金佛山方竹果实发育特征及淀粉粒动态变化
2023-11-29伍虹雨林树燕丁雨龙
伍虹雨,林树燕,丁雨龙,张 玉,杨 露, 秦 敏,蔡 鸥
(1. 南京林业大学,南方现代林业协同创新中心,南京林业大学竹类研究院,江苏 南京 210037; 2. 南京林业大学生命科学学院,江苏 南京 210037)
金佛山方竹(Chimonobambusautilis)是我国西南地区特有竹种,主要分布在海拔1 200~2 200 m的重庆南川区、贵州省桐梓县、正安县和绥阳县以及云南省镇雄等地[1],竹笋味美可口营养价值高[2],种子也含有丰富的营养物质,是我国西南高山地区极具特色的笋用竹种;除笋用外,在贵州省桐梓县原生金佛山方竹林面积达28 000 hm2,在生态景观方面也有独特价值。由于竹类植物年龄的不确定性,金佛山方竹母竹移栽造林可能导致成片开花造成竹子死亡的潜在危险。因此,相关学者进行了大规模金佛山方竹实生苗育苗研究,同时有关金佛山方竹种子萌发、不同种源之间实生苗差异,以及实生苗林与天然林发笋规律差异等研究也逐步展开[1,3]。
种子的发育是否正常是植物自然更新能力的影响因素之一。由于禾本科植物如小麦(Triticumaestivum)、水稻(Oryzasativa)的果实类型为颖果,颖果果皮不开裂且与种皮高度愈合,难以分离,因此常称为种子,一般由胚、胚乳和种皮构成。竹类植物果实根据果皮的性状特征将果实分为基本型颖果、浆果状颖果、坚果状颖果、半坚果状颖果以及两型果[4],其中浆果状颖果果实肉质,果皮较厚,含水量较高[5]。金佛山方竹作为禾本科植物,花期为每年8月至次年2—3月[2],3月上旬至4月下旬为果期。果实成熟后椭圆形,含水量高,为浆果状颖果,不耐储藏。竹类植物营养生长周期长,不易开花、花期短以及结实率低的特点导致有关竹类植物胚、胚乳的研究比较少[6],前期研究多集中在基本型颖果的发育过程,而对于浆果状颖果的发育过程还不甚了解。淀粉作为竹子种子内部贮存的主要营养成分,在种子发育的整个过程中淀粉粒的动态积累情况有无变化规律,此方面的研究很少。近几年,重庆、贵州大娄山山脉分布的金佛山方竹出现零星开花,这为研究花后果实的发育提供了珍贵的材料。笔者详细研究了金佛山方竹花后胚、胚乳和果皮的发育过程,并观察了果实发育过程中淀粉粒的动态变化,为理解方竹果实的发育过程提供精准可靠的依据,也为进一步完善竹类植物的生殖生物学机理提供基础资料。
1 材料与方法
1.1 试验材料
2019年3月2日至4月25日,于金佛山方竹盛花期结束至果实成熟期间,每隔1 d固定50个自然传粉后的子房,直至果实成熟,样品放入盛有甲醛-乙酸-乙醇固定液(FAA) (70%乙醇、甲醛、冰醋酸体积比为18∶1∶1)的玻璃瓶中抽气固定备用。果实采摘后,室内阴凉处继续放置7 d,每天固定20粒。
1.2 试验方法
果实发育特征观察:制片前,爱氏苏木精整染,后复水返蓝,再梯度脱水常规石蜡法制片,切片厚度为8 μm,番红-固绿着色,中性树脂胶封片。在显微镜(LEICA DM2500)下拍照观察,利用Photoshop进行显微图像处理。
