茂兰圆果化香群落主要种群生态位特征和种间联结性分析
2023-12-09罗小菊李秋英吴高殷刘映良
谭 娅,罗小菊,李秋英,王 爽,吴高殷,刘映良*
(1.贵州师范大学 生命科学学院,贵州 贵阳 550025;2.贵州师范大学 图书馆,贵州 贵阳 550025)
生态位理论和种间联结性研究是种群生态学和群落生态学的重要内容[1]。生态位理论广泛应用于种间关系、群落结构、生物多样性和种群进化[2]及物种的共存和竞争机制[3]等研究中。种间关系则反映物种空间上的分布模式及生境对其影响[4]。运用生态位和种间联结理论,有学者探讨滨海沙地人工防护林群落[5]及入侵植物群落[6]的稳定性,并研究天然林群落不同阶段的动态变化,也有学者发现黄土高原天然次生林中辽东栎(Quercuswutaishanica)种群正在扩大并占主导地位[7]。这些研究有助于理解群落的物种组成、确立种间关系、保护物种多样性、应对环境对物种的影响及物种对环境的作用,对植被资源保护、可持续利用和植被恢复重建等具有重要意义。
圆果化香(Platycaryalongipes)属胡桃科(Juglandaceae)化香属(Platycarya)落叶阔叶小乔木,基数羽状复叶,主要分布于贵州、广西、广东等地[8],是喀斯特森林生态系统的主要建群种之一,对于喀斯特森林生态系统的构建、稳定性起着重要作用,特别是对于喀斯特地区先锋物种群落的形成、维持和群落环境构建等起着至关重要的作用。前人对圆果化香药用成分、形态生理、群落结构、群落竞争、叶绿体基因组等[8-9]方面进行了研究。目前,关于圆果化香种群和群落的生态位与种间联结的相关研究未见报道。研究喀斯特地区圆果化香群落的生态位特征和种间关系,探讨群落的稳定性,为今后喀斯特区植被恢复及石漠化治理提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
茂兰自然保护区(25°09′20″-25°20′50″N,107°52′10″-108°45′40″E)位于贵州省黔南布依族苗族自治州荔波县南部。海拔430.0~1 078.6 m,保护区总面积212.85 km2,是典型的喀斯特峰丛地貌。地面岩石裸露率高,土层浅薄,土壤以黑色石灰土为主,持水量较低,地表水缺乏[10]。属中亚热带季风湿润气候,年平均气温15.3 ℃,全年降水量1 750~1 950 mm,年平均相对湿度83%。生长期237 d,年日照时数1 272.8 h。
1.2 样方选取与植被调查
以茂兰自然保护区为研究区域(图1),于2021年10-12月在该区域通过典型样地调查,以喀斯特森林的主要建群种圆果化香为研究对象,按照群落最小表现面积法设置大小为20 m×20 m的样地,共调查18个样地,每个样地设置10个4 m×10 m的样方。对样方内所有木本植物进行统计,内容包括种名、胸径、树高、冠幅及株数等。用GPS记录样地经纬度、海拔,测定其坡度、坡向、坡位和地貌类型等。
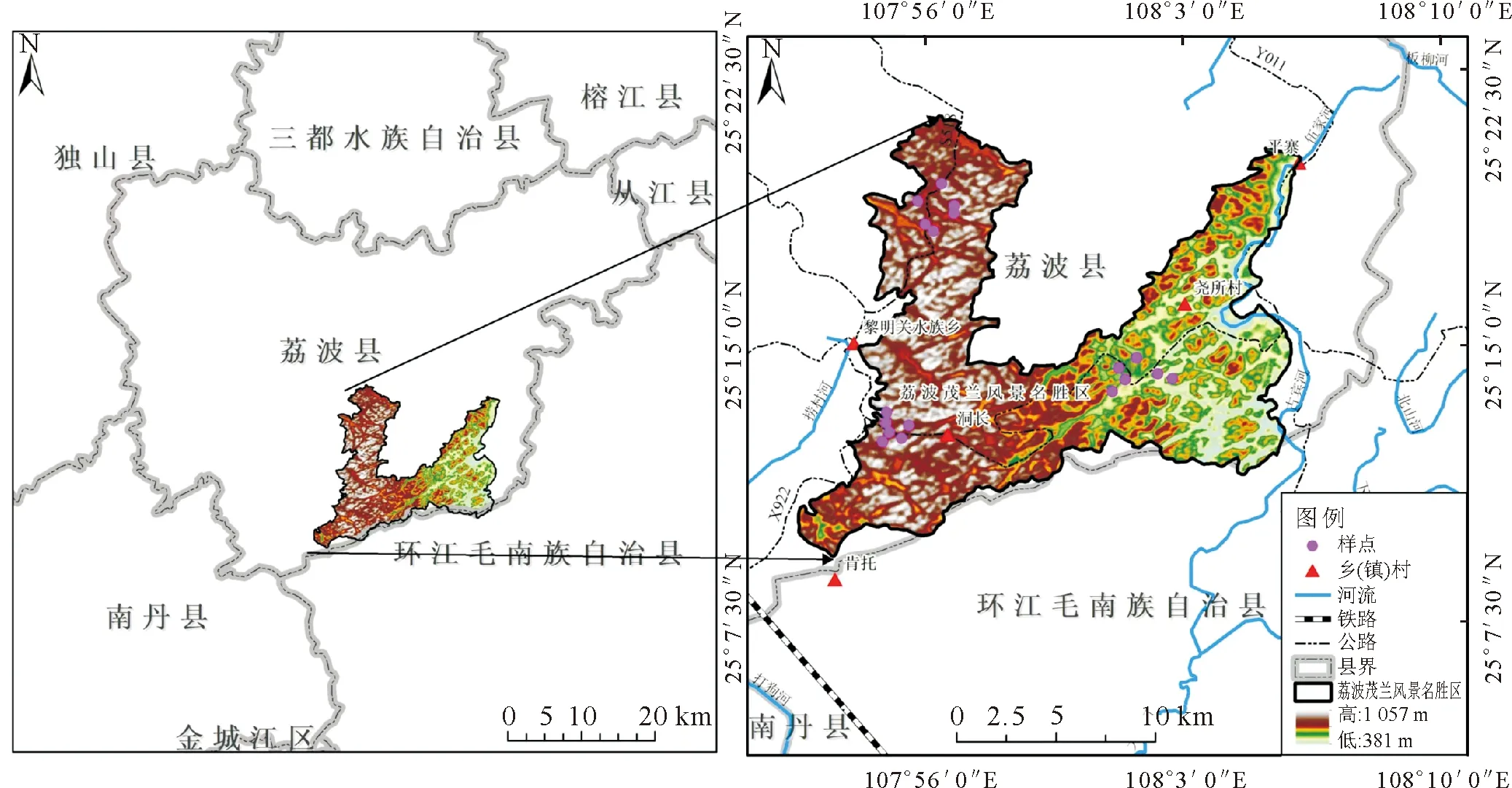
图1 研究区位置及样方布设Fig.1 Location of the study area and quadrats setting
1.3 数据分析
