海金沙植物的繁殖特性及植株生长习性研究进展
2023-11-28童家赟黄满霖魏汝芯詹若挺
童家赟, 黄满霖, 魏汝芯, 詹若挺,2
[1.广州中医药大学中药学院,广东广州 510006;2.岭南中药资源教育部重点实验室(广州中医药大学),广东广州 510006]
中药海金沙来源于蕨类植物海金沙科海金沙属草本植物[,Lygodium japonicum(Thunb.)Sw.]的干燥成熟孢子,具有清热利湿、通淋止痛的功效[1]。现代研究表明,海金沙提取物能够促进输尿管的蠕动,增加输尿管上段压力,从而有利于结石排出;还能降低肾脏中草酸和钙离子含量,增加排尿量,抑制草酸钙晶体从亚稳态转化为稳态晶体,从而降低结石形成风险[2-6]。海金沙药材为临床常用中成药尿石通丸、复方石淋通片、肾石通颗粒、三金排石汤、排石颗粒等的重要原料。其中尿石通丸、复方石淋通片是治疗尿路结石的常用药,为国家二级中药保护品种,已列入基本药物目录[7]。据了解,仅东莞市亚洲制药科技有限公司对海金沙孢子粉的年需求量就超过50 吨。然而,海金沙原植物常散生于自然环境中,一直以来只能通过人工采收,药材采收效率极低。随着经济的发展和人工成本的提高,药农采收野生海金沙药材的积极性急剧降低,供不应求导致近年来该药材的价格较快上涨,相关中成药的生产和可持续发展受到严重制约。此外,同属植物曲轴海 金 沙(L. flexuosum)和 小 叶 海 金 沙(L.microphyllum)与海金沙形态非常相似,地理分布亦有重叠,常有误采和混用现象[8]。因此,海金沙这种较大宗的野生来源药材的生产,急需开展集约化和规模化种植来保证产量和临床用药的质量。以下对海金沙繁育和植物生长条件相关的研究概况进行综述,以期在前人研究的基础上优化其繁育方法和条件,为最终解决其药材生产问题提供参考。
1 海金沙的繁殖特性及繁殖机制研究概况
1. 1 海金沙世代生活史中的组织形态的发生发育相较于种子植物从种子萌发到幼苗形成的速度,蕨类植物的幼苗形成则较缓慢和复杂。蕨类植物具有明显的有性世代的配子体(单倍体)和无性世代的孢子体(二倍体)交替现象。早在1991年,国内学者对海金沙(Lygodium japonicum)从孢子萌发到长成幼孢子体苗的全过程进行了较为仔细的观察,研究表明整个过程总共约需半年时间[9]。海金沙孢子经无菌播种6 d 后萌发,形成丝状体,接着长成心形的配子体,播种后约30 ~35 d 生成精子器,接着生成颈卵器;成熟的配子体授精后约半个月长成幼孢子体,再经过4个月可以长成具3片叶的幼苗[9]。之后,日本的高桥尚子(Takahashi N)等[10]采用生物显微镜和金相显微镜观察了海金沙孢子经萌发后形成顶端细胞分生组织(apical-cellbased meristem)和多细胞分生组织(multicellular meristem)两种分生组织,以及细胞经过一系列平周和垂周分裂后而形成心形配子体的全过程。刘朝辉等[11]采用透射电镜观察了海金沙的精子细胞发育过程,发现其结构发育与薄囊蕨亚纲的其他植物相似。郭严冬、Cao J G 等[12-13]采用树脂包埋切片、透射电镜和细胞化学技术观察了海金沙配子体发育生成颈卵器和卵细胞的过程。邵文等[14]采用石蜡切片观察了海金沙颈卵器中卵子受精后产生合子、合子进行细胞分裂形成四分体、最后形成基足细胞和营养器官(第一和第二叶)的整个过程。由以上研究可知,目前海金沙从孢子萌发到长成幼孢子体的组织形态发生全过程已经比较清楚,其示意图见图1。
