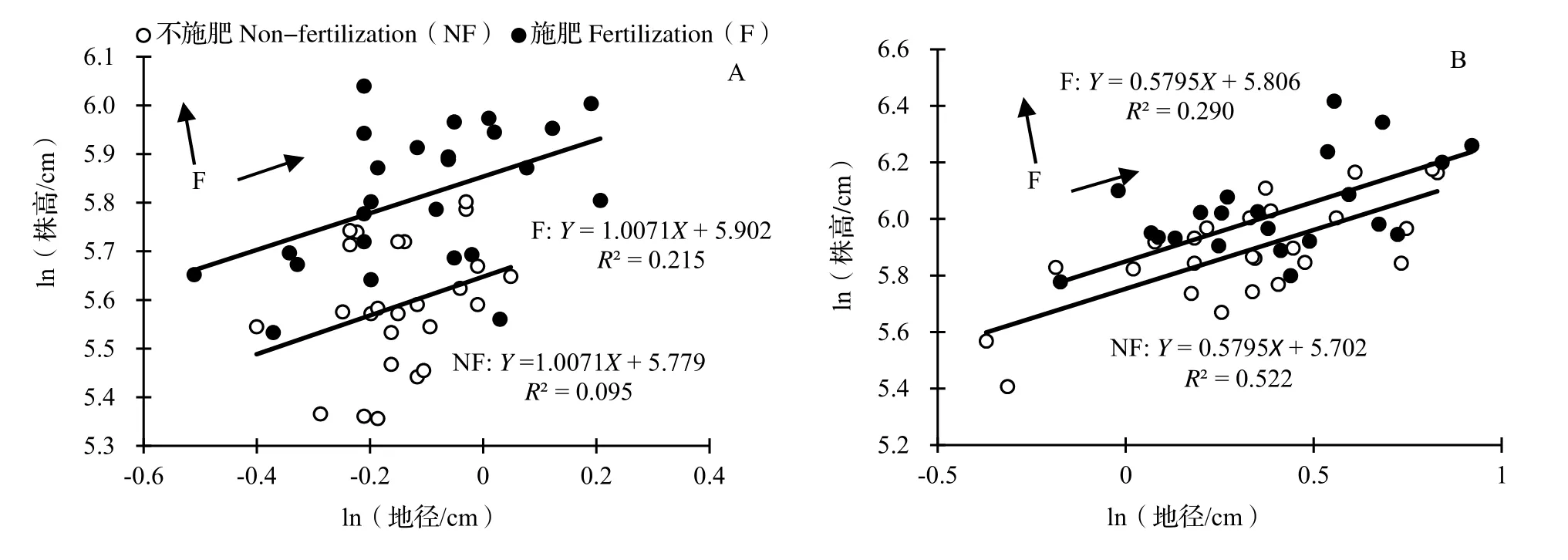
图2 不遮阴(A)和遮阴(B)处理下不同施肥水平水竹地径与株高的异速生长关系Fig. 2 Allometric relation of ground diameter and height of Ph. heteroclada with different fertilization under no shading (A) and shading (B)
在遮阴处理下,施肥与不施肥处理间水竹地径、株高异速生长关系的斜率之间差异不显著(P= 0.956;图2B),共同斜率为0.579 5(95%CI= 0.460 2 ~ 0.729 8,P< 0.001),显著小于1.0(P< 0.001),表明施肥没有改变水竹地径和株高的异速生长关系。在共同斜率下进一步分析得到,施肥下SMA 主轴在Y 轴方向出现了正向的显著移动(P< 0.05),移动量是0.107,即1.113 cm,表明在同等地径下,施肥处理的株高大于不施肥处理的株高。在沿共同斜率方向上,施肥和不施肥处理的SMA 主轴移动量也达到显著水平(P< 0.05),表明整体上施肥处理水竹的地径与株高比不施肥处理的地径与株高更大。
比较图2A 与图2B 可以看出,不管是施肥或不施肥处理,不遮阴和遮阴水竹的地径、株高标准主轴SMA 斜率之间存在显著差异(SMA 分析,P= 0.001),前者与1.0 无显著差异(P> 0.05),后者显著小于1.0(P< 0.05)。
2.2.2 不同遮阴处理下施肥对水竹地径、枝下高异速生长关系的影响 在不遮阴处理下,施肥和不施肥处理下水竹的地径与枝下高异速生长指数存在显著差异(P= 0.013,图3A),施肥处理的斜率1.525(95%CI= 0.931~ 2.498)小于不施肥处理的斜率3.471(95%CI= 2.335 ~ 5.159),且两者均显著大于1.0(P< 0.001)。

图3 不遮阴(A)、遮阴(B)处理下不同施肥水平水竹地径与枝下高的异速生长关系Fig. 3 Allometric relation of ground diameter with clear bole height of Ph. heteroclada with different fertilization under no shading (A) and shading (B)
