结直肠癌干细胞中miR-520c-3p的表达及其对丙酮酸代谢的调控机制研究
2023-12-11赵英旋白玉勤欧阳卫东
赵英旋,白玉勤,欧阳卫东
(1.赤峰学院附属医院 普外四科,内蒙古 赤峰 024005;2.内蒙古赤峰市肿瘤医院 病理科,内蒙古 赤峰 024005)
丙酮酸的代谢途径取决于细胞当时的条件以及有机体种类或组织的类型[1]。大多数分化的哺乳动物细胞将丙酮酸导入线粒体,在那里被氧化以产生高效的三磷酸腺苷(adenosine triphosphate,ATP)[2]。然而,癌细胞将丙酮酸及其前体转用于其他合成代谢过程,或将其转化为乳酸排出细胞外[3]。这种代谢适应是由生物化学家Otto Warburg在20世纪20年代首次描述的,被称为Warburg 效应。多种机制促成了癌症的这种代谢失调,但丙酮酸的合成和代谢起着核心作用[4-7]。丙酮酸代谢的改变似乎是促成和促进许多癌症发生发展的关键,包括结直肠癌[8-12]。但是,目前对这一过程在分子水平上的调控研究很少。microRNA(miRNA)是一种长度约为19~24 nt 的内源性非编码小RNA,通过靶向靶基因mRNA 的3'端非编码区,引起转录抑制或调节mRNA降解[13-14]。Deng等[15]报道,miR-23a在结直肠癌干细胞中的过表达间接促进了参与氧化磷酸化的丙酮酸脱氢酶(pyruvate dehydrogenase,PDH)的激活,从而产生足够的ATP 用于肿瘤细胞增殖。然而,结直肠癌发生发展中存在多种miRNA 的表达失调,这种失调是否影响结直肠癌干细胞的丙酮酸代谢仍然未知。本研究通过TCGA数据库发现miR-520c-3p 在结直肠癌中表达上调,并且高表达miR-520c-3p 的结直肠癌患者有较差的预后。但是,miR-520c-3p 是否参与丙酮酸代谢尚不清楚。基于此,本研究旨在探讨miR-520c-3p 调控结直肠癌干细胞丙酮酸代谢的潜在机制。
1 材料与方法
1.1 材料
HCT-116 细胞株购自宝如亿(北京)生物技术有限公司(货号:RX1525)。HT-29 细胞株购自深圳市文乐生物科技有限公司(货号:iCell-h078)。SW-1116 细胞株购自上海信裕生物科技有限公司(货号:26-5140)。DMEM 培养基购自北京赛音图科技有限公司(货号:C11995500BT)。MPC1 抗体购自广州吉英生物科技有限公司(货号:42898)。GAPDH 抗体购自艾比玛特医药科技(上海)有限公司(货号:BCJ200003H)。miR-520c-3p 模拟物(mimics)和miR-520c-3p 抑制物(inhibitor)由北京擎科设计合成。含MPC1 抑制物(shMPC1)和阴性对照序列(shNC) 的慢病毒由汉坦生物提供。L-乳酸检测试剂盒购自北京轩泽伟业科技有限公司(货号:K-LATE)。葡萄糖检测试剂盒购自北京奇松生物科技有限公司(货号:BQS118529-100T)。CyQuant 细胞增殖测定试剂盒购自ThermoFisher 公司(货号:C7026)。RNeasy Mini Kit 购自北京强欣博瑞生物技术有限公司(货号:74134)。高容量cDNA 反转录试剂盒购自ThermoFisher 公司(货号:4374967)。SYBR Green I Master Mix 购自北京擎科生物科技有限公司(货号:TSE203)。
1.2 方法
1.2.1 细胞培养和处理根据文献[16],应用无血清的肿瘤干细胞培养基分别从HCT-116、HT-29、SW-1116 细胞株中分离纯化人结直肠癌干细胞株 HCT-116CSC、 HT-29CSC、 SW-1116CSC。HCT-116CSC、HT-29CSC、SW-1116CSC 置于含各种生长因子无血清DMEM 培养基中培养;HCT-116、HT-29、SW-1116、细胞培养于含100 mL/L 胎牛血清的DMEM 培养基中。所有细胞在含5% CO2的37 ℃培养箱中培养。