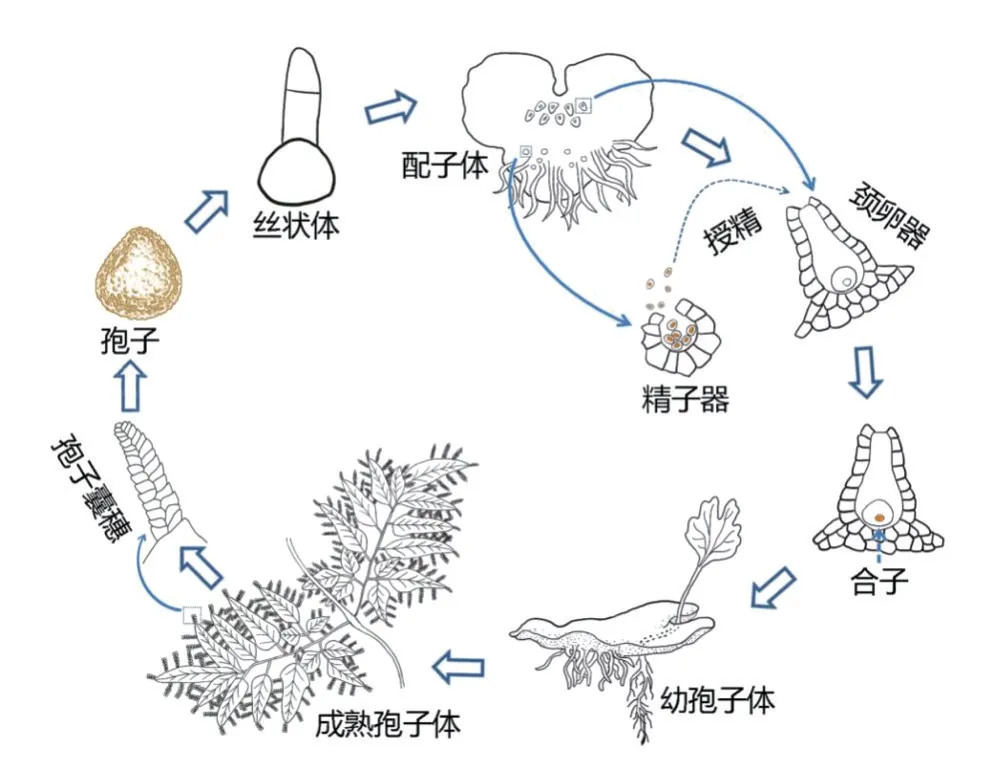
图1 海金沙生活史示意图Figure 1 Schematic diagram of the life history of, Lygodium japonicum
1.2 海金沙配子体性别决定机制
1. 2. 1 遗传因素和内在激素调控雌雄配子体的生成 早在1960 年,Ulrich N[15]就证实了在无菌培养过程中,海金沙的成熟配子体提取物添加入培养基后能诱导其他年幼配子体提前产生精子器。1975 年,竹野清俊(Takeno K)等[16-17]发现外源的赤霉素类(gibberellins,GAs)可诱导海金沙配子体的精子器形成;赤霉素类的活性强度由大到小排列为GA7>GA4>GA9>GA3>GA5>GA1>GA8;随 后发现外源的赤霉素类会抑制颈卵器的生成,活性强 度 由 大 到 小 排 列 为GA4= GA9>GA7>GA3>GA1=GA5=GA8。1979 年,竹野清俊等[18-19]验证了海金沙配子体提取物也会抑制颈卵器的分化;随后在原叶体培养基中加入外源的成精子囊素(antheridiogen)如GA9等,发现GA9的抑制效应与配子体提取物相当,认为其机制可能与由GA9转化生成的GA9-甲酯发挥的效应有关;但是其控制配子体在不同时间点产生性别分化的机制尚不明确。
竹野清俊等[20]基于以上研究之后,通过实验观察提出了海金沙配子体性别分化时间序列模型:当居群内的原叶体(配子体)处于不同的发育阶段时,最先达到1.5 mm 宽的原叶体形成颈卵器,这时只发育形成单性的雌性配子体。然后,原叶体开始生成控制生殖器官形成的天然物质;但这些物质并不影响那些较大的正在分化颈卵器的单性的雌性原叶体。而居群内发育迟缓的原叶体对这些大的原叶体分泌的物质非常敏感,使得颈卵器的形成受到抑制,从而产生精子器,也就是使这些更小的原叶体发育形成单性的雄性配子体。最终,雌性原叶体可以在不受外源成精子囊素的影响下产生精子器,从而产生双性配子体。
2014 年,日本的田中君木(Tanaka J)等[21]克隆了海金沙赤霉素合成途径中相关基因并作表达模式分析,并通过蛋白双杂交和免疫印迹等一系列试验,阐明了蕨类植物赤霉素合成途径及其信号传导的时空分离是决定配子体性别的重要分子机制。田中君木等认为,首先,早熟的配子体生成GA9,然后被甲酯化成GA9-甲酯, GA9-甲酯被分泌到环境中,从而被邻近的晚熟配子体(丝状体阶段)摄取;GA9-甲酯被摄取之后,在晚熟的配子体细胞内被脱甲酯重新生成GA9,之后GA9被GA3氧化酶(GA3ox)催化生成GA4,接着具有生物活性的GA4通过GA4→GID→DELLA 通路触发了配子体上精子器的生成,但抑制了颈卵器的发生。
1.2.2 环境因素影响雌雄配子体的生成 蕨类植物的性别分化除了受到自身遗传因素和内在激素调控外,也受到培养密度、营养条件、光照等环境因素的影响[22]。高晓雯等[23]的研究表明,水分对配子体形态发育的影响也较大。随着水分的增加,海金沙孢子的萌发速率加快,丝状体的长度增加。在水分不足条件下,配子体易发育成雄性配子体,而在水分充足条件下,配子体易发育成两性配子体或雌性配子体。次生假根的长度也可随着水分的增加而增加。但是该研究未对培养过程的基质加水量进行准确计量。后续研究需要参照该报道的方法进一步确定有利于分化生成两性配子体或适当比例的单性配子体及有利于授精产生合子的最适合水分量。