在遮阴处理下,施肥与不施肥处理水竹地径、枝下高异速生长关系的斜率之间差异不显著(P= 0.479,图3B),共同斜率为0.881 4(95%CI= 0.663 6 ~ 1.170 5,P< 0.001),显著小于1.0(P< 0.05),表明施肥没有改变地径和枝下高的异速生长关系。在共同斜率下进一步分析得到,施肥下SMA 主轴在Y 轴方向出现了正向的极显著移动(P< 0.001),移动量是0.322,即1.379 cm,表明在同等地径下,施肥处理水竹的枝下高大于不施肥处理。在沿共同斜率方向上,施肥和不施肥的SMA 主轴移动量也达到极显著水平(P< 0.001),表明整体上施肥的水竹地径与枝下高比不施肥的水竹地径与枝下高更大。
比较图3A 与图3B 可以看出,不管是施肥或不施肥处理,不遮阴和遮阴水竹的地径、枝下高标准主轴SMA斜率之间存在显著差异(SMA 分析,P= 0.001),前者显著大于1.0(P< 0.001), 后者显著小于1.0 (P< 0.05)。
2.2.3 不同遮阴处理下施肥对水竹地径、枝下节数异速生长关系的影响 在遮阴处理下,由不同施肥处理下水竹地径与枝下节数的异速生长分析可见,施肥和不施肥处理下的地径与枝下节数异速生长指数存在极显著差异(P< 0.001,图4A),施肥处理的斜率0.755 4(95%CI= 0.514 9 ~ 1.108 2)小于不施肥处理的斜率1.755 8(95%CI= 1.189 0 ~ 2.592 7),且前者显著小于1.0(P< 0.001),后者显著大于1.0(P< 0.001)。
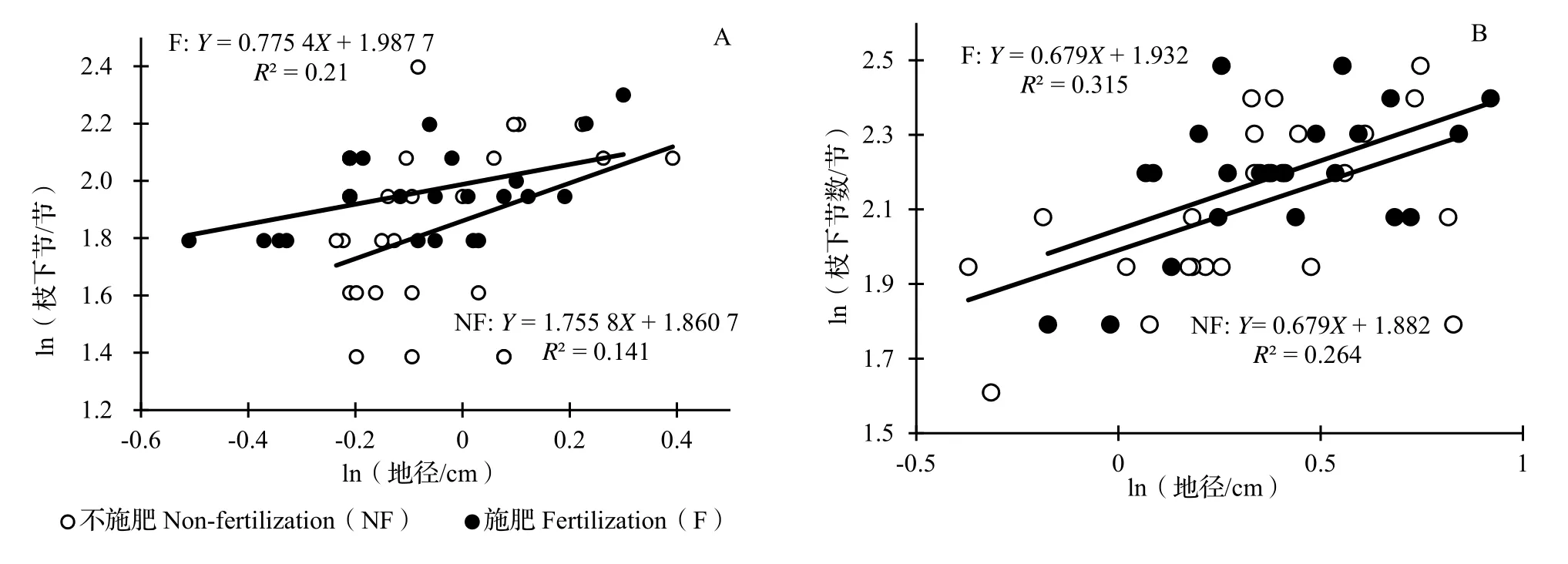
图4 不遮阴(A)、遮阴(B)处理下不同施肥水平水竹地径与枝下节数的异速生长关系Fig. 4 Allometric relationship of ground diameter with clear bole nodes of Ph. heteroclada with different fertilization under no shading (A)and shading (B)
在不遮阴处理下,施肥与不施肥水竹地径与枝下节数的异速生长分析可见,两者的异速生长指数没有显著差异(P= 0.830,图4B),共同斜率为0.679 0 (95%CI= 0.524 4 ~ 0.879 3,P< 0.001),显著小于1.0(P< 0.001)。进一步分析SMA主轴在Y轴方向和斜率方向的移动可以看到,在施肥和不施肥处理下沿Y轴和斜率方向的SMA主轴移动量均未达到显著水平(P= 0.554 > 0.05,P= 0.201 > 0.05)。表明在本试验中,施肥处理下水竹的地径与枝下节数与不施肥理下的地径与枝下节数没有显著变化。
