温度对白皮松种子萌发过程中储藏物质代谢及酶活性的影响
2023-11-29郭聪聪沈永宝史锋厚
郭聪聪,沈永宝,史锋厚
(1. 江苏农林职业技术学院风景园林学院,江苏 句容 212400;2. 南京林业大学风景园林学院,江苏 南京 210037; 3. 南京林业大学林草学院,国家林业和草原局南方林木种子检验中心,江苏 南京 210037)
种子萌发以胚从其包被组织(种皮、胚乳)中伸出为这一生理过程的完成。种子代谢和细胞活动需要在吸水或吸胀作用下重新启动,随后引起胚的膨大和胚根(或其他器官)突破包被组织。呼吸代谢在种子吸胀后随即被激活,从而为萌发过程的一系列事件提供代谢能量。种子开始萌发时,贮藏组织中营养物质代谢开始启动,为幼苗的生长和形态建成提供营养[1]。
温度是种子萌发所需的重要环境因素,由于所处的生长环境和生态特征不同,每种植物种子都需要在一个特征性的限定温度范围内萌发。温度可直接作用于吸水和生理代谢反应来调控种子萌发过程,从而影响种子的发芽率和萌发速率[2]。温度过高或过低都会影响种子内部的生理代谢活动,如改变酶的活性、物质代谢和呼吸作用异常甚至产生某些有害物质,从而影响种子萌发与幼苗生长[3]。在高温下,豇豆(Vignaunguiculata)种子中呼吸高峰和α-淀粉酶活性高峰提前,蛋白质分解速度加快,而低温则导致萌发不启动[4]。在一定温度范围内,薄壳山核桃(Caryaillinoinensis)种子中淀粉酶活性随温度升高而提高,温度通过改变淀粉酶活性来影响糖代谢进而调控种子萌发[5]。穿心莲(Andrographispaniculata)种子适温下萌发时储藏物质的转化利用率要显著高于高温或低温下的物质代谢强度[6]。温度是限制玉米(Zeamays)种子萌发的主要因素,低温环境影响了贮藏物质的转运,从而阻碍种子萌发,抑制幼苗生长[7]。同样,低温降低了银杏(Ginkgobiloba)种子的呼吸速率,抑制了蛋白质和淀粉的水解,可溶性糖和游离氨基酸被用于种子萌发过程中的物质和能量代谢[8]。
白皮松(Pinusbungeana)是中国特有树种,分布于山西、陕西、河南、四川、甘肃、湖北及湖南北部[9]。其树姿优美、树皮奇特,观赏价值高,在城市园林绿化中被广泛应用,生产上以种子繁殖为主。前人关于白皮松种子萌发的研究主要侧重于休眠原因方面,如种皮结构与透水性、胚发育情况与离体培养以及种皮抑制物种类与含量等[10-13],萌发温度通常选择25 ℃,但该温度下种子发芽率较低,认为种子萌发所需的外部条件尤其是温度并未得到满足。因此,本研究进行不同温度下的萌发试验以探索在高温环境中白皮松种子是否产生了热抑制或者热休眠现象,并对适宜温度和热抑制温度下种子萌发过程中胚和胚乳贮藏物质的代谢以及相关酶活性变化进行分析,旨在探索温度影响白皮松种子萌发的生理机制,并在一定程度上增进对种子萌发过程中热抑制机理的认识和理解。
1 材料与方法
1.1 试验材料
