甲烷氧化菌产单细胞蛋白及其在水产饲料中的应用
2023-11-27宋增武孙立瑞锁博海刘思淼葛怿泽辛嘉英
■ 宋增武 孙立瑞 锁博海 刘思淼 葛怿泽 辛嘉英,2*
(1.哈尔滨商业大学食品科学与工程重点实验室,黑龙江哈尔滨 150076;2.中国科学院兰州化学物理研究所羰基合成与选择氧化国家重点实验室,甘肃兰州 730000)
联合国粮食及农业组织预测,到2030 年,水产养殖生物将占世界海产品供应量的53%[1],全球水产养殖产量将增加2 600万吨,额外需要4 000万吨饲料[2],而水产养殖的进一步增长可能受到饲料资源短缺的限制[3-4]。由于水产动物对蛋白质含量要求较高,一般是畜禽的2~4 倍,因此目前水产饲料中蛋白质的主要来源是利用鱼、虾、蟹制成的鱼粉,其中含有高质量的蛋白质和必需氨基酸以及未知生长因子,具有良好的适口性、较高的消化吸收率[5],可保证水产养殖生物保持较快速度生长,提高收益。鱼粉的广泛使用直接导致了鱼粉饲料供应不足,价格居高不下[6-7],因此急需利用现有自然资源寻找可替代蛋白饲料,进而开发高质量的替代饲料资源[8-9]。从这个意义上说,单细胞蛋白(Single Cell Protein,SCP)是替代鱼粉蛋白的有效解决方案[10-11]。
单细胞蛋白,也被称为微生物蛋白、菌体蛋白,是指利用酵母、霉菌、藻类等单细胞生物经过发酵培养和扩增之后获得的生物体蛋白[12-14]。人类在几个世纪前就开始把微生物用作食物和饲料,许多微生物自古以来就被用作饮食的一部分,例如公元前2500 年,发酵酵母就被用作面包的发酵剂,公元前50-100 年,埃及人和希腊人使用乳酸菌处理牛奶和奶酪[15],非洲人把具有“蛋白之王”之称的螺旋藻用作食物来弥补当地人缺少的蛋白质[16]。单细胞蛋白的生产技术是在100多年前发展起来的,而大规模生产是在20 世纪出现的。第一次世界大战前后,德国认识到可以使用微生物作为微生物蛋白质大量生产,因而,面包酵母是在添加铵盐的糖蜜培养基中产生的[17-18]。自此以后从细菌、真菌和藻类培养物中产生的蛋白质被广泛用于食物和饲料中,由于其蛋白质含量高、所含的必需氨基酸种类丰富、脂肪含量低、含有多种微量元素,可以用作膳食补充剂添加在动物饮食中,也可以作为部分蛋白的替代品[19-21]。文章对甲烷氧化菌产SCP的营养特性及在水产饲料中的应用效果等作以综述,综合评价营养价值对水产动物生长性能及健康的影响。为甲烷氧化菌产SCP在水产饲料领域中的利用和开发提供参考依据。
1 甲烷氧化菌的特性及新菌株的分离鉴定
1.1 甲烷氧化菌的特性
甲烷氧化菌是一类利用甲烷作为唯一碳源和能源的革兰氏阴性菌,按照是否利用环境中的氧气作为电子受体,可分为好氧甲烷氧化菌和厌氧甲烷氧化菌两大类。工业上常用于生产单细胞蛋白的是Ⅰ型甲烷氧化菌中的γ-变形菌门,应用较多的是Methylococcus capsulatus(Bath)菌株[22-23]。他们是一类独特的革兰氏阴性细菌,偏好氧气浓度较高的环境,并且更容易进行一些基因操作(见表1)。