1.2.2 TCGA 数据库数据集分析根据TCGA 数据库(http://cancergenome.nih.gov) 数据集 COAD 和COADREAD,利用Survexpress 比较目标基因高表达和低表达的结直肠癌患者的生存状况。
1.2.3 葡萄糖和乳酸含量的检测乳酸含量通过使用L-乳酸检测试剂盒进行测定。葡萄糖通过葡萄糖检测试剂盒进行检测。将结直肠癌干细胞HCT-116CSC、HT-29CSC、SW-1116CSC 在DMEM 基本培养基中培养,然后根据制造商的说明收集细胞并进行乳酸或葡萄糖含量测定。
1.2.4 Western blot结直肠癌干细胞使用裂解缓冲液50 mmol/L HEPES,150 mmol/L NaCl,10% Glycerol,1% Triton X-100,1.5 mmol/L MgCl,1.0 mmol/L EGTA,10 mmol/L 焦磷酸钠,100 mmol/L 氟化钠,1 mmol/L苯甲基磺酰氟(PMSF)裂解。样品的蛋白质浓度通过BCA 检测进行量化,并进行归一化处理,以确保各孔之间的样品负荷恒定。然后对样品进行SDS-聚丙烯酰胺凝胶电泳(PAGE)和免疫印迹。用18%丙烯酰胺凝胶进行解析,并转移到甲醇激活的0.45 μm Immobilon FL PVDF 膜上。在2.5%的牛奶中阻断1 h 后,清洗膜并使用在含有0.1% Tween-20 的TBS 中制成的5% BSA 溶液中稀释的特异性抗体进行检测。洗涤膜并使用荧光共轭二抗进行探测,并使用LI-COR Biosciences Odyssey 系统进行扫描。
1.2.5 质量同位素分析所有的13C 研究都是在含有10%透析FBS 的培养基中进行的,准备时要使葡萄糖或谷氨酰胺池100%被标记为13C,而另一个池则未被标记。缺少葡萄糖和谷氨酰胺的DMEM 由粉末制备,然后补充15 mmol/LD-(U-13C)葡萄糖。细胞在60 或100 mm 的盘子里生长到80%汇合,然后用PBS 冲洗,用含13C 的培养基培养6 h。然后在0.5 mL 1∶1 的冷甲醇:水的混合物中冻融3 次,提取细胞。通过离心除去大分子和碎片,加入50 nmoL 的2-氧代丁酸,完全蒸发带有水样代谢物的上清液,并在42 ℃下在100 μL 的三甲基硅基供体中衍生30 min。使用与Agilent 5973 质量选择检测器联网的Agilent 6970 气相色谱仪对代谢物进行分析。所有代谢物的保留时间和质量碎片特征都用纯标准品进行验证。为了确定不同样品的相对代谢物丰度,将代谢物的总离子流峰的面积与内标的面积进行比较,并对蛋白质含量进行归一化。质量同位素分布分析测量了每个代谢物池中包含所有可能数量的13C 原子的部分;也就是说,一个代谢物可能包含0、1、2...n个13C 原子,其中n=代谢物中碳原子的数量。对于每个代谢物,使用MSDChem 软件分析含有母分子中所有碳原子的信息碎片离子,整合从m+0 到m+n的所有质量同位素的丰度,其中m=没有任何13C 的碎片离子的质量。然后对每个质量同位素的丰度进行数学校正,以考虑到自然丰度同位素,最后转化为总库的百分比。
1.2.6 细胞耗氧量的测定代谢测量是在标准的24 孔Seahorse 微孔板上在Seahorse XF24 分析仪上进行的。糖酵解是根据糖酵解应激测试套件测量的,并显示为细胞外酸化率(extracellular acidification rate,ECAR)。当细胞在含有10 mmol/L 丙酮酸钠作为唯一呼吸底物培养时,使用氧消耗率(oxygen consumption rate,OCR)测量丙酮酸的氧化。对于所有的实验,在分析前16~18 h,每孔有8 万个细胞。