于2018年10月在甘肃省天水市小陇山(105°30~106°30′E,34°0′~34°40′N),海拔700~2 500 m的天然白皮松林采集成熟的白皮松球果,待球果自然晾干至种鳞开裂时,经人工敲打脱粒并去除种翅和杂质。种子含水率9.92%,生活力96%。
1.2 研究方法
1.2.1 种子萌发试验
1)初始萌发试验。将种子室温吸胀96 h后摆放在铺有湿润脱脂棉的发芽盒中,分别置于10、15、20、25和30 ℃条件下进行萌发试验,每个温度处理3×100粒种子。胚根长度 ≥ 2 mm即视为种子萌发,30 d后试验结束时统计发芽率(初始萌发率)。
2)萌发恢复试验。萌发试验结束后,将各温度下未萌发的种子转移到适宜的温度条件(初始萌发试验获得的理想温度)下继续萌发,观察30 d内种子的萌发情况并统计发芽率(适温萌发率)和腐烂率。
1.2.2 种子萌发过程中内含物的测定
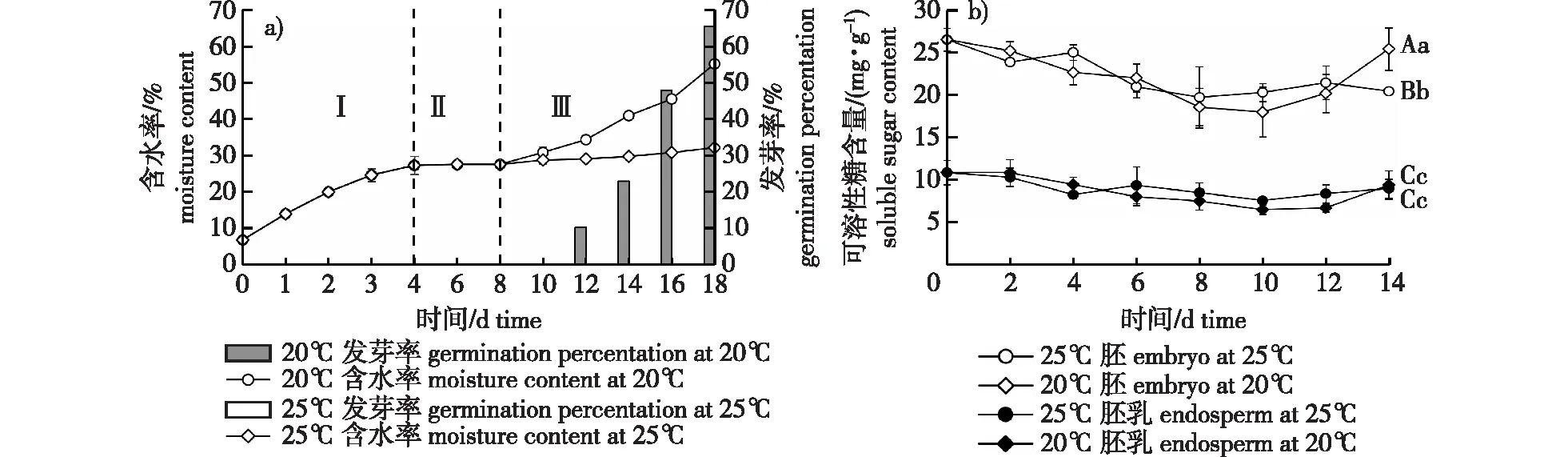
不同大、小写字母分别表示在0.01和0.05水平上差异显著。下同。Different uppercase letters and lowercase letters indicated significant differences at the 0.01 and 0.05 levels, respectively. The same below.图1 白皮松种子萌发过程含水率、发芽率和可溶性糖含量的变化Fig. 1 Changes of water content,germination and soluble sugar content percentage of Pinus bungeana seeds during germination
将种子室温吸胀96 h后摆放在铺有湿润脱脂棉的发芽盒中,分别置于20 ℃(适宜萌发的温度)和25 ℃(产生热抑制的温度)培养箱中培养。试验过程中每2 d取样1次,分别用于含水率(种子吸胀期间每天检测含水率)和生理指标的测定,14 d后结束取样(此时适温20 ℃条件下已有半数种子“露白”即胚根刚突破种皮)。每次取样后抽取3×40粒种子,用小钳子夹裂后去掉种皮,剥出种胚、胚乳切片混匀,分别称取胚和胚乳各0.3 g,贮存在-80 ℃冰箱用于生理指标的测定。
1)含水率的测定。每次取样后,取出3×30粒种子,擦干表面水分后,用小钳子将种子夹裂去掉种皮,参照《International rules for seed testing》林木种子检验规程[14],采用低温烘干法对胚乳(包括胚)进行含水率测定。
2)贮藏物质及相关酶活性的测定。可溶性蛋白含量的测定采用考马斯亮蓝-G250染色法,可溶性糖和淀粉含量的测定采用蒽酮比色法,粗脂肪含量的测定采用索式抽提法,淀粉酶活性的测定采用3,5-二硝基水杨酸法[15]。蛋白酶活性的测定采用Folin-酚法[16]。酸性磷酸酶活性的测定采用试剂盒(购于南京建成生物工程研究所)法[17]。
1.3 数据处理
用Excel 2011处理数据和绘制图表,SPSS 19.0进行方差分析和相关性分析。
2 结果与分析
2.1 温度对种子萌发的影响