淀粉粒变化观察:石蜡切片脱蜡后,经过各级酒精复水之后,移入0.5%(体积分数)高碘酸液中,经席夫试剂,漂洗液,脱水,固绿染色,透明,最后用中性树脂胶封固。在显微镜(LEICA DM 2500)下观察拍照。
扫描电镜观察:用蒸馏水将不同发育时期种子表面的FAA 进行冲洗,依次经过70%(体积分数,下同)→90%→100%的乙醇梯度脱水(2次)。临界点干燥仪(K850英国EMITECH公司)中干燥,样品粘台,离子溅射仪(E1010日本日立公司)喷金,Quanta 200环境扫描电镜(美国FEI有限责任公司)下进行拍照与观察。
2 结果与分析
2.1 金佛山方竹种子胚的发育
经观察分析可知,胚的发育过程分为原胚期、胚分化期、胚成熟期共3个时期(图1)。
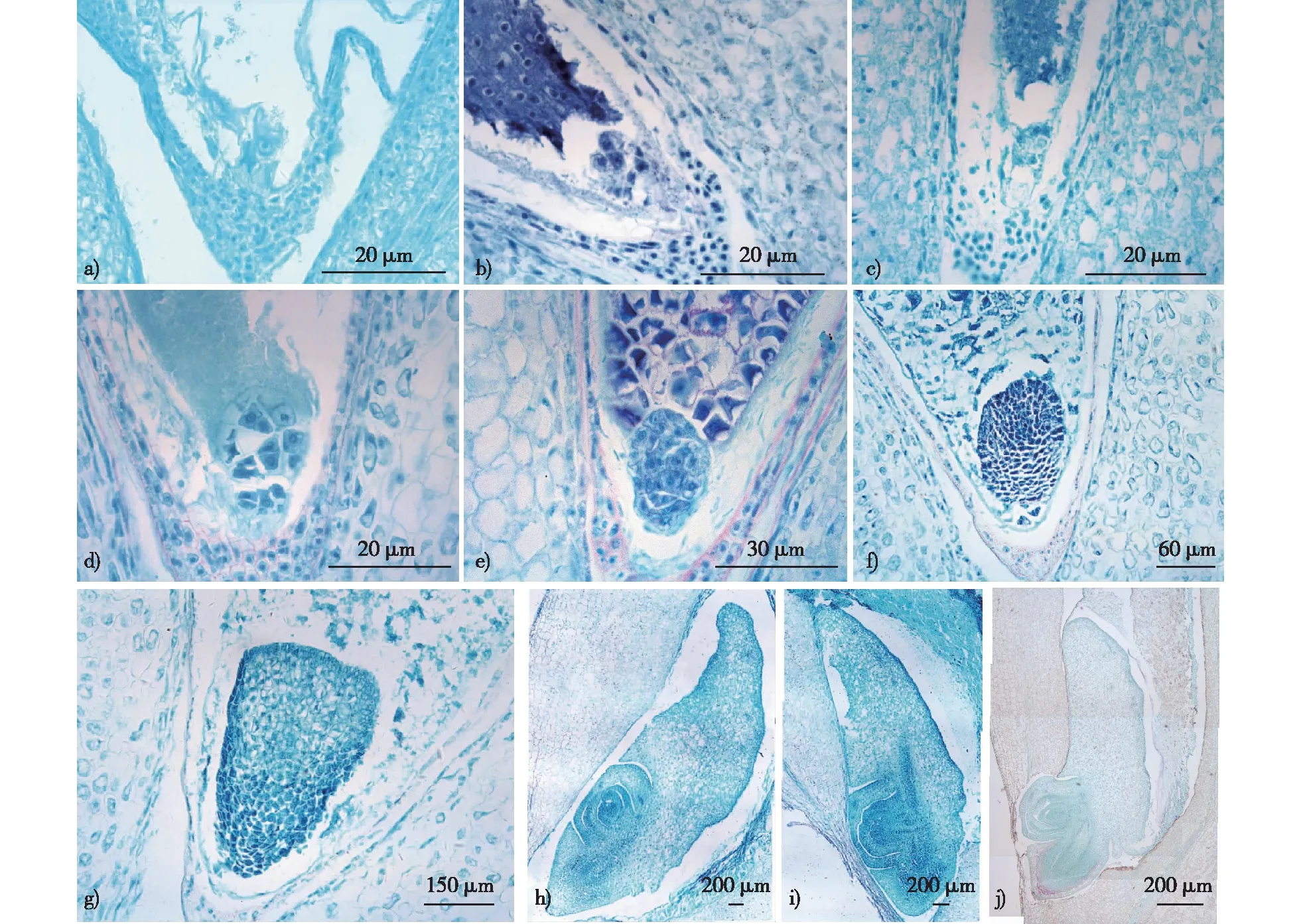
a.合子zygote;b和c.原胚期proembryo;d.多细胞原胚multicellular proembryo;e、f、g.梨形胚期pear shaped embryo;h.胚分化期differential embryo;i和j.成熟胚mature embryo。图1 金佛山方竹胚的形成过程Fig. 1 The embryo development of Chimobambusa utilis
金佛山方竹种子胚的结构由胚根、胚芽、盾片、胚轴、胚芽鞘、外胚叶和胚柄等部分组成(图1i)。受精后精卵细胞结合产生合子(图1a),当受精后的子房长度为3 mm时,合子进行第1次有丝分裂,产生二细胞原胚,靠近合点端的细胞为顶细胞,靠近珠孔端的细胞为基细胞,顶细胞横向分裂,形成四胞原胚,而基细胞暂停分裂,顶细胞继续向各方向分裂,形成一团细胞,此时子房长度为4~6 mm,胚为原胚期(图1b、1c)随后基细胞开始分裂,子房长度为7~9 mm,胚发育时期为梨形原胚(图1d)。胚继续纵向伸长,随着细胞不断分裂,胚逐渐增大,成为梨形胚(图1e、1f、1g)。
梨形胚之后将进行器官的分化,当子房长度为10~12 mm时,胚上部形成盾片(图1h),在盾片分化期,果实大小已不再变化,直至采摘,采摘后的果实要经过后熟作用,完成胚的进一步发育,为胚成熟期(图1i、1j)。
2.2 金佛山方竹种子胚乳的发育
胚乳是由精细胞和极核互相融合而形成的,首先形成初生胚乳核,之后胚乳核不断分裂。经观察分析,当子房长度为3 mm,合子进行第1次有丝分裂时,胚囊中已经形成了一定数量的胚乳核(图2a),此时位于合点端的反足细胞并未完全解体消失;金佛山方竹种子的胚乳发育类型为核型胚乳,游离核沿着胚囊壁分布(图2b、2c),此时子房长度为4~6 mm;在梨形胚时期,靠近胚方向的游离核先形成细胞壁,之后向合点端方向细胞化(图2d、2e),此时子房长度为7~9 mm;当子房长度为10~12 mm时,在梨形胚至胚分化初期,胚囊近珠孔端的细胞已完全细胞化(图2f、2g),此时果实长度基本稳定,不再继续增长,随着种子后熟,胚乳逐渐充满整个胚囊(图2h),糊粉层为胚乳的最外层细胞。