1.2.7 RNA 抽取、miRNA 测序和实时荧光定量PCR根据制造商的说明,使用RNeasy Mini Kit 从细胞中提取和纯化总RNA。对于miRNA 测序,纯化的总RNA 交由华大基因测序。对于实时荧光定量PCR,使用高容量cDNA 反转录试剂盒合成cDNA。随后,用LightCycler 480 SYBR Green I Master Mix 进行实时PCR。所有反应至少进行了4 次。
1.2.8 细胞增殖水平的测定细胞增殖是通过在96 孔板中每孔分选5 000 个单细胞进行的。3 d 内每24 h 抽吸1 次培养基,用PBS 清洗2 次,然后储存在-80 °C,收获平板。使用CyQuant 细胞增殖测定试剂盒测定增殖,使用Varioskan Flash 读片机测定荧光。
1.3 统计学处理
用GraphPad 5.0 软件,通过单因素方差分析或t检验来分析各组之间的差异。P<0.05 为差异有统计学意义。
2 结果
2.1 miR-520c-3p表达对结直癌干细胞增殖的影响
过表达或敲低miR-520c-3p 后,结直肠癌细胞(HCT-116、HT-29、SW-1116) 的增殖水平均无明显变化(均P>0.05)。过表达miR-520c-3p 后,结直肠癌干细胞 (HCT-116CSC、 HT-29CSC、 SW-1116CSC)的增殖水平均明显增强(均P<0.05);敲低miR-520c-3p 后,结直肠癌干细胞HCT-116CSC、HT-29CSC、SW-1116CSC 的增殖水平明显降低(均P<0.05)(图1)。
2.2 miR-520c-3p表达对结直肠癌干细胞丙酮酸代谢的影响
过表达或敲低miR-520c-3p 后,结直肠癌细胞的丙酮酸氧化水平均无明显变化(均P>0.05)。过表达miR-520c-3p 后,结直肠癌干细胞的丙酮酸氧化水平均明显下降(均P<0.05);敲低miR-520c-3p后,结直肠癌干细胞的丙酮酸氧化水平均明显上升(均P<0.05)(图2A)。用D-(U-13C)葡萄糖培养后,未标记的柠檬酸盐(m+0)在敲低miR-520c-3p的结直肠癌干细胞中均明显减少(均P<0.05),而高阶柠檬酸盐标记(m+1、m+4 和m+5)均明显增加(均P<0.05);未标记的柠檬酸盐(m+0)在过表达miR-520c-3p 的结直肠癌干细胞中均明显增加(均P<0.05),而高阶柠檬酸盐标记(m+1、m+4 和m+5) 均明显减少(均P<0.05)(图2B-D)。过表达miR-520c-3p 后,结直肠癌干细胞产生的乳酸均明显增高(均P<0.05);敲低miR-520c-3p 后,结直肠癌干细胞产生的乳酸均影响减少(均P<0.05)(图2E-G)。
2.3 miR-520c-3p的靶基因分析与鉴定
过表达或敲低miR-520c-3p 后,结直肠癌干细胞SW-1116CSC 中多种基因的转录水平受到调控,经过生物信息学软件预测发现miR-520c-3p 可以与线粒体丙酮酸载体1(mitochondrial pyruvate carrier 1,MPC1)mRNA 3'UTR 相结合(图3A-B)。荧光素酶报告实验发现miR-520c-3p 可以靶向MPC1 mRNA 3'UTR (P<0.05)(图3C)。过表达miR-520c-3p 后,结直肠癌干细胞中MPC1 的mRNA 和蛋白水平均明显下降(均P<0.05);敲低miR-520c-3p 后,结直肠癌干细胞中MPC1 的mRNA 和蛋白水平均明显上升(均P<0.05)(图3D-E)。同时,TCGA 数据库结直肠癌数据集COAD 和COADREAD 中低表达MPC1 的结直肠癌患者有较差的预后(P<0.05)(图3F)。