试验结果表明,白皮松种子在15和20 ℃温度条件下均具有较高的初始发芽率(87.33%和92.67%)。温度升高至25和30 ℃,种子初始发芽率急剧降低至10%以内。温度过低,种子亦不能萌发,10 ℃下种子初始发芽率为0。初始萌发试验结束后,将各温度下未萌发的种子转移到20 ℃下(适宜温度)继续培养30 d后发现,在25和30 ℃中未萌发的种子均能在适宜的温度下恢复萌发,各温度下种子最终萌发率(初始萌发率与适温萌发率之和)达85%以上且在不同温度间无显著差异(P≥0.05)。萌发恢复试验结束后,各温度下种子的腐烂率均在10%左右,其中25和30 ℃条件下腐烂率稍高,分别为11.33%和12.00%。
2.2 种子萌发过程中含水率的动态变化
20 ℃条件下,种子萌发过程中(包括吸胀)的水分变化呈现“快—慢—快”的节奏,表现出“S”形变化规律(图1)。由图1a可知,整个吸水过程大致可以分为3个阶段:第 Ⅰ 阶段0~4 d,属于吸胀吸水时期(吸胀阶段),种子快速吸水,含水率急剧增加,为萌发提供充足的水分;第 Ⅱ 阶段4~8 d(置床0~4 d),为水分吸收滞缓期,种子吸水缓慢,相对含水率变化不大(27%~28%),为萌发生理生化活动做准备。10 d(置床6 d)后,种子逐渐开始萌发,形态上也发生了巨大的变化,外观上表现为胚乳吸水膨大导致内种皮破裂,然后珠孔端外种皮开裂,继而胚根萌动。此时吸水进入第 Ⅲ 阶段,即生长吸水时期,种子含水率迅速增加,细胞生命活动旺盛。18 d(置床14 d)后,种子发芽率达到60%以上,含水率达到55.21%,是刚置床时(4 d)含水率的2倍、初始含水率的8倍。25 ℃条件下,种子吸水的Ⅰ、Ⅱ 阶段与20 ℃下种子水分变化规律一致,二者含水量曲线基本重合。但18 d(置床14 d)后,该温度下基本上无种子萌发,吸水也停留在第Ⅱ阶段滞缓期,未进入生长吸水时期(第 Ⅲ 阶段),种子含水率略有上升但变化不大,最终含水率为32.17%(图1a)。
2.3 种子萌发过程中营养物质代谢变化
2.3.1 可溶性糖含量变化
可溶性糖类是种子萌发初期碳水化合物中最直接的利用物质,为细胞分裂增殖和胚生长提供原料和能源物质。20 ℃条件下,置床后胚中可溶性糖含量逐渐降低,6 d后含量下降为22.02 mg/g,比0 d含量降低了4.54 mg/g(图1b)。6~10 d可溶性糖分解速度加快,10 d时含量降至最小值17.97 mg/g。之后,随着萌发的进行,可溶性糖含量开始回升,14 d时胚中可溶性糖含量升高至25.45 mg/g。25 ℃条件下,0~6 d胚中可溶性糖含量由26.56 mg/g下降到20.97 mg/g,此后含量变化不大,14 d含量为20.44 mg/g,极显著低于20 ℃下胚中可溶性糖含量(P<0.01)。在两个温度条件下,胚乳中可溶性糖含量与胚中含量变化趋势基本一致,但变化幅度较小,14 d时不同温度下胚乳可溶性糖含量差异不显著。
2.3.2 淀粉和淀粉酶活性变化