比较图4A 与图4B 可以看出,不管是施肥或不施肥处理,不遮阴和遮阴水竹的地径、枝下节数异速生长关系的斜率之间均存在显著差异(SMA 分析,P= 0.001),前者显著大于1.0(P< 0.001),后者显著小于1.0(P< 0.001)。
2.3 遮阴和灌溉对水竹秆形特征异速生长关系的影响
2.3.1 不同遮阴处理下灌溉对水竹地径、株高异速生长关系的影响 在不遮阴处理下,灌溉和不灌溉处理下水竹地径与株高的异速生长指数没有显著差异(P= 0.594,图5A),共同斜率为1.188(95%CI= 0.900 ~ 1.567,P< 0.001),显著大于1.0(P< 0.001),表明不同水分条件未改变地径和株高的异速生长关系。进一步分析SMA主轴在Y轴方向和斜率方向的移动可以看到,灌溉和不灌溉处理下沿Y轴移动的SMA 截距没有达到显著水平(P= 0.668 > 0.05),但灌溉处理下水竹沿斜率方向出现显著性移动(P< 0.001)。表明在本试验中,灌溉处理下水竹的地径与株高整体比不灌溉处理下水竹的地径与株高更大。
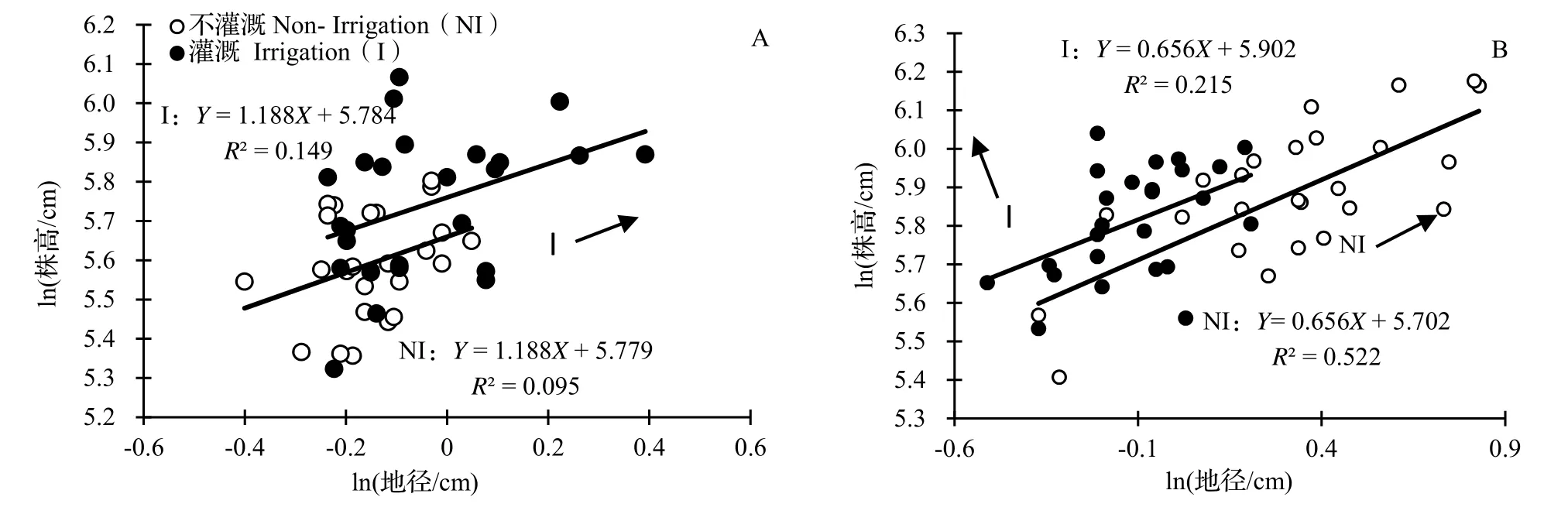
图5 不遮阴(A)、遮阴(B)处理下灌溉水平的水竹地径与株高的异速生长关系Fig. 5 Allometric relationship of ground diameter with height of Ph. heteroclada with different irrigation treatments under no shading (A) and shading (B)
在遮阴处理中,灌溉与不灌溉处理下水竹地径、株高异速生长关系的斜率之间没有显著(P= 0.156,图5B),共同斜率为0.656 0(95%CI= 0.519 5 ~ 0.834 3,P< 0.001),显著小于1.0(P< 0.001),表明不同灌溉处理未改变地径和株高的异速生长关系。进一步分析SMA 主轴在Y 轴方向的移动可以看到,灌溉处理下SMA 的截距沿着Y 轴出现了显著的正向移动(移动量0.208,Wald 检验,P< 0.001),表明在同等地径下,灌溉处理下水竹的株高大于不灌溉处理下水竹的株高。在沿共同斜率方向的移动上,与灌溉处理的水竹相比,不灌溉处理发生了正向的极显著移动(P< 0.001),表明整体上不灌溉处理下水竹的地径与株高比灌溉处理下的地径与株高更大。比较图5A 与图5B 可以看出, 不管是灌溉或不灌溉处理,不遮阴和遮阴水竹地径、株高异速生长关系的斜率之间均存在显著差异(SMA 分析,P< 0.001),且前者大于1.0(P< 0.001),后者小于1.0(P< 0.001)。
