PagAPY1基因调控银腺杨耐旱性的作用机制研究
2023-11-29王质璞李卓蓉罗志斌邓澍荣
王质璞,李卓蓉,罗志斌,邓澍荣
(林木遗传育种全国重点实验室,国家林业和草原局森林培育重点实验室, 中国林业科学研究院林业研究所,北京 100091)
干旱是制约植物生长的主要非生物胁迫。在我国,干旱和半干旱土地面积占全国土地总面积的45%以上[1]。随着全球气候变化加剧,干旱对森林等生态系统的威胁将愈发频繁和严重[2]。植物对干旱的适应性包括增加根系深度和生物量,减少叶片水分散失,激活脱落酸(ABA)等逆境响应激素信号,提高渗透调节物含量等[3-4]。相比模式草本植物和农作物,目前对林木适应干旱的分子调控机制还研究不足。解析重要抗旱基因功能,揭示林木抗旱机制,对利用基因工程技术开展林木改良,提高森林生态功能和生产力具有重大意义[5]。
核苷三磷酸双磷酸水解酶(nucleoside triphosphate diphosphohydrolase,NTPDase),也称Apyrase(APY),属于E型ATPase家族,其底物为多种核苷三磷酸或双磷酸(NTP/NDP),通过水解NTP/NDP的γ-和β-磷酸酐键,生成单磷酸核苷(NMP)和磷酸基团[6]。Apyrase蛋白一般包括5个保守结构域(apyrase conserved region,ACR),其中ACR1—4在植物中相对保守。APY在植物生长发育和逆境适应等方面发挥重要功能[7-8]。在拟南芥(Arabidopsisthaliana)中,AtAPY1和AtAPY2是调节生长发育的关键基因,过表达AtAPY1或AtAPY2促进植株生长,而双突变体生长受严重抑制[9]。同时,抑制AtAPY1/2基因表达阻碍了气孔闭合[10]。在豌豆(Pisumsativum)中过表达PsAPY1改善了根系构型和植株抗旱性[8]。胡杨PeAPY2通过促进囊泡运输提高植物低温耐性[11]。目前对Apyrase抗逆相关功能研究主要集中在草本植物,或通过异源表达验证功能,林木自身Apyrase的功能鉴定及其调控抗逆性的机制研究尚不充分。
早期研究认为,植物Apyrase主要位于质膜或质外体中,调控胞外ATP(extracellular ATP, eATP)水平[12]。eATP是一类重要的信号分子,它通过细胞主动分泌或质膜受损后被动扩散等方式释放到胞外,与位于质膜上的嘌呤受体相结合,激活下游的胞质Ca2+、H2O2等信号,调控植物生长或诱导防御反应[13-15]。eATP的生理作用具有双重性,较低浓度eATP促进生长,而高浓度(毫摩尔范围)eATP抑制生长,并可引发细胞程序性死亡[16-17]。此外,eATP对气孔运动具有调节作用,施加低浓度的eATP促进气孔开放,而高浓度eATP促进气孔闭合[18]。在植物生长发育和环境适应过程中,eATP生理功能的发挥取决于Apyrase等酶的精确调控。