a、b和c.原胚期胚乳endosperm during proembryo period;d和e.梨形胚期胚乳endosperm during pear-shaped embryo period;f和g.梨形胚后期胚乳endosperm in late pear-shaped embryo stage;h.胚分化期胚乳endosperm cells in embryo differentiation stage。图2 金佛山方竹胚乳的形成过程Fig. 2 The endosperm development of C. utilis

a、b、c、d、e、f、g、h.采后第3、4、5、6、7、8、9、10天的果实内部形态the fruit morphology from the 3rd to the 10th day after collecting。图3 采摘后发生后熟Fig. 3 Post-ripening after C. utilis fruit collecting
在此阶段,种子从竹株上掉落,随后果实的胚、胚乳经过后熟作用至完全成熟,胚乳细胞充满整个胚囊,转为果实的贮藏组织。金佛山方竹的种子在采后3~5 d(图3a—3c),胚乳中部中空,采后的6~10 d(图3d—3h),胚乳细胞逐渐充满胚囊,种子完成后熟。受精后,胚和胚乳的发育进程不同步,胚乳的发育早于胚的发育,当胚乳发育到一定程度后,胚才会开始分裂和分化,当种子完成后熟时,胚乳和胚也已发育完全。
2.3 金佛山方竹种子果皮以及种皮的发育
经观察研究,金佛山方竹的成熟果实为浆果状颖果,由果皮、种皮、胚乳及胚组成。含水率高,果皮较厚,最外侧为一层表皮细胞,表皮细胞以内由多层薄壁细胞组成(图4)。
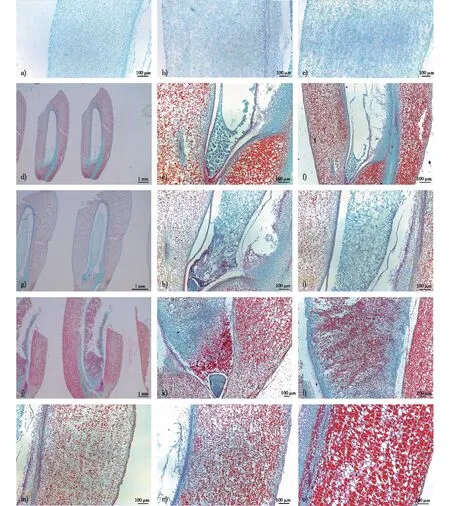
a.5 mm果实的果皮the pericarp when seed is 5 mm; b.10 mm果实的果皮the pericarp when seed is 10 mm; c.12 mm果实的果皮the pericarp when seed is 12 mm; d、e和f.原胚时期胚乳细胞淀粉粒分布starch granules in endosperm cells during proembryo period; g、h和i.梨形胚时期胚乳细胞淀粉粒分布starch granules in endosperm cells during pear-shaped embryo period; j、k和l.盾片分化时期胚乳细胞淀粉粒分布starch granules in endosperm cells during differential embryo period; m.原胚时期果皮淀粉粒starch granules in pericarp during proembryo period; n.