2.3.2 不同遮阴处理下灌溉处理对水竹地径、枝下高异速生长关系的影响 在不遮阴处理下,由灌溉与不灌溉处理下水竹地径与枝下高的异速生长分析可见,灌溉和不灌溉处理下水竹地径与枝下高异速生长指数没有显著差异(P= 0.729,图6A),共同斜率为3.310(95%CI= 2.494 ~ 4.391,P< 0.001),显著大于1.0(P< 0.001)。进一步分析SMA 主轴在Y 轴方向和斜率方向的移动可以看到,在灌溉和不灌溉处理下,沿Y 轴移动的SMA截距没有达到显著水平(P= 0.371 > 0.05),但在灌溉处理下水竹沿斜率方向出现显著性移动(P< 0.05)。以上结果表明在本试验调查中,灌溉处理下水竹的地径与枝下高整体比不灌溉处理下的地径与枝下高更大。
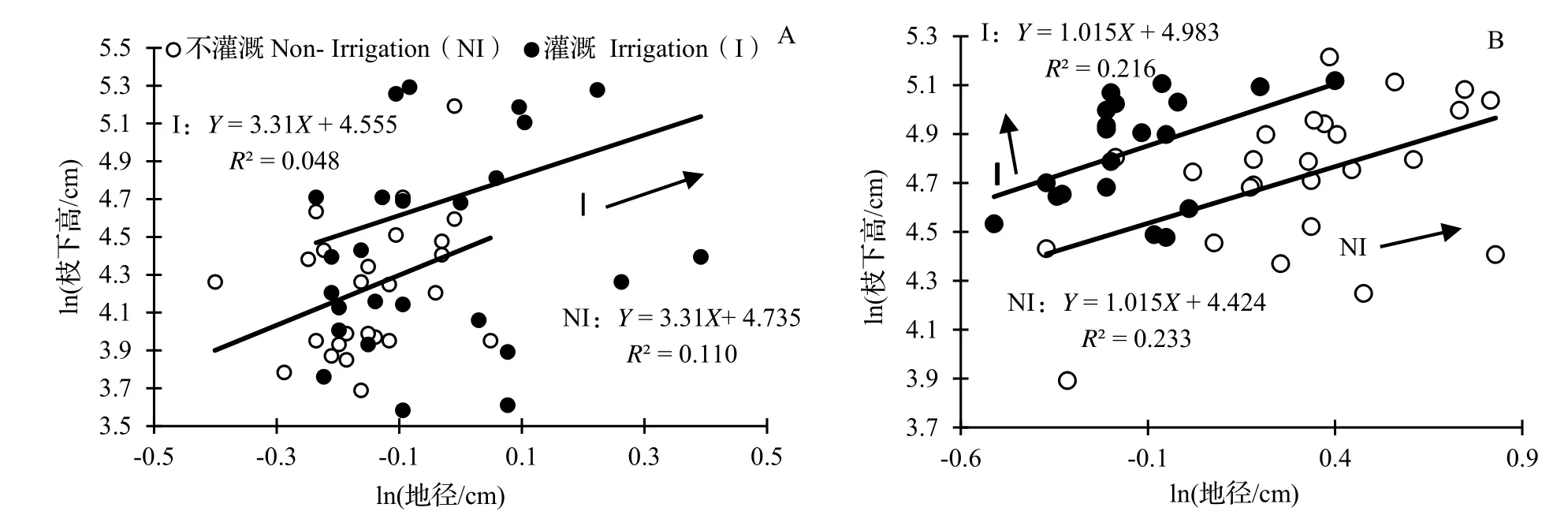
图6 不遮阴(A)、遮阴(B)处理下灌溉水平的水竹地径与枝下高的异速生长关系Fig. 6 Allometric relationship of ground diameter with clear bole height of Ph. heteroclada with different irrigation treatments under no shading (A) and shading (B)
在遮阴处理下,由不同灌溉处理下水竹地径与枝下高的异速生长分析可见,灌溉与不灌溉处理水竹的地径、枝下高SMA 斜率之间差异不显著(P= 0.658,图6B),共同斜率为1.015(95%CI= 0.768 8 ~ 1.337 4,P< 0.001),与1.0 没有显著差异(P> 0.05)。在Y 轴方向上进一步分析得到,灌溉处理下SMA 主轴在Y 轴方向出现了正向的极显著移动(P< 0.001),移动量是0.567,即1.763 cm,表明在同等地径下,灌溉处理水竹的枝下高大于不灌溉处理水竹的枝下高。在沿共同斜率方向的移动上,灌溉和不灌溉的移动量也达到显著水平(P< 0.05),表明整体上不灌溉处理水竹的地径与枝下高比灌溉处理水竹的地径与枝下高更大。比较图6A 与图6B 可以看出,不管是灌溉或不灌溉、遮阴或不遮阴处理,水竹地径、枝下高异速生长关系的斜率之间均存在显著差异(SMA分析,P= 0.001),前者显著小于1.0(P< 0.001),后者与1.0 无显著差异(P> 0.05)。