然而,近期证据表明AtAPY1和AtAPY2主要位于高尔基体,而较少分布于细胞质膜或质外体[19]。通过分析微粒体中的水解酶活性,发现AtAPY1/2主要以UDP和GDP为底物,水解UDP、GDP等核苷二磷酸,产生NMP,从而调控高尔基体内的糖基化过程[20-22]。但同时推测AtAPY1/2也可能通过水解高尔基体内或分泌囊泡中的ATP,调节eATP水平[23-24]。目前,林木中主要Apyrase基因的细胞定位和生理功能还有待进一步明确。
银腺杨(Populusalba×P.glandulosa)生长速度快、环境适应性强,是我国北方重要的造林树种,被广泛应用于防风防沙、水土保持以及用材造纸等领域[25]。本研究利用生物信息学方法鉴定出响应干旱胁迫的Apyrase成员PagAPY1,获得了过表达PagAPY1转基因杨树,分析该基因调控杨树抗旱性的生理功能,为阐明林木Apy基因调控干旱适应性的分子机制提供理论基础。
1 材料与方法
1.1 实验材料
银腺杨来源于中国林业科学研究院。银腺杨组培苗首先在1/2 MS培养基上扩繁,培养条件为(25±1) ℃、16 h光照/8 h黑暗,光照强度125 μmol/ (m2·s)。培养1个月后移栽至10 cm×10 cm塑料盆中,基质为营养土、蛭石、珍珠岩体积比5∶1∶1, 培养条件为25 ℃/18 ℃、16 h光照/8 h黑暗,相对湿度约60%,2个月后选取长势一致的植株用于干旱处理。
1.2 基因序列分析
以拟南芥AtAPY1序列为索引通过BLAST程序搜索银腺杨基因组(https://www.ncbi.nlm.nih.gov/datasets/taxonomy/153471/)中的同源基因,获得PagAPY1全长序列,利用ClustalX进行氨基酸多重序列比对。
1.3 载体构建与遗传转化
利用CTAB法提取银腺杨幼苗总RNA,使用PrimescriptTM1st strand cDNA synthesis kit(Invitrogen)合成cDNA,利用基因特异性引物PagAPY1-F/R(表1)扩增PagAPY1全长编码区,通过同源重组技术将PagAPY1克隆至Gateway入门载体pDONR207,随后又通过LR反应分别重组至亚细胞定位载体pK7WGF和植物过表达载体pK7WG2D。分离拟南芥叶片原生质体并利用PEG法共转化高尔基体marker(Golgi-CFP)与重组质粒pK7WGF-PagAPY1[26],使用激光共聚焦显微镜(ZEISS LSM880,德国)观察目的蛋白的亚细胞定位。将pK7WG2D-PagAPY1转化农杆菌GV3101,利用叶盘法[27]侵染野生型银腺杨,利用卡那霉素(50 mg/L)筛选阳性转基因植株。