梨形胚时期果皮淀粉粒starch granules in pericarp during pear-shaped embryo period; o.盾片分化时期果皮淀粉粒the starch in pericarp during embryo differentiation period。图4 金佛山方竹果皮的形成过程以及胚乳和果皮中淀粉粒动态变化Fig. 4 The formation process of pericarp and the dynamic changes of starch granules in endosperm and pericarp of C. utilis
果皮由基部向顶部逐渐增厚。随着果实的不断生长,果皮厚度不断增加(表1,图4a、4b、4c)。金佛山方竹的种皮由珠被发育而成,种皮在果皮内侧,细胞排列紧密,与果皮相连,不易分开,向内连接糊粉层。
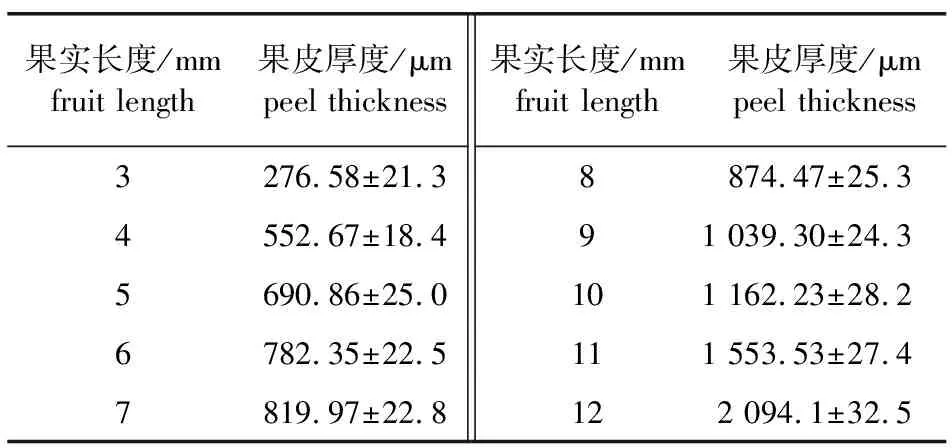
表1 金佛山方竹果实长度和果皮厚度对应表
2.4 金佛山方竹种子发育过程中淀粉粒变化
在胚乳细胞化前期,游离核中几乎观察不到淀粉粒。在原胚时期,靠近胚方向的胚乳细胞化,胚乳细胞中淀粉粒极少(图4d、4e、4f)。至梨形胚时期,靠近胚的胚乳细胞中出现少量的淀粉粒(图4g、4h、4i),直到盾片分化,胚乳中可见大量淀粉粒分布(图4j、4k、4l)。淀粉粒主要存在于果皮的薄壁细胞中,在果实发育的整个过程中,大量淀粉粒存在于果皮的薄壁细胞中(图4m、4n、4o)。
综上所述,淀粉粒在成熟胚乳细胞中大量分布,在胚乳细胞发育过程中呈现从少到多的变化趋势(图5)。在幼嫩果实胚乳薄壁细胞中,淀粉粒数量很少(图5a、5b),刚采摘下来的果实中,胚乳细胞中含有的淀粉粒较之前增多(图5c)。采摘后,种子进入后熟期,采后3~7 d,淀粉粒逐渐充满整个胚乳细胞(图5d—5h)。

a.9~10 mm果实胚乳细胞内淀粉粒starch grains morphology in endosperm when fruit long is 9-10 mm; b.11 mm果实胚乳细胞内淀粉粒starch grains morphology in endosperm when fruit long is 11 mm; c.12 mm果实胚乳细胞内淀粉粒starch grains morphology in endosperm when fruit long is 12 mm; d, e, f, g和h.果实采后第3、4、5、6、7天胚乳细胞内淀粉粒starch grains morphology in endosperm on the 3rd, 4th, 5th, 6th, 7th day after collecting。图5 金佛山方竹种子胚乳内淀粉粒扫描电镜观察Fig. 5 Starch granules in endosperm of C. utilis by scanning electron microscope