2.3.3 不同遮阴处理下灌溉对水竹地径、枝下节数异速生长关系的影响 在不遮阴处理下,灌溉和不灌溉处理下水竹的地径与枝下节数异速生长指数没有显著差异(P= 0.679,图7A),共同斜率为1.864(95%CI= 1.413~ 2.459,P< 0.001),显著大于1.0(P< 0.001)。进一步分析SMA 主轴在Y 轴方向和斜率方向的移动可以看到,灌溉和不灌溉处理下SMA 主轴沿Y 轴的移动量没有达到显著水平(P= 0.101 > 0.05),但灌溉处理下水竹沿斜率方向出现显著性移动(P< 0.05)。表明在本试验中,灌溉处理下水竹的地径与枝下节数整体比不灌溉处理下水竹的地径与枝下节数更大、更多。在遮阴处理中,灌溉与不灌溉处理水竹地径、枝下节数异速生长关系的斜率之间差异不显著(P= 0.772,图7A),共同斜率为0.725 6(95%CI= 0.559 0 ~ 0.942 0,P< 0.001),显著大于1.0(P< 0.001)。进一步分析SMA 主轴在Y 轴方向和斜率方向的移动可以看到,灌溉处理下SMA 截距沿着Y 轴出现了显著的正向移动(移动量0.143,Wald 检验,P< 0.05),表明在同等地径下,水竹枝下节数不灌溉处理大于灌溉处理。在沿共同斜率方向的移动上,灌溉和不灌溉的SMA 主轴的移动量也达到极显著水平(P<0.001),表明整体上不灌溉处理下水竹的地径与枝下节数比灌溉处理下水竹的地径与枝下节数更大和更多。

图7 不遮阴(A)、遮阴(B)处理下灌溉水平水竹地径与枝下节数的异速生长关系Fig. 7 Allometric relationship of ground diameter with clear bole nodes of Ph. heteroclada with different irrigation treatments under no shading (A) and shading (B)
比较图7A 与图7B 可以看出,不管是灌溉或不灌溉处理,不遮阴和遮阴处理,水竹地径、枝下节数异速生长关系的斜率之间均存在显著差异(SMA 分析,P< 0.001),前者大于1.0(P< 0.001),后者小于1.0(P< 0.001)。
3 讨论
植物的表型可塑性是适应异质环境的一个重要特征,使植物能够把资源最佳地分配到不同的器官,以避免或降低相邻植物的竞争[9]。异速生长是生物体不同性状对环境选择压力采取的生态适应策略[10],植物通过光合作用形成的光合产物主要用于生存、繁殖等方面,但是可利用的光合产物是有限的,因此在植物体内各功能构件之间的资源分配常表现为相互矛盾[11],这些相互矛盾的需求产生了多种权衡策略[12],异速生长就是这种权衡最直接的表现形式之一[13]。