表1 试验中所用的引物序列
1.4 基因表达量分析
利用CTAB法提取对照或干旱处理下野生型(WT)/过表达(OE)杨树叶片总RNA,使用Takara公司(中国)TB Green©Premix Ex TaqTM试剂盒进行反转录和荧光定量PCR(qRT-PCR),每个样品3次技术重复,选取PagActin2/7作为内参基因,利用2-ΔΔCT法计算相对表达量,PagAPY1及其他干旱响应基因的特异性引物序列见表1。
1.5 干旱处理及抗旱生理指标分析
野生型和转基因杨树土培2个月后用于干旱处理,首先每天浇水50 mL以保持土壤水分,7 d后停止浇水开始干旱处理,干旱时长为14 d,对照组保持每天浇水50 mL。分别在干旱后第7天和第14天时拍照观察表型,干旱处理后恢复浇水3 d,统计野生型和转基因杨树的存活率、株高、根长和根干质量等指标。另外分别在干旱0、24、48 和72 h对野生型杨树叶片取样用于PagAPY1表达量分析。
选取正常浇水时野生型和转基因植株测定离体叶片失水率。叶片离体后立即称质量,记为0 min质量,随后分别在10、20、30、40、60、90以及150 min时称质量。叶片失水率计算为每个时间点质量的减少量与0 min质量的比值,每个基因型3棵植株,每棵植株4个样本重复。
选取干旱处理0、24 和48 h各杨树株系功能叶测定气孔开度。用镊子撕取叶片下表皮制作临时玻片,在20倍光镜下观察气孔并拍照,计算气孔开度。另外测定ABA处理对各株系气孔开度的影响,选取正常生长良好的野生型及转基因植株功能叶,在荧光灯下光诱导3 h,随后用0、10、30 μmol/L ABA溶液处理2 h,制作临时玻片观察气孔并计算其开度。
选取生长良好且状况一致的野生型及转基因杨树无菌苗进行PEG6000处理,分别接种至含有0%、2%和3%(质量分数)的PEG6000的继代培养基中培养,生长28 d后测量根长。
1.6 数据处理
利用Excel和SPSS 20.0软件处理数据,使用多重比较(Duncan)进行显著性分析。
2 结果与分析
2.1 银腺杨PagAPY1基因鉴定及表达量分析
利用拟南芥AtAPY1序列在银腺杨基因组中搜索Apy同源基因,筛选得到PagAPY1全长基因。氨基酸序列比对表明,PagAPY1与拟南芥AtAPY1、豌豆PsAPY1等高度同源,均含有Apyrase保守结构域,特别是ACR1—4相似度较高(图1a)。位于ACR1中的天冬氨酸D71、丝氨酸S74,ACR2中的苏氨酸T147、精氨酸R151,ACR3中的谷氨酸E196,ACR4中的天冬氨酸D223是决定Apyrase催化活性的保守氨基酸[6]。