而淀粉粒在果皮发育整个过程中都存在,分布在果皮薄壁细胞内(图6)。在果实幼小期(3~6 mm),果皮细胞中淀粉粒开始累积(图6a、6b),而果实在中等大小直至大小不再变化时(7~12 mm),果皮细胞内淀粉粒数量增多(图6c、6d)。自采后直至第7天,果皮细胞内仍分布着大量淀粉粒(图6e—6i)。
淀粉粒在形态、大小上存在区别,胚乳中和果皮内淀粉粒形态包括圆形、椭圆形、多面体形。发育初期,淀粉粒比较小,多为圆形、椭圆形。发育后期,淀粉粒以多面体形居多。

a.3~4 mm果实中果皮淀粉粒starch grains morphology in pericarp when the fruit long is 3-4 mm; b.5~6 mm果实中果皮淀粉粒starch grains morphology in pericarp when the fruit long is 5-6 mm; c.7~9 mm果实中果皮淀粉粒starch grains morphology in pericarp when the fruit long is 7-9 mm; d.10~12 mm果实中果皮淀粉粒starch grains in pericarp when the fruit long is 10-12 mm; e, f, g, h, i.果实采后第3、4、5、6、7天果皮细胞内淀粉粒starch grains in pericarp on the 3rd, 4th, 5th, 6th, 7th day after collecting。图6 金佛山方竹果皮内淀粉粒扫描电镜观察Fig. 6 Starch granules in the pericarp of C. utilis by scanning electron microscope
3 讨 论
竹类植物属于禾本科竹亚科,果实类型亦为颖果,根据果实形态可分为基本型颖果、坚果状颖果、半坚果状颖果及浆果状颖果,其形态解剖结构可作为竹类植物分属的重要依据[7]。竹类植物如翠竹(Pleioblastuspygmaeus)、黄条金刚竹等属于基本型颖果[8-9],与小麦、水稻的果实解剖结构比较接近,果皮厚度较浆果状颖果薄[10]。郭起荣等[11]发现,毛竹果皮仅由内珠被内层和子房外壁3~4层细胞组成。竹类植物种子成熟时,浆果状颖果有较高的含水率,种子软而不耐贮藏,基本型颖果有较低的含水率,种子硬而较耐贮藏。这两种类型,果皮与种皮均相连不易分离。但金佛山方竹果实胚乳易物理剥离,而异叶苦竹(Arundinaviasimoniif.heterophylla)等基本型颖果果实的胚乳与种皮及果皮不易物理剥离[6]。