淀粉是种子中最普遍存在的贮藏糖类,在淀粉酶的催化作用下分解成小分子糖类,为新生组织和器官提供原料或作为呼吸基质为萌发提供能量。20 ℃条件下,种子中的淀粉含量随萌发时间的延长呈逐渐降低的趋势(图2a)。6 d时胚中淀粉含量为54.89 mg/g,较0 d时含量降低了5.10 mg/g。此后淀粉水解速度加快,14 d时胚中淀粉含量降至43.63 mg/g。25 ℃条件下,胚中淀粉未发生明显降解,14 d时淀粉含量为56.38 mg/g,仅比0 d含量降低3.61 mg/g,极显著高于20 ℃条件下胚中淀粉含量(P<0.01)。20和25 ℃条件下,胚乳淀粉水解速度均较为缓慢,14 d时淀粉含量分别为17.01和20.99 mg/g,且二者之间无显著差异。
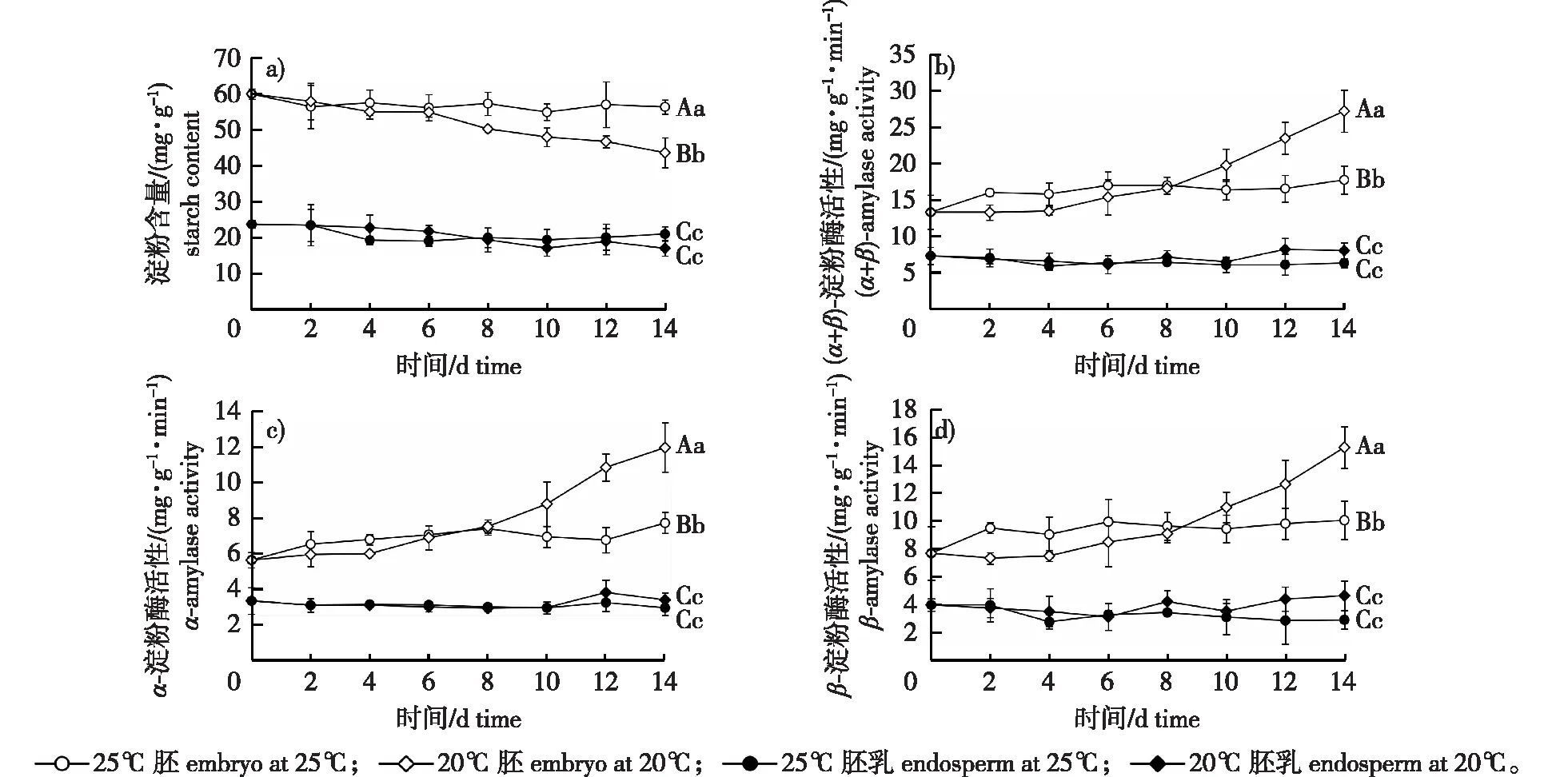
图2 白皮松种子萌发过程淀粉含量和淀粉酶活性变化Fig. 2 Changes of starch content and amylase activity of P. bungeana seeds during germination
20 ℃条件下,胚中淀粉酶活性随萌发时间的延长而增强(图2b—2d)。6 d时胚中(α+β)-淀粉酶、α-淀粉酶和β-淀粉酶的活性分别提高至15.37、6.90和8.47 mg/(g·min)。之后新的淀粉酶开始合成,(α+β)-淀粉酶、α-淀粉酶和β-淀粉酶活性大幅升高,14 d时3种酶活性约为25 ℃条件下胚中淀粉酶活性的1.6倍,差异达到极显著水平(P<0.01)。在两个培养温度下,胚乳中(α+β)-淀粉酶、α-淀粉酶和β-淀粉酶活性均未出现明显变化,不同温度之间的酶活性亦无显著差异。
2.3.3 可溶性蛋白和蛋白酶活性变化
吸胀种子酶系统激活和细胞膜修复等活动都需要贮藏蛋白或其降解产物参与,产生的氨基酸可用于新蛋白合成,为胚的生长提供氮素营养。20 ℃条件下,胚中可溶性蛋白含量整体上表现为降低趋势(图3a)。4 d时,胚中蛋白含量为32.42 mg/g,比0 d降低2.56 mg/g。4~10 d,新的结构蛋白和酶蛋白开始合成,胚中蛋白含量重新回升至36.61 mg/g。10~14 d,随着胚根伸长,胚中蛋白被大量分解消耗含量降低。25 ℃条件下,胚中可溶性蛋白未出现大幅降解,与0 d时含量仅相差1.66 mg/g,14 d时含量极显著高于20 ℃条件下胚中含量(P<0.01)。胚乳中可溶性蛋白含量在两个温度培养过程中变幅均不大,波动范围分别为21.96~25.63和21.86~24.21 mg/g。
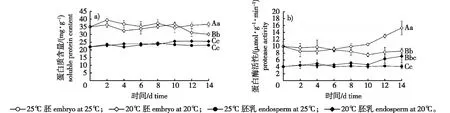
图3 白皮松种子萌发过程可溶性蛋白含量和蛋白酶活性变化Fig. 3 Changes of soluble protein content and protease activity of P. bungeana seeds during germination

图4 白皮松种子萌发过程粗脂肪含量和和酸性磷酸酶活性变化Fig. 4 Changes of crude fat content and acid phosphatase activity of P. bungeana seeds during germination