将PagAPY1与YFP荧光蛋白融合,瞬时转化拟南芥原生质体,显微观察发现PagAPY1与高尔基体marker(CFP)具有严格的共定位,表明PagAPY1定位于高尔基体中(图1b)。在干旱胁迫24 h时,PagAPY1基因表达显著升高,相比0 h提高了约4倍;干旱处理48 h时,PagAPY1表达受诱导的趋势回落至对照的1.75倍左右,表明PagAPY1表达在短期内受到干旱胁迫诱导(图1c),推测PagAPY1可能在银腺杨响应干旱胁迫中起作用。
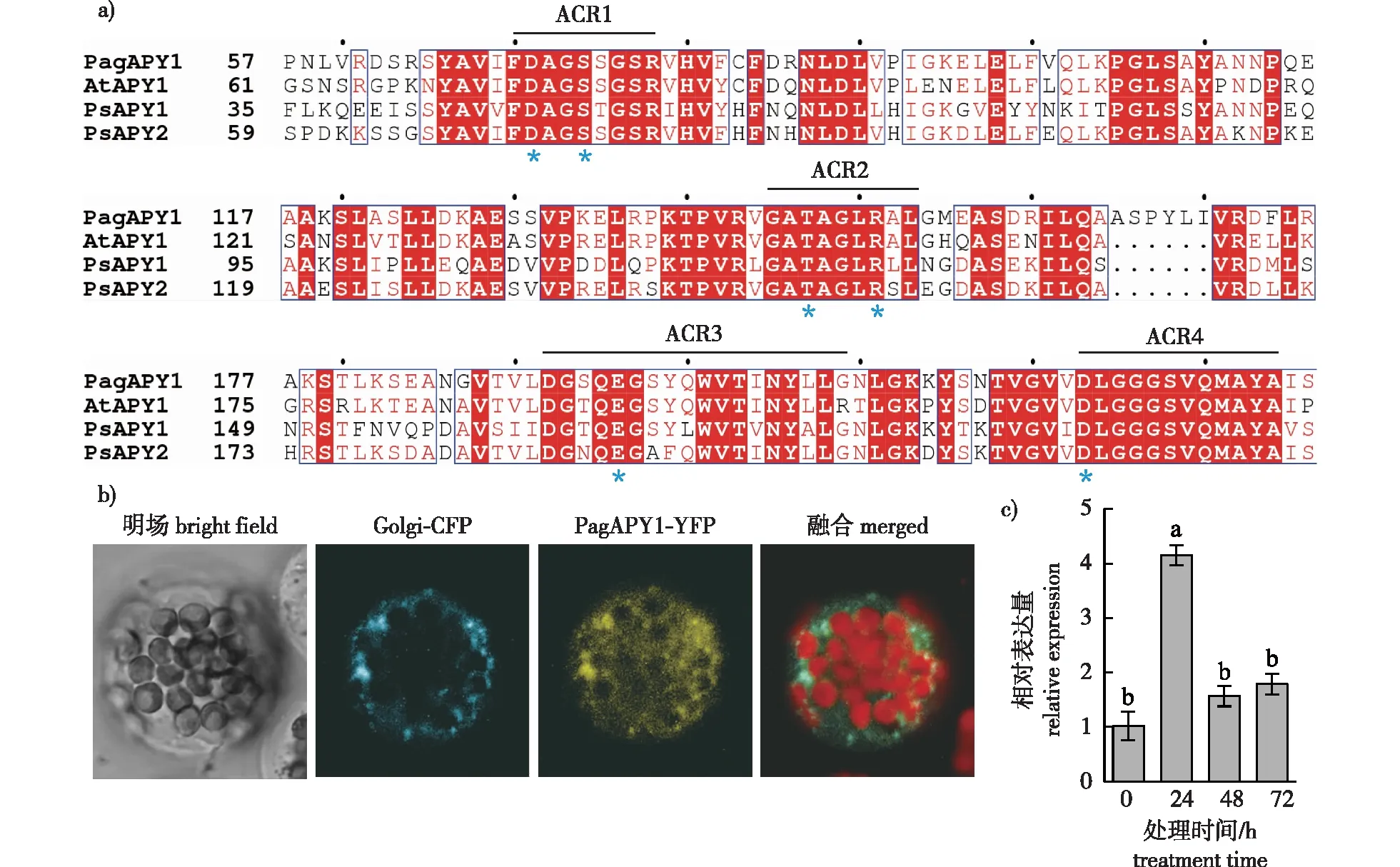
a)银腺杨APY1与拟南芥、豌豆同源蛋白的多重序列比对 multiple sequence alignment of PagAPY1 with homologues in Arabidopsis and pea. 其中:At.拟南芥Arabidopsis;Ps. 豌豆Pisum sativum;蓝色星号标记为保守氨基酸残基blue asterisks indicate conserved amino acid residues;b) PagAPY1亚细胞定位 subcellular localization of PagAPY1;c) 干旱处理后PagAPY1表达量的变化。不同字母表示差异显著,P<0.05。下同。Gene expression level of PagAPY1 in response to drought. Different letters indicate significant differences, P<0.05. The same below.图1 多重序列比对、亚细胞定位与基因表达对干旱的响应Fig. 1 Amino acid alignment, protein subcellular localization and drought-induced expression of PagAPY1
2.2 PagAPY1过表达杨树株系的获得
为了鉴定PagAPY1在杨树抗旱过程中的功能,利用农杆菌介导的遗传转化获得了转基因银腺杨植株。采用qRT-PCR对8个PCR鉴定为阳性的株系进行表达量分析,结果见图2。

图2 转基因杨树中PagAPY1表达量检测Fig. 2 Expression level of PagAPY1 in transgenic poplars
OE1—8转基因株系中PagAPY1的表达量均显著高于野生型银腺杨,其中OE1、OE2和OE3株系的表达量上调最为显著。因此,选取OE1—3株系进行后续试验。
2.3 PagAPY1提高转基因杨树抗旱性