竹类植物如异叶苦竹[12]、翠竹[13]等雌蕊胚囊成熟后反足细胞表现为数量多以及存留时间长的特点,合子期仍可见大量反足细胞。原胚时期反足细胞才逐渐解体,与本研究金佛山方竹观察结果一致。其他禾本科植物如小麦胚胎发育初期,胚囊合点端的反足细胞从外向内逐渐衰退[14],水稻反足细胞在胚囊形成后开始增殖,并且发展为多细胞团,存留时间很久[15]。反足细胞在受精后增殖,细胞内多核仁和大液泡,并从合点端移向边缘,在反足细胞解体消失的地方胚乳游离核增多,其发生的原因可能与反足细胞的功能有关系[16]。
有些植物如大麦(Hordeumvulgare)[17]、龙须草(Eulalcopsisbinata)[18]等种子胚乳发育过程中,在游离核时期和细胞化时期胚乳细胞核核仁大多数为1个,只有极少数有2~3个。而玉米(Zeamays)胚乳细胞分裂增殖时,处在细胞分裂间期的游离核内含1个至数个核仁[19]。华山新麦草(Psathyrostachyshuashahica)胚乳形成过程中,核仁大部分为多数,有少数为1个[20]。
小麦的合子及原胚期的发育主要是依赖卵细胞来提供营养[21]。在多细胞原胚期,胚乳游离核的分裂和游离核形成细胞壁时,需珠心细胞和反足细胞提供营养。梨形胚时期,胚乳细胞淀粉粒增多,其主要靠胚柄对珠心细胞贮藏物质的吸收和运输提供营养。到盾片分化期时,主要由胚乳细胞来提供营养,胚柄仅提供少量的营养。胚乳细胞中淀粉粒丰富,当种子萌发时,淀粉粒转为胚继续生长的能量。玉米原胚期时,游离胚乳核中许多小球形淀粉粒处于不断积累同时又不断被利用的状态[22],是碳水化合物的暂时性贮藏形式。青稞(H.vulg)胚乳中的淀粉粒在进入发育的高峰期时,形态上发生很大变化,部分淀粉粒从圆形变成多边形、扁圆形等。谷物类颖果果皮中都有淀粉粒的积累,如小麦[23]、高粱(Sorghumbicdor)[24]、水稻[25]等。
种子后熟是指种子从母体上自然脱落后,内部需经过一系列的生理变化,种子才能达到完全成熟[26]。后熟可分为形态后熟、生理后熟,或二者并存[27]。也有研究认为种子后熟是休眠的一种,有生理休眠、形态休眠、形态生理休眠等[28]。种子后熟现象与种胚发育程度密切相关。银杏种子(Ginkgobiloba)后熟过程实质上是种胚不断分化生长的过程[29]。竹类植物种子如颖果一般没有后熟期,只要有适宜发芽的温度、水分、空气条件即可发芽[30],而有些浆果状颖果类型如筇竹(Chimonobambusatumidissinoda)种子[31]采摘后需经过10~20 d后可完全成熟。
金佛山方竹胚囊双受精后反足细胞数量较多、存留时间长。金佛山方竹种子胚和胚乳发育过程不同步,胚乳发育早于胚。种胚中盾片体积大,胚芽与胚根的夹角随着种子的生长发育由近似直角转变为锐角。金佛山方竹果实为浆果状颖果,绿色,不具腹沟,果皮较厚,肉质,含水量高。果实生长发育过程中,果皮薄壁细胞不断增加并加厚,种皮在果皮内侧,当果实成熟时种皮细胞解体,仅留残迹,糊粉层细胞位于胚乳细胞的最外侧1~2层。金佛山方竹果实存在后熟现象,采摘后14 d左右胚和胚乳才能分化成熟。金佛山方竹游离胚乳核中极少有淀粉粒的存在,只在成熟胚乳细胞中大量存在,而果皮薄壁组织在合子期到果实成熟期间一直存在淀粉粒。