20 ℃条件下,胚中蛋白酶活性整体上表现为升高趋势(图3b)。种子置床后,胚中蛋白酶活化,10 d时酶活性提高至10.5 μmol/(g·min)。此后,蛋白酶活性快速提高,14 d时酶活性为15.31 μmol/(g·min)。25 ℃条件下,胚中蛋白酶活性无明显升高过程,14 d时酶活性为8.57 μmol/(g·min),略低于0 d酶活性9.92 μmol/(g·min),极显著低于20 ℃条件下胚中的胚蛋白酶活性(P<0.01)。20 ℃条件下,胚乳中蛋白酶活性在0~10 d变化不大,波动范围为4.07~5.09 μmol/(g·min),10 d后酶活性逐渐升高,14 d时酶活性为7.08 μmol/(g·min),显著高于25 ℃条件下胚乳蛋白酶活力水平4.09 μmol/(g·min)(P<0.05)。
2.3.4 粗脂肪含量和酸性磷酸酶活性变化
种子中贮藏的脂肪不仅可以为胚的生长提供能量,也可通过糖异生为新生组织构建提供碳源。脂肪是白皮松种子最主要的贮藏物质,其含量占60%以上(图4a)。20 ℃条件下,0~10 d胚中脂肪含量变化不大(552.17~545.78 mg/g),10 d后脂肪开始分解,14 d时含量降至351.64 mg/g,比0 d时降低了200.53 mg/g;25 ℃条件下,14 d时胚中脂肪含量为482.54 mg/g,比0 d时降低了69.63 mg/g,极显著高于20 ℃胚中脂肪含量(P<0.01);在两个温度下,胚乳中脂肪均未发生明显降解,始终处于较高含量水平668.01~703.73 mg/g。
20 ℃条件下,0~8 d胚中酸性磷酸酶活性基本保持不变,此后酶活性快速升高(图4b)。14 d时酶活性为23.00 μmol/(g·min),比8 d时酶活力水平提高了7.7 μmol/(g·min),极显著高于25 ℃条件下胚中酸性磷酸酶活性15.50 μmol/(g·min)(P<0.01)。20 ℃条件下,胚乳中酶活性在8 d后有所升高,14 d时酶活性为11.09 μmol/(g·min)。25 ℃条件下,胚乳中酸性磷酸酶活性变化不大,波动范围为8.10~9.20 μmol/(g·min)。
2.4 各指标相关性分析
相关性分析结果表明,在20 ℃条件下,随着萌发进程的推进,种子发芽率不断升高,胚和胚乳中营养物质含量和相关酶活性均发生了相应改变(表1)。其中,发芽率与种子中淀粉酶、蛋白酶和酸性磷酸酶活性存在极显著正相关关系(P<0.01),与种子中淀粉、可溶性蛋白和胚中粗脂肪含量之间存在极显著负相关关系(P<0.01),这表明在20 ℃条件下营养物质的含量变化和酶活性的升高在种子萌发中发挥着重要作用。另外,胚中可溶性蛋白含量与蛋白酶活性间、淀粉含量与淀粉酶活性间、粗脂肪含量与酸性磷酸酶活性间均呈极显著相关(R2=-0.578**,R2=-0.827**,R2=-0.837**),说明胚中储藏物质在相关酶的作用下不断分解而含量下降,为胚的生长提供源源不断的能量和营养。