干旱处理7 d,野生型银腺杨出现了较为严重的萎蔫现象,表明此时受到干旱胁迫的严重伤害(图3a)。与野生型相比,PagAPY1过表达株系叶片只出现了轻微萎蔫。在干旱处理14 d时,野生型和转基因植株都出现了明显萎蔫,野生型相较于转基因植株的萎蔫程度更为严重,叶片已经干枯,而转基因株系叶片干枯现象不明显。复浇水3 d后,大部分野生型植株已经死亡,而转基因植株相比野生型恢复程度更好。如图3b所示,复水后野生型植株存活率为25%,而转基因株系的存活率均大于50%,尤其是OE1株系的存活率高达75%。同时,干旱条件下过表达株系的株高显著大于野生型(图3c)。干旱条件下,转基因植株的平均根长增加了20.9%~29.6%,根干质量比野生型增加了22.6%~38.1%(图3d和3e)。以上结果表明,过表达PagAPY1通过促进根系生长提高转基因植株的耐旱性。

图3 干旱胁迫对野生型和转基因杨树生长的影响Fig. 3 Effects of drought stress on the growth of wild-type and transgenic poplars
2.4 渗透胁迫对不定根生长的影响
进一步研究渗透胁迫下PagAPY1对根系发育的影响发现,2%和3%的PEG6000处理几乎完全抑制了野生型银腺杨不定根生长(图4a和4b),与野生型相比,过表达株系不定根生长受PEG抑制程度显著减小,在2%的PEG6000处理下降低了13.47%~26.93%,在3%的PEG6000处理下降低了33.02%~46.71%。可见,渗透胁迫下过表达PagAPY1促进了根系生长。

。图4 高渗处理对不定根生长的影响Fig. 4 Effects of osmotic stress on adventitious root growth
2.5 干旱胁迫下过表达PagAPY1减少叶片水分散失
为了进一步明确PagAPY1提高杨树抗旱性的作用,测定了离体叶片的失水率(图5a)。结果表明,过表达杨树株系的叶片失水率显著低于野生型。在150 min时,野生型叶片失水率达到38%,而转基因株系为30%,说明PagAPY1提高了叶片的保水能力。与0 h相比,在干旱处理24 h后,野生型杨树的气孔开度逐渐降低,尤其在48 h时降低程度显著(图5b)。与野生型相比,转基因杨树在干旱24 h时气孔开度即发生显著降低,48 h后仍明显低于野生型(图5b和5c)。进一步ABA处理试验显示,在10 μmol/L ABA的处理下野生型植株气孔开度变化不显著,在30 μmol/L ABA处理后才表现出轻微闭合,与野生型相比,过表达植株叶片的气孔开度在10 μmol/L ABA的处理下显著降低,在30 μmol/L ABA处理后降低幅度更大。