同时,植物异速生长的稳定性也受光照、水分、养分等生态因子组合与变异的制约。通过所有水竹地径与株高、枝下高、枝下节数数据计算共同斜率,线性拟合较好,三者关系均呈极显著的正相关关系(P< 0.001),符合异速生长关系,但地径与枝下高、枝下节数图中部分点出现大幅度偏离,导致散点图呈现“喇叭状”,这主要与不同施肥、遮阴、灌溉处理引起环境条件的变化和异质生境中水竹的适应策略有关,原因有待下一步深入探讨。
3.1 施肥对水竹地径、株高、枝下高、枝下节数的影响
施肥是提高竹林生产力的重要举措。本研究中,与不施肥相比,施肥处理下水竹地径与枝下高、枝下节数标准主轴的斜率更小,但施肥未显著改变地径与株高的异速生长斜率。这与庄若楠[14]对毛竹Phyllostachysedulis的研究结果一致。施肥导致光合产物在枝下高和枝下节数上的积累减少与施肥增加的光合产物分配有关。王春胜等[15]对西南桦Betulaalnoides研究认为相对于冠幅而言,胸径、单株材积等构件因子对施肥的响应较慢。对应于本研究的水竹而言,施肥将光合产物可能相对更多地投入到冠幅中,叶面积指数变大,有利于提高光能的利用率[16],而枝下高、枝下节数降低也有利于节约养分向冠层下部运输的成本,导致施肥处理的地径与枝下高、枝下节数的异速生长斜率小于不施肥的。另外,也可能和水竹分株之间的生理整合有关,克隆分株间通过联系通道产生非对称性竞争。
施肥处理样地4 的植株密度最大,资源竞争激烈,使植株产生自疏效应,促使其趋向于矮化生长状态,即地径与枝下高、枝下节数的异速生长指数下降,因此,施肥的枝下高、枝下节数生长速率低于地径的生长速率。
施肥未显著改变水竹地径与株高的标准主轴斜率,原因是竹类植物的株高和地径的可塑性较小,茎秆的粗细在生长初期就已经确定,在成竹后高度一般不再增加[14],所以地径与株高SMA 斜率相对稳定。但施肥SMA主轴在Y 轴方向和斜率方向出现正向显著移动,可能是施肥后水竹将增加的总初级生产量绝大部分用于地上部分的生长,使得整体上施肥处理水竹的地径与株高比不施肥处理水竹的更大。
3.2 灌溉对水竹地径、株高、枝下高、枝下节数的影响
林地的土壤水分状况是影响竹类植物造林成活率的重要因素。水分处理未显著改变水竹地径与株高、枝下高、枝下节数异速生长的SMA 斜率,这与水竹对水分生理整合的利用策略有关。童姹紫[17]研究认为在干旱环境下,竹鞭通过生理整合功能为维持新竹的高蒸腾需求提供了水分支持,并通过夜间液流对毛竹茎秆储存水进行补充,这一机制有助于在干旱条件下缓解水竹对水分的需求。本研究发现,灌溉虽然不能显著改变水竹地径与株高、枝下高、枝下节数的异速生长关系,但在不同光照条件下存在差异。