但在白皮松种子萌发初期,胚根的生长会优先利用胚自身储藏的营养物质,胚乳中储藏的营养物质尚未发生明显降解,所以各营养物质含量与其相关酶活性之间无显著相关性;在25 ℃条件下,14 d内种子发芽率不足1%,细胞内代谢活动微弱,营养物质的分解代谢活动尚未全面启动(表2)。发芽率与胚中粗脂肪含量极显著负相关(R2=-0.598**),与淀粉酶活性显著正相关(R2= 0.435*)。胚乳中淀粉、可溶性蛋白含量与其相关酶活性显著负相关(P<0.05),二者在酶的作用下分解为小分子糖类,用于种子的呼吸代谢耗能。胚中淀粉在淀粉酶作用下水解产生的糖类随即用于维持自身的呼吸作用,可溶性糖含量与淀粉酶活性显著负相关(R2=-0.621*)。

表1 20 ℃条件下白皮松种子萌发过程中发芽率与胚和胚乳生理指标相关性分析

表2 25 ℃条件下白皮松种子萌发过程中发芽率与胚和胚乳生理指标相关性分析
3 讨 论
3.1 种子萌发对温度的响应
不同植物种类对萌发温度的需求不同,这决定了植物的分布规律,以其限制在适宜的温度区域内生长。许多乔灌木的种子能在较大温度范围内萌发,如北美短叶松(Pinusbanksiana)种子在15~27 ℃的温度内,发芽率没有显著差异。挪威云杉(Piceaexcelsa)和蓝粉云杉(Piceapungens)在16~32 ℃范围内的各种温度下,种子都能顺利发芽[18]。中间温度25 ℃适合多种植物如南京椴(Tiliamiqueliana)[19]、宝华玉兰(Magnoliazenii)[20]和大花四照花(Cornusflorida)[21]等种子的萌发。而有些种子需要在特定的温度范围内才能发芽良好。本研究表明,白皮松种子适宜的萌发温度范围较窄,在15~20 ℃发芽率达90%左右,温度超过20 ℃则萌发过程受阻,25 ℃条件下发芽率大幅降低(小于10%)。白皮松种子发芽对温度的响应是其长期对生长环境选择与适应的结果。种源产地春季日间平均最高气温17~26 ℃,夜间平均最低气温4~12 ℃,日间平均气温10~19 ℃,当地白皮松种群在春季达到了最佳的发芽条件。梣属(Fraxinus)植物也有类似的发芽习性,如在高温25 ℃条件下培养时间超过3 d,初生休眠已解除的水曲柳(Fraxinusmandshurica)种子发芽率将急剧下降,并逐渐进入热休眠状态[22]。花白蜡(Fraxinusornussubsp.cilicica)[23]种子在15 ℃下发芽率较高,20 ℃以上的恒温或变温模式下发芽率迅速降低,并且诱导了种子次生休眠(热休眠)的产生。萌发恢复试验表明,那些在高温下未萌发的白皮松种子依然保持着较高的活力水平,转移到适宜温度下不需要任何解除休眠的预处理措施即可恢复萌发。可见,高温对白皮松种子萌发的抑制作用是可逆的,即热抑制效应[24],区别于梣属植物种子的热休眠。当萌发的温度降低时,高温对草本植物莴苣(Lactucasativa)和二棱大麦(Hordeumvulgaresubsp.distichon)[25]种子萌发产生的抑制作用同样可以逆转。在这些情况下,种子并未进入次生休眠状态或者失活,而是保持不萌发的“静止”状态以应对不利的外部高温环境。