1.3.1 生态位特征分析 参考文献[11]计算各物种重要值并选取排序前15的物种进行生态位和种间联结性分析。选用Levins(BL)[12]和Shannon-Wiener(BS)[13]、Pianka生态位重叠(Oik)[14]及Schoener生态位相似性指数(Cik)[15]分析物种在群落中的生态位特征。
1.3.2 关联性检验与稳定性分析 利用方差比率法(VR)测定总体联结性[5],以统计量W来检验显著度。并运用贡献定律法[16]分析群落稳定性。通过X2检验种间联结关系[17],采用联结系数(AC)进一步测定种间关联程度[5]。
1.3.3 数据统计与分析 采用SPSS 26.0进行数据分析整理,运用Origin 2021绘图。
2 结果与分析
2.1 圆果化香群落主要物种重要值与生态位宽度分析
群落中共有木本植物4 816株,隶属49科88属130种。茂兰喀斯特森林15个优势树种重要值见表1,圆果化香的重要值和生态位宽度最高,为阳生性树种,生长在中坡和上坡,争夺阳光、空间等环境资源更为有利。重要值和生态位宽度最低的刺叶桂樱生长于山坡阳处疏密杂林或山谷沟边阴暗阔叶林及边缘,生态位较窄,对资源利用能力较弱。群落中主要物种的BL和BS在大小排序基本一致。粗糠柴和清香木在BS排序为第3、4位,而BL排序上则为第4、3位,可能是生境习性相似所致,两者在生态位宽度上差异不明显。

表1 圆果化香群落主要木本植物种群的重要值和生态位宽度Table 1 Importance values and niche breadth of the main woody plant populations in Platycarya longipes community
2.2 圆果化香群落生态位重叠与生态位相似性指数
群落中种对间的Oik都未>0.20(表2),<0.10的有95对,占90.48%,表明物种间利用相似资源程度不高,种间竞争并不激烈,在群落演替过程中物种的生态位倾向于特化。生态位完全相似(Oik=1.00)的种对有5个,占4.76%;物种间生态位相似比例较高(0.50≤Oik<1.00),占95.24%,生态位宽度大的物种生存能力强,分布广,数量多,在各个资源位处于优势地位,因此种对生态位相似性比例较大,如圆果化香与其他多数物种的生态位相似性指数>0.60。
2.3 圆果化香群落种间联结性与稳定性分析
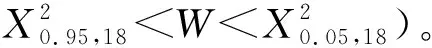

图2 圆果化香群落的稳定性Fig.2 Stability of P.longipes community
AC>0.6的种对有3对,AC<-0.6的有17对(图3)。由于圆果化香出现在所有样地中,故与小叶青冈、亮叶猴耳环和荔波鹅耳枥等14个优势种均无联结(AC=0)。0 本研究表明,群落中物种的生态位宽度取决于重要值大小及其频度、生境异质性程度。圆果化香的重要值和生态位宽度最大,均能与其他物种发生生态位重叠。主要树种间整体上生态位重叠不明显(Oik为0~0.14),竞争不激烈,但生态位相似性指数较高(Cik为0.47~1.00),可能与喀斯特地区群落植被呈斑块状镶嵌分布有关。种间正联结与负联结基本持平或正联结种对稍占优势,种间关联度普遍较弱,种间关系未固定,群落处于不稳定演替后期,并向正联结的方向发展。 