a) 叶片失水率leaf water loss rate;b) 干旱处理下气孔的显微观察microscopic observation of stomata under drought stress;c) 干旱处理下的气孔开度分析stomatal aperture under drought stress;d) 不同浓度ABA处理下的气孔开度分析stomatal aperture under ABA treatments。图5 干旱和ABA处理对叶片水分保持的影响Fig. 5 Effects of drought and ABA treatment on leaf water retention capacity
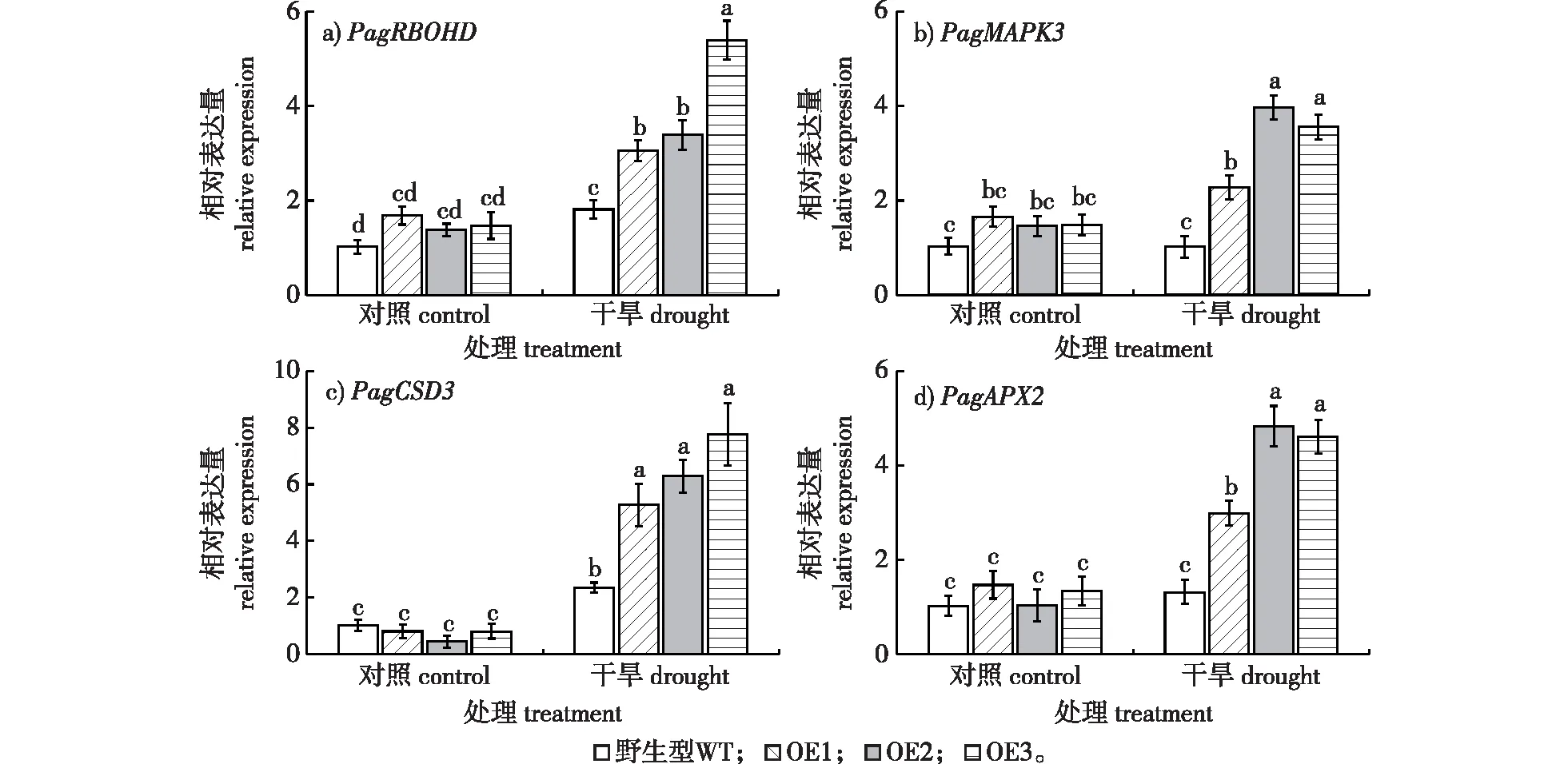
图6 干旱处理下抗逆相关基因的表达Fig. 6 Expression level of stress-related genes under drought
2.6 抗旱相关基因表达
为了进一步探究PagAPY1调控杨树抗旱性的机制,利用qRT-PCR技术检测抗逆相关基因表达。在干旱处理48 h后,过表达植株中NADPH氧化酶RBOHD、促分裂原活化蛋白激酶MAPK3、铜锌超氧化物歧化酶CSD3和抗坏血酸过氧化物酶APX2基因表达相比野生型显著上调(图6),表明干旱条件下过表达PagAPY1促进了活性氧信号和抗氧化系统。
3 讨 论
3.1 PagAPY1的分子功能
Apyrase的亚细胞定位决定其生理功能的发挥。已有研究表明,拟南芥AtAPY1和AtAPY2主要位于高尔基体中,以UDP为主要底物[20],而生理和免疫学证据表明AtAPY1/2参与调控了eATP水平[9]。胡杨(P.euphratica)PeAPY2在质膜、高尔基体和内质网中均有分布,主要通过细胞分泌途径调控eATP信号活性[11]。通过亚细胞定位分析,发现PagAPY1位于高尔基体中。氨基酸序列分析表明,PagAPY1具有保守的ACR结构域,并具有决定催化NTP/NDP水解活性的保守氨基酸残基,推测PagAPY1位于高尔基体中,调控高尔基体内或分泌囊泡中的NTP/NDP水平。
3.2 干旱胁迫下PagAPY1促进杨树根系生长
通过促进根系生长以增加水分吸收是植物适应干旱胁迫的重要策略[3]。在干旱条件下,PagAPY1过表达株系根长以及根系生物量相比野生型显著提高。此外,在PEG6000高渗处理时,过表达PagAPY1促进了不定根生长。这些结果表明干旱胁迫下PagAPY1通过促进根的生长,提高根系水分获取,从而增加植株抗旱性。在拟南芥atapy1apy2双突变体中,Apyrases活性降低阻碍了根部生长素极性运输,抑制了根系生长[28]。