3.2 温度对种子萌发过程中水分含量变化的影响
当种子与水分接触后,立即快速吸水,为各类贮藏物质的水解和多种酶的活化准备充足的水分。之后由于内、外种皮的束缚导致种子内部膨压增大,细胞吸水受阻,此时进入种子的水分几乎为零。种子利用已吸收的水分进行代谢活动,代谢的恢复则需要相关酶和其他功能蛋白的催化[26],而这些代谢活动与种子特性和水合过程中环境条件的关系密不可分。20 ℃条件下,白皮松种子呼吸作用逐渐增强,酶促反应加剧,胚生命活动趋于旺盛,营养物质发生分解,细胞渗透压上升产生渗透吸水,胚生理活动逐渐由量变到质变转化,种子开始萌动。该结果与花旗松(Pseudotsugamenziesii)[27]种子萌发过程水分变化规律相一致。25 ℃条件下,由于温度过高致白皮松种子呼吸速率和酶促反应被抑制,细胞代谢活动停滞而不能顺利进入生长吸水阶段。类似地,在豆科植物家山黧豆(Lathyrussativus)中,超最适温度条件下则种子硬实性增加,从而阻止了种子的水合作用和萌发进程[28]。在不同温度下,种子水合程度的差异影响了萌发过程中的代谢活动进而表现为发芽率的差异。
3.3 不同萌发温度条件下种子的营养物质代谢规律
在种子萌发过程中,除吸收水分以外种子一般无需从外界摄取营养物质。萌发初期代谢所需酶类及大分子类物质已普遍存在于干种子中,吸胀后代谢活性快速恢复,呼吸和代谢作用显著增强,贮藏物质在酶的催化作用下被分解为小分子化合物,为新细胞和组织的构建提供大量的基础材料[1]。温度则可以影响种子内部酶活性的变化和营养物质代谢活动,从而刺激种子对外界环境温度产生不同的响应。
3.3.1 适温萌发时贮藏物质与酶活性的动态变化
种子在适宜的外界环境中,通过启动内部一系列的酶促反应,贮藏的营养物质得以转化与利用。适温20 ℃条件下,白皮松种子从相对静止状态变为生理代谢活跃状态,胚中营养物质分解与合成代谢活动强烈,但胚乳中的贮藏物质并未出现明显降解。这是由于在成熟干燥的白皮松种子中,胚自身储存了一定量的营养物质,种子在萌发时优先利用胚中贮藏的物质。且萌发初期胚乳中的物质并不能运送到胚中供其呼吸和代谢利用,萌发前期营养物质的消耗主要来自胚自身的碳水化合物。种子置床后,胚中贮藏的可溶性糖被用于合成胚轴伸长所需的物质以及呼吸代谢消耗,含量降低。而后淀粉逐渐水解成小分子糖类和其他碳水化合物,以保证萌发初期物质和能量的供应充足。随着萌发过程的继续推进,胚中贮藏的蛋白质在酶的催化下降解为溶解状态的氨基酸(酰胺类)运送到胚参与新生细胞原生质的构成,或经过脱氨基作用转化成糖类供给胚生长利用。脂肪作为白皮松种子最主要的贮藏物质,其利用时间最迟,分解发生于子叶高度充水、根茎显著生长时期[29]。胚根突破种皮后,胚中脂肪在酸性磷酸酶作用下大量降解为小分子物质(甘油和游离脂肪酸),或者经过糖异生途径将分解产物转化为糖类用于胚的生长和幼苗形态建成。