在不遮阴条件下,灌溉处理下SMA 主轴沿共同斜率方向出现显著移动,表明与不灌溉处理相比,灌溉处理下水竹将更多同化物转移到地径、株高、枝下高、枝下节数。韩学琴等[18]对辣木Moringaolerfera研究认为在水分充足时生长趋向于将更多的光合产物投入株高生长,以增强垂直方向生长和提高获取光合的能力为主要生长策略。在本研究中,水竹在灌溉处理时,将较多的资源分配至地上部分,提高地径、株高、枝下高、枝下节数的相对生长速度,满足其正常生长的需要。
在遮阴条件下,表现为在同等地径下,灌溉处理下水竹具有更高的株高和枝下高,但由于不灌溉处理下增加了地径、株高、枝下高增长(水分缺少沿着主轴斜率正向移动),导致不灌溉处理下水竹地径、株高的增长速度高于灌溉处理的地径、株高的增长速度。对于枝下节数而言,整体上不灌溉水竹的地径与枝下节数比灌溉处理的地径与枝下节数更大、更多。潘玉梅等[19]认为,与水分因子相比,光照因子对植株形态特征和生物量的影响更显著。这种遮阴后增加了秆形因子的生长速度,水竹为增强在弱光环境中对光资源的竞争能力,通过改变水分整合强度的表型可塑性[20],削弱了水分对秆形特征的影响,可能将更多的光合产物投入到株高、枝下高、枝下节数的生长中,减小了对其他构件投资造成的不必要损失。
3.3 遮阴对水竹地径、株高、枝下高、枝下节数的影响
光是植物进行光合作用和生长发育的基本条件。本研究中,无论是施肥或不施肥、灌溉或不灌溉,遮阴均使水竹地径与株高、枝下高、枝下节数异速生长的SMA 斜率显著降低,即遮阴使株高、枝下高、枝下节数的生长速率低于地径的生长速率,且这种响应在不施肥、不灌溉下更为强烈。这可能与遮阴条件下光合碳摄取不足、代谢运输受阻有关[21]。遮阴会降低水分蒸散速率, 使输送到冠层上部叶片的水分减少,同时也限制了外部二氧化碳进入叶片,也会影响光合产物的分配和可用性,光合碳摄取不足影响到水竹对水分和养分的吸收、运输、利用及其对环境的适应能力,导致库存碳水化合物输导功能降低和需求减少[22],最终使生长受限。水分和养分受限制时,水竹秆形的可塑性响应表现得更为剧烈。
4 结论
不考虑遮阴、施肥和灌溉处理下,水竹地径与株高、枝下高、枝下节数的关系均呈显著的正相关关系。通过6 个样地的比较,遮阴和施肥对水竹秆形特征及其异速生长关系产生了强烈的影响,斜率变异分别可达227%和210%,而灌溉没有改变其异速生长关系,综合影响程度表现为遮阴>施肥>灌溉,水竹秆形生长可以通过其秆形的可塑性调节适应不同的经营措施。