敲低MPC1 后,结直肠癌干细胞的丙酮酸氧化水平均明显下降(均P<0.05),而同时敲低miR-520c-3p和MPC1 后结直肠癌干细胞丙酮酸氧化水平均无明显变化(均P>0.05)(图3G)。用D-(U-13C)葡萄糖培养后,未标记的柠檬酸盐(m+0) 在敲低MPC1 的结直肠癌干细胞中均明显增加(均P<0.05),而高阶柠檬酸盐标记(m+1、m+4 和m+5)均明显减少(均P<0.05)(图3H-J)。敲低MPC1 后,结直肠癌干细胞产生的乳酸均明显增高均(P<0.05),而同时敲低miR-520c-3p 和MPC1 后结直肠癌干细胞的乳酸水平无明显变化(均P<0.05)(图3K-M)。敲低MPC1 后,结直肠癌干细胞产生的增殖能力均明显增强(均P<0.05),而同时敲低miR-520c-3p 和MPC1 后结直肠癌干细胞的增殖水平无明显变化(均P>0.05)(图3N-P)。
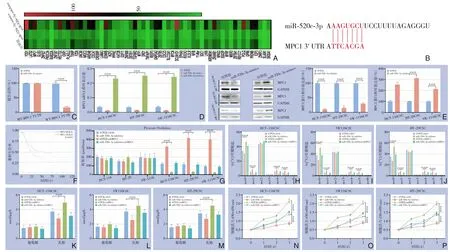
图3 miR-520c-3p 靶向MPC1 调控结直肠癌干细胞丙酮酸代谢 A:过表达或敲低miR-520c-3p 后,结直肠癌干细胞SW-1116CSC的转录组热图;B:生物信息学软件预测miR-520c-3p与MPC1 mRNA 3'UTR的结合位点;C:荧光素酶报告实验检测miR-520c-3p 与MPC1 mRNA 3'UTR 的结合;D-E:过表达或敲低miR-520c-3p 后,结直肠癌细胞中MPC1 的mRNA和蛋白质表达水平;F:MPC1不同表达水平对结直肠癌生存曲线的影响;G:图中所示处理下,结直肠癌细胞以及结直肠癌干细胞的丙酮酸氧化水平;H-J:各种处理下,D-(U-13C)葡萄糖培养后质量同位素分析柠檬酸盐(m+0、m+1、m+2、m+3、m+4、m+5、m+6);K-M:各种处理下,结直肠癌干细胞的乳酸和葡萄糖水平;N-P:各种处理下,结直肠癌干细胞的增殖水平Figure 3 MiR-520c-3p regulating pyruvate metabolism of colorectal cancer stem cells by targeting MPC1 A:Heatmap of transcriptome analysis in SW-1116CSC colorectal cancer stem cells with overexpression or knockdown of miR-520c-3p;B:Prediction of binding sites between miR-520c-3p and MPC1 mRNA 3'UTR using bioinformatics software;C:Luciferase reporter assay to detect the binding of miR-520c-3p to MPC1 mRNA 3'UTR;D-E:The mRNA and protein expression levels of MPC1 in colorectal cancer cells after overexpression or knockdown of miR-520c-3p;F:Ⅰmpact of different expression levels of MPC1 on survival curve of colorectal cancer;G:Pyruvate oxidation levels in colorectal cancer cells and colorectal cancer stem cells under various treatments;H-J:Mass isotopomer analysis of citrate(m+0,m+1,m+2,m+3,m+4,m+5,m+6)after D-(U-13C)glucose culture in different treatments;K-M:Lactate and glucose levels in colorectal cancer stem cells under various treatments;N-P:Proliferation levels of colorectal cancer stem cells under various treatments