2 海金沙的育苗影响因素研究概况
2.1 影响孢子萌发因素的研究蕨类植物孢子萌发除了受到自身遗传因素和内在激素调控外,也受到培养密度、营养条件、温度、光照等环境因素的影响[24-25]。如紫萁(Osmunda japonica)孢子播种密度在100 ~3 000 个/cm2的区间时,随着密度的升高,孢子的萌发率越高,密度为3 000 个/cm2时的萌发率可达95.4%;而当密度达到5 000 个/cm2时,其萌发率反而降低[26]。目前,海金沙播种密度试验未见相关报道。无机盐方面,郭治友等[27]的研究认为,低盐的Knop’s培养基和全量(高盐)的MS培养基对海金沙孢子萌发影响不显著。但也有研究显示,Knop’s 培养基、1/8 的MS 培养基和1/4 的MS培养基上的孢子萌发率较MS培养基更高[28]。两篇报道互相矛盾,有待进一步验证。
有研究认为,冷冻保存可显著降低小叶海金沙(L.microphyllum)孢子萌发率,但是对海金沙孢子萌发无显著影响[29]。海金沙孢子在低温条件下保存多长时间后而不会显著降低萌发率,这也是值得探讨的问题。
2.2 雌雄配子体交配和幼孢子体生成的研究竹野清俊(Takeno K)等[30]开展的分离双性配子体并与单性配子体进行交配的试验显示,只有约30%的雌性配子体生成幼孢子体,而单个的双性配子体未能形成幼孢子体;并认为该结果可能是由配子体内自交产生的纯合子不能发育导致的。但是,Lott M S 等[31]通过研究认为,海金沙孢子萌发后,成熟配子体主要是雌雄同体型,同一配子体内自交高于90%。以上二者的研究结论相去甚远,需要进一步证实。
Sakaushi S 等[32]在高速视频显微镜、透射电子显微镜(TEM)和扫描电子显微镜(SEM)下观察发现,海金沙精子的鞭毛朝向同一方向,并且只能朝前方游动,即使遇到障碍物也不会向后游动;并认为这个特点可能会影响雌雄配子体的授精。到底有多少个孢子形成配子体后能成功授精成为合子并生成幼孢子体,其最适合的环境因素有哪些,目前未见相关的报道。
2.3 幼苗繁育(植株再生)研究海金沙可采用分株繁殖的方式,操作上比较简单[33],但分株繁殖方式的繁殖系数较低,满足不了规模化种植对大量种苗的需求,而通过孢子萌发有可能实现种苗的大规模生产。由于蕨类植物生活史(世代交替史)的复杂性,要通过孢子萌发途径进行海金沙的繁殖和植株再生,相较一般的种子植物步骤更繁琐,要解决的问题也更多。目前仅见Maeda M等[34-35]对海金沙幼孢子体叶片提取的原生质体进行了植株再生的初步研究;郭治友等[27]采用Knop’s 培养基、MS 培养基等进行了孢子无菌萌发和植株再生的初步研究;另有将海金沙孢子播于泥炭土上进行萌发和育苗的专利报道[36]。有关海金沙孢子萌发育苗方面的研究报道非常少,迄今也未见采用海金沙幼芽或嫩叶进行愈伤组织培养扩繁的研究。
3 海金沙植株生长习性研究现状
目前,对海金沙进行规模化和集约化种植的研究尚未见报道,以下只是针对野生或少量盆栽植株生长状态的观察。
3. 1 光照对海金沙植株生长的影响一般认为,蕨类植物大多是阴生植物,海金沙野生植株也常生长在树林下有遮荫的环境中。张泽宏等[37]观察了海金沙等5 种蕨类植物叶片解剖结构,根据叶片薄、没有海绵组织和栅栏组织的分化、气室大等特征,认为这些是蕨类适应阴生环境的特征。但目前海金沙植株生长最适合的光照强度或遮蔽度并未见报道。