过表达豌豆PsAPY1后根系显著增加,同时提高了转基因植株的耐旱性[8]。由于同源蛋白AtAPY1定位于高尔基体,通过水解高尔基体内腔中的NDP(如UDP、GDP)调控蛋白糖基化修饰过程[20],PagAPY1也可能通过水解高尔基体中的UDP或GDP等,促进生长素运输蛋白如ABCB(ATP-BINDING CASSETTE subfamily B transporters)转运蛋白和PIN(PIN-FORMED)蛋白等的糖基化修饰[8],维持生长素的运输活性,促进根系生长。
3.3 PagAPY1提高杨树叶片保水能力
PagAPY1过表达植株离体叶片的失水率比野生型显著降低,说明PagAPY1降低了叶片的失水率。植物体内水分主要通过叶片气孔散失。在干旱条件下,过表达植株比野生型气孔闭合程度更高。ABA是干旱胁迫下促进气孔闭合的关键激素,过表达株系对ABA诱导的气孔关闭更为敏感。由此推测,PagAPY1通过促进ABA诱导的气孔关闭,促进干旱胁迫下气孔闭合,以减少水分散失。近期研究表明,在一定浓度范围内,eATP对气孔开放有促进作用,而过表达Apyrase抑制了eATP对气孔开放的促进作用。Atapy1/atapy2突变体对ABA诱导的气孔关闭不敏感,而在拟南芥中过表达Apyrase促进了ABA诱导的气孔关闭。由于PagAPY1位于高尔基体中,其可能通过水解分泌囊泡中的ATP,减少分泌到细胞外的ATP数量,降低eATP水平,从而抑制了eATP介导的气孔开放。此外,PagAPY1在高尔基体中对蛋白糖基化的调控作用是否影响了ABA诱导的气孔关闭,尚未可知,有待进一步研究。
3.4 PagAPY1提高了抗逆相关基因表达水平
干旱胁迫下,PagAPY1过表达杨树中RBOHD和MAPK3基因表达显著上调。NADPH氧化酶是产生活性氧信号的关键酶,参与对非生物胁迫的响应。近年来,NADPH氧化酶被鉴定可被eATP信号激活,产生活性氧和下游Ca2+信号,从而传递eATP介导的防御信号[29-30]。类似地,MAPK3也参与传递了eATP信号[31]。在干旱条件下,MAPK3通过保护膜系统免受氧化胁迫伤害,同时促进其他胁迫相关基因表达以提高抗旱性[32]。因此推测PagAPY1可能通过调节eATP水平,维持胞外ATP稳态,增强转基因植株对干旱胁迫的防御反应。相一致地,过表达杨树中抗氧化酶基因CSD3和APX2在干旱胁迫下显著上调,说明PagAPY1通过促进活性氧信号,增强了抗氧化酶系统活性。
总之,在干旱胁迫下,PagAPY1通过维持蛋白糖基化和eATP稳态平衡,促进生长素极性运输,提高根系生长;通过调控叶片中eATP含量,促进气孔闭合,减少叶片水分散失;同时,激活活性氧信号,增加抗氧化酶系统活性,从而提高了杨树抗旱性。