3 讨论
在全世界的癌症发病率和病死率排名中,结直肠癌分别位列第三和第四[17-20]。每年有超过100 万的结直肠癌新病例,大约有70 万例死于该病[21]。多种因素参与结直肠癌的发生和发展,包括致癌基因的激活和肿瘤抑制基因的失活[22-24]。清楚地了解结直肠癌的发生、发展、迁移和复发的机制,将大大有助于结直肠癌的早期诊断和治疗。本研究通过前期TCGA 数据集分析与后期功能实验发现,miR-520c-3p/MPC1 轴调控结直肠癌干细胞中的丙酮酸代谢水平。
大多数分化的细胞通过糖酵解在细胞液中把葡萄糖转化为丙酮酸,然后在线粒体中进行丙酮酸氧化[25]。相反,增殖型细胞,包括许多肿瘤细胞和干细胞,有力地进行糖酵解,但限制了线粒体丙酮酸氧化的比例[26]。本研究发现过表达miR-520c-3p 后,结直肠癌干细胞的增殖水平显著上升、丙酮酸氧化水平显著下降、乳酸显著增高。用D- (U-13C)葡萄糖培养后,未标记的柠檬酸盐(m+0)在过表达miR-520c-3p 的结直肠癌干细胞中增多,而高阶柠檬酸盐标记(m+1、m+4 和m+5)减少。含有高阶标记的柠檬酸盐是由于葡萄糖衍生的丙酮酸转移到线粒体,然后丙酮酸脱氢酶、丙酮酸羧化酶和三羧酸循环的活动而产生的。在较短的标记时间内,标记的差异更为明显,然而,m+0 柠檬酸盐的增多表明葡萄糖对TCA 循环的贡献率减少。由于柠檬酸盐中13C 的积累导致m+1、m+4 和m+5 的丰度增加,m+0 柠檬酸盐随着miR-520c-3p 通量的增加而明显增加,但m+2 和m+6 柠檬酸盐的丰度变化非常小。这些数据表明,miR-520c-3p 表达减少了丙酮酸通量进入线粒体,减少了丙酮酸氧化,减少了三羧酸循环中乙酰辅酶A 的水平。
本研究发现miR-520c-3p 可以靶向MPC1 mRNA 3'UTR。过表达miR-520c-3p 后,结直肠癌干细胞中MPC1 的mRNA 水平和蛋白水平显著下降。同时,本研究的数据显示,敲低MPC1 的结直肠癌干细胞中线粒体代谢发生了改变,增加了细胞内丙酮酸和乳酸的数量。MPC1 是一种将丙酮酸转移到线粒体的内膜蛋白,抑制MPC1 的表达会使癌细胞转向上皮细胞-间充质转化和谷氨酸溶解[27]。谷氨酰胺分解的增加是由于旁路机制补偿了线粒体中丙酮酸的损失,并通过谷氨酰胺流入维持三羧酸循环中间产物的补充[28]。尽管丙酮酸被认为是一种抗氧化分子,但由于丙酮酸转化为乳酸并外流到细胞外空间,可能无法有效发挥作用[29]。此外,通过抑制胱氨酸/谷氨酸逆向转运蛋白溶质载体家族7 成员 11 (solute carrier family 7 member 11,SLC7A11,也称xCT) 消耗谷胱甘肽(glutathione,GSH)也会增加线粒体的代谢,从而导致MPC1 沉默的癌细胞的铁中毒敏感性增加[30]。因此,抑制MPC1 的表达会改变结直肠干细胞中丙酮酸代谢。
综上所述,结直肠癌中高表达的miR-520c-3p与较差的预后相关,其作用机制可能与miR-520c-3p靶向MPC1 调控结直肠癌干细胞中的丙酮酸代谢水平,促进了结直肠癌干细胞的增殖有关。
利益冲突:所有作者均声明不存在利益冲突。
作者贡献声明:赵英旋负责科研设计、数据分析;白玉勤负责病例收集、文章写作修改;欧阳卫东负责指导以及文章修改。