3.2 温度与湿度对海金沙生长发育的影响温度与湿度对海金沙植株的生长发育会产生一定的影响,冬季低温和干旱条件下可出现植株枯萎或枯叶[38];对普通温室内培养的11 种蕨类植物生长发育进行为期一年半的观察,只有海金沙和鹿角蕨(Platycerium wallichii)两种植物未生成孢子,推测可能是由于温度与湿度不适合所致[38]。另有学者观察发现,低海拔气候温暖地区的海金沙植株能正常产生孢子,而在高海拔冷凉地区则不能[39]。有研究表明,近缘物种小叶海金沙(L.microphyllum)在较高温度环境中的群落长势明显较好;室内模拟环境下温度与其光合速率呈显著正相关[40]。但适合海金沙植株生长的温度和水分条件范围至今尚不清楚。
3.3 海金沙的病虫害值得注意的是,海金沙和小叶海金沙在美国东南部的佛罗里达州和澳大利亚等地被认为是严重影响生态的入侵植物,多位学者从生物防治的角度研究了真菌感染和飞蛾对海金沙生长的影响。如Rayachhetry M B 等[41-43]学者从染病植株上分离病菌后重新感染植株,发现假柄锈菌(Puccinia lygodii)或者二级霉(Bipolaris sacchari)感染可致海金沙和小叶海金沙叶片坏死。有学者通过观察发现,炭疽病菌(Glomerella cingulata)感染可使海金沙和小叶海金沙顶梢枯死,小羽叶边缘褐变和萎蔫[44-45]。此外,还发现鳞翅目(Lepidoptera)草螟科(Crambidae)的两种飞蛾,Neomusotima conspurcatalis和,Austromusotima camptozonale特 异 性地产卵在海金沙和小叶海金沙的植株上,且其幼虫啃食能有效控制宿主植物的生长[46-47]。在未来规模化种植生产过程中,以上真菌和飞蛾的防治应当引起重视。
4 小结和展望
目前,国内外对于蕨类植物的繁殖特性及繁殖技术研究主要集中于巢蕨等观赏或濒危蕨类植物[48-50],药用蕨类植物相关研究则很少。海金沙从孢子萌发到生成配子体,再长成幼孢子体的全过程,其组织形态发生业已清楚;配子体性别决定机制的遗传因素和内在激素调控也得到了阐明。但是,适合孢子萌发、配子体授精等繁殖过程中的环境因素有待进一步优化;迄今未见规模化的育苗和规范化种植技术研究。
目前,至少还有以下问题需要进一步探讨:①筛选最适合海金沙孢子萌发的条件,包括培养密度、营养量(包括无机盐、蔗糖等)、温度及光照等。②筛选适合孢子贮藏且有利于其在一段时间内保持活力的温度与湿度条件。③筛选孢子形成配子体及授精成为合子,并生成幼孢子的最适合环境条件,如影响配子体授精的温度、水分、培养密度、光照等。④研究建立外殖体(幼芽或嫩叶)进行无性繁殖的离体快繁体系。⑤进行栽培技术研究:如探究适合植株生长和孢子囊生成的光照、水肥、温度等条件;研究田间管理方式,如搭设支架等对植株生长和孢子囊生成的影响;探究适宜的采收期。建议今后应通过对上述影响海金沙育苗和栽培的关键技术条件进行系统的研究、探索和优化,为海金沙药材生产提供技术支撑,以维持其药材的可持续发展及保障有关中成药原料的供应。
(我校中药学院药用植物教研室教师彭光天博士和童毅博士对本文进行了审阅并提出意见和建议,特此致谢!)