所以,脂肪含量在萌发过程中降低时也伴随着碳水化合物含量的显著增加[30]。白皮松种子中储藏物质的利用与油料作物芝麻(Sesamumindicum)[31]、欧洲榛子(Corylusavellana)[32]和阿月浑子(Pistaciavera)[33]种子萌发代谢顺序基本一致。而在油松(Pinustabuliformis)种子萌发的初始阶段,最先利用贮藏物质是蛋白和脂肪,二者的动员快于糖类和淀粉[34]。在花旗松种子中,脂类物质降解后引起碳水化合物含量变化进而种子萌发[27]。在萌发的挪威云杉(Piceaabies)种子中,蛋白质的水解发生于脂类的动员之前[35]。虽然这些植物的种子均属于高脂肪类种子,但不同的植物种子萌发时储藏物质的动员顺序并不一致。
3.3.2 高温萌发时种子内部生理活动对热抑制的响应
热抑制是一种被动状态,通过高温诱导生理功能的变化而施加于处于温度胁迫条件下的种子。升高的温度直接作用于参与种子萌发抑制机制的单一反应,或者通过关键成分的变性亦或通过增强与抑制物质合成相关的过程来诱导功能的缺失[36]。高温往往会破坏酶结构使酶活性降低甚至失活,阻碍植物正常的代谢过程。25 ℃条件下,过高的温度阻碍了白皮松种子萌发进程的推进,吸胀种子微弱的呼吸作用消耗了部分贮藏糖类物质来维持相对“静止”状态。同时,高温抑制了种子中酸性磷酸酶活性和脂肪的降解,蛋白酶的低活力水平影响了蛋白质的分解以及新的功能蛋白酶类等的合成与加工,并且导致了胚对小分子物质可溶性糖和可溶性蛋白吸收利用的减少。相类似,25 ℃条件同样减缓了休眠已解除的水曲柳种子中淀粉酶活性的变化,阻碍了脂肪水解酶的合成,并减弱了种子中糖类和蛋白为幼胚生长提供能量和氮源的能力,从而抑制种子萌发并产生二次休眠[37]。值得注意的是,白皮松种子在高温下萌发时,各种酶活性均处于低水平状态,但其只是暂时被抑制而并未变性失活,这在萌发恢复试验中最终的高发芽率中可得以反映。即种子萌发的这种不耐热因子是可逆的,当回到适宜的温度时,其再活化的程度取决于其失活的程度[38]。报春花科植物Myrsineparvifolia[39]在25和30 ℃条件下其种子发芽被完全抑制,将种子转移到20 ℃培养后萌发被重新激活,但在高温下培养的时间越长,种子发芽前需要在低温下的时间就越长。
4 结 论
在种子萌发事件中,贮藏物质在酶的催化和调节作用下经动员后转变成可运输的形式并运送到需求的位点,以支持产能和合成反应。本研究得出,白皮松种子萌发的适宜温度条件为15~20 ℃,温度继续升高则发芽率急剧降低产生热抑制现象;在适温20 ℃条件下,白皮松种子内催化大分子物质分解的各类酶均呈现出高活力水平,淀粉、蛋白和脂肪等贮藏物质顺利降解。且其在种子萌发时优先利用胚自身贮藏的物质,最先利用的是胚中的碳水化合物,然后是蛋白质和脂类的动员,脂肪的大量降解始于胚根突破种皮后。在热抑制温度25 ℃条件下,白皮松种子中淀粉酶、蛋白酶和酸性磷酸酶的活性被高温所抑制,营养物质的分解代谢受阻,胚不能吸收和利用到足够的营养来完成萌发过程。