3.2.1 圆果化香群落生态位宽度特征 物种的生态位宽度越大,其对资源利用能力和竞争能力越强,在不同环境中具有较强的适应性,往往朝着泛化种方向发展[18]。通常重要值大的物种生态位宽度大,重要值小的物种重要值小,而有的物种重要值大小与生态位宽度排序并不一一对应[19],如荔波鹅耳枥重要值较大但生态位宽度却相对较低,而光叶海桐的重要值和生态位宽度排序一致。生态位宽度既与重要值大小有关,还与其频度、生境异质性有重要联系[20]。群落中物种的BL和BS指数在排序结果上基本一致,仅清香木、粗糠柴、亮叶猴耳环和荔波鹅耳枥略有变化。圆果化香重要值和生态位宽度均居首位,为喜光树种,也是优势种和建群种之一,光补偿点和饱和点高,耐旱及耐贫瘠,其芽、枝、叶等器官拓展生存空间,提高光合面积,且根系发达,穿透力强,从而提高对恶劣环境的生存适应和竞争能力[17]。 3.2.2 圆果化香群落生态位重叠特征 圆果化香与其他主要物种均发生了生态位重叠(0.05≤Oik≤0.06),表明生态位宽度较大的种群,与其他种群产生重叠的可能性较大。但整个群落的Oik集中分布在0~0.20,处于较低水平,喀斯特生境异质性,物种通过生态位分化来减弱种间竞争[20]。生态位宽度高的物种,其生态位重叠值指数也相对较高,两者呈正相关关系[5]。而本研究中物种的生态位宽度与生态位重叠值之间并没有表现出高度一致性。程中秋等[21]认为生态位宽度较大的阿尔泰狗娃花(Heteropappusaltaicus)、丝叶山苦荬(Ixerischinensis)和叉枝鸦葱(Scorzoneradivaricata)与其他物种的重叠指数并不高,表明生态位宽度和生态位重叠指数存在不一致的情况。马晓迪等[20]研究台湾相思(Acaciaconfusa)群落同样发现生态位宽度较大的种群存在较小的生态位重叠。植被在演替进程中对环境资源利用分化严重,资源呈现高度的空间异质性。 3.2.3 圆果化香群落生态位相似性特征 整体上群落的生态位相似性程度较高,群落不稳定。物种生态位相似性与其生态位宽度呈正相关[1],但也有生态位宽度较小的物种之间生态位相似性较高[19]。如生态位宽度大的圆果化香与其他物种的相似性不高,而生态位宽度较小的马尾松与刺叶桂樱的生态位相似性却很大,由于群落空间格局的不均匀性、复杂性导致部分物种向适宜生境斑块集中,以及物种间的生物学特性不同,最终导致这些物种的生态位宽度较小,而生态位相似性却较高[22]。生态位相似性较高的物种间其生态位重叠指数相对较高[2],如香叶树与小果柿、枫香树与马尾松的生态位重叠值大,生态位相似性也较高。而亮叶猴耳环与枫香树、马尾松、枫香树与刺叶桂樱等生态位相似性大,但生态位重叠值小。原因与生态位宽度类似,可能是生境的理化性质和物种的生态习性差异所致。 3.2.4 圆果化香群落种间联结性特征 演替中的群落种间关系逐渐以正相关为主,来确保物种间的稳定共存,因而演替至后期总体上表现为正联结[23]。圆果化香群落种间整体上呈不显著的负关联,总体联结性较为松散,处于不稳定状态。然X2检验和AC显示,正负联结种对数量上基本持平或正相关种对略占优势,预测圆果化香群落正向不稳定正联结的方向发展。与前人对黔中喀斯特次生林群落演替趋势向正向演替发展的判断一致[17]。群落不稳定,物种趋于独立分布的主要原因可能是地面岩石裸露较大,物种分布不连续以及自身生态习性差异所致。3 结论与讨论
3.1 结论
3.2 讨论