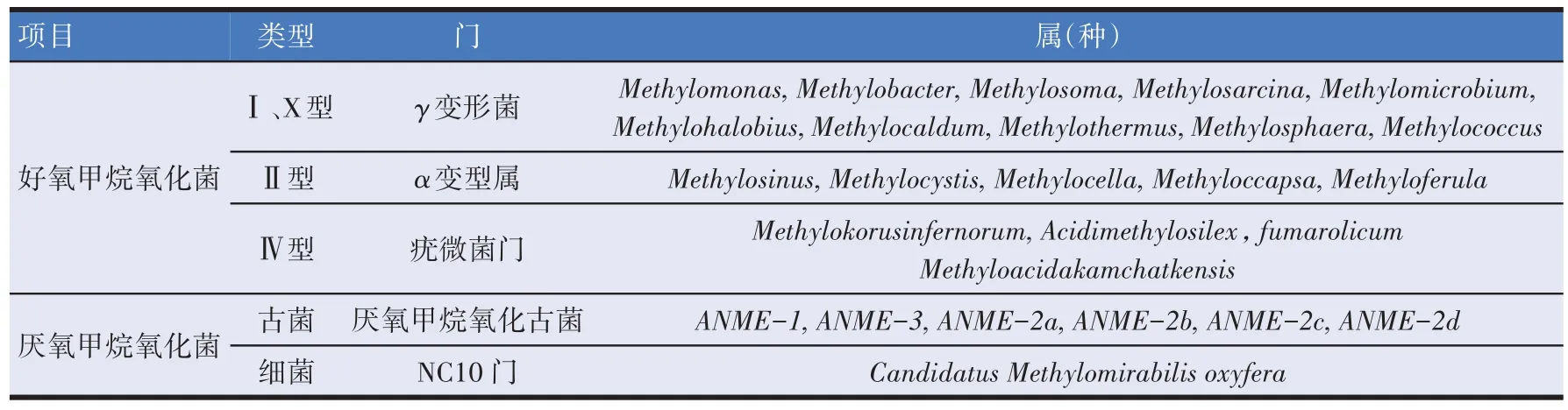
表1 甲烷氧化菌的分类[24-26]
甲烷氧化菌生长时,以甲烷作为碳和能源的主要来源,把甲烷转化成二氧化碳[27-28],产生富含蛋白质的生物质[29-30],生物质中85%~95%来源于CH4[31]。甲烷氧化菌的特征酶是甲烷单加氧酶(MethaneMonooxygenase,MMO)[32]。甲烷氧化菌在MMO 作用下把甲烷氧化为甲醇,在甲烷转化的第一步之后,甲醇被氧化成甲醛,甲醛可以进一步氧化或同化为细胞碳固定途径的底物。甲醇的氧化由甲醇脱氢酶(Methanol dehydrogenase,MDH)催化,对于革兰氏阴性嗜甲烷细菌,甲醇脱氢酶以特定形式存在于胞质中,并利用含有吡咯喹啉醌(Pyrroloquinoline Quineone,PQQ)作为其催化辅助因子。产生的甲醛可以以三种方式被使用:第一种是进一步氧化成CO2;第二种是通过核酮糖单磷酸(RuMP)循环同化;第三种通过丝氨酸循环同化。上述两个循环的操作导致产生丙酮酸,然后产生乙酰辅酶A作为进一步细胞代谢的关键中间体。在RuMP途径中,甲醛用于合成果糖-6-磷酸,并使用指定的核酮糖磷酸合成酶通过糖磷酸中间体进一步转化为核酮糖-5-磷酸,用于生物质生长[33]。
1.2 甲烷氧化菌新菌株的分离鉴定
甲烷氧化菌在自然界中分布十分广泛,湿地、稻田、泥潭沼泽、污水污泥等都有甲烷氧化菌的存在,近年来也有许多学者分理出具有优良蛋白潜质的菌株。顾华兵等[34]从污泥中分离、筛选获得一株甲烷氧化菌MO-01,根据该菌株的形态学、生理生化试验和16S rDNA序列同源性分析,证实该菌株与Methylobacterium zatmanii菌株有99%的同源性,属于Methylobacterium属,优化了甲烷氧化菌的发酵条件,为以后放大生产SCP 提供了科学基础。Thu等[35]从厌氧发酵器的废水中分离出新的菌株——氧化甲烷BG3菌株,确定该菌株为甲基单胞菌属,BG3 菌株的生物质作为牲畜的优质蛋白质来源,具有巨大的应用潜力。
2 甲烷氧化菌生产SCP 及其与其他菌共培养生产SCP
2.1 甲烷氧化菌生产SCP
20 世纪90 年代一家挪威公司用荚膜甲基球菌生产了SCP,但由于当时廉价的植物蛋白、微生物培养技术等限制,市场竞争激烈,造价较高的微生物蛋白无法继续发展。近年来,微生物领域迅猛发展,大豆等植物蛋白价格持续上涨,导致水产饲料成本上涨[36]。而甲烷氧化菌以甲烷为唯一碳源和能量,不受土地、光照等因素限制,且生产出的SCP 蛋白含量高,吸引企业进行产品商业化,表2 为甲烷氧化菌生产单细胞蛋白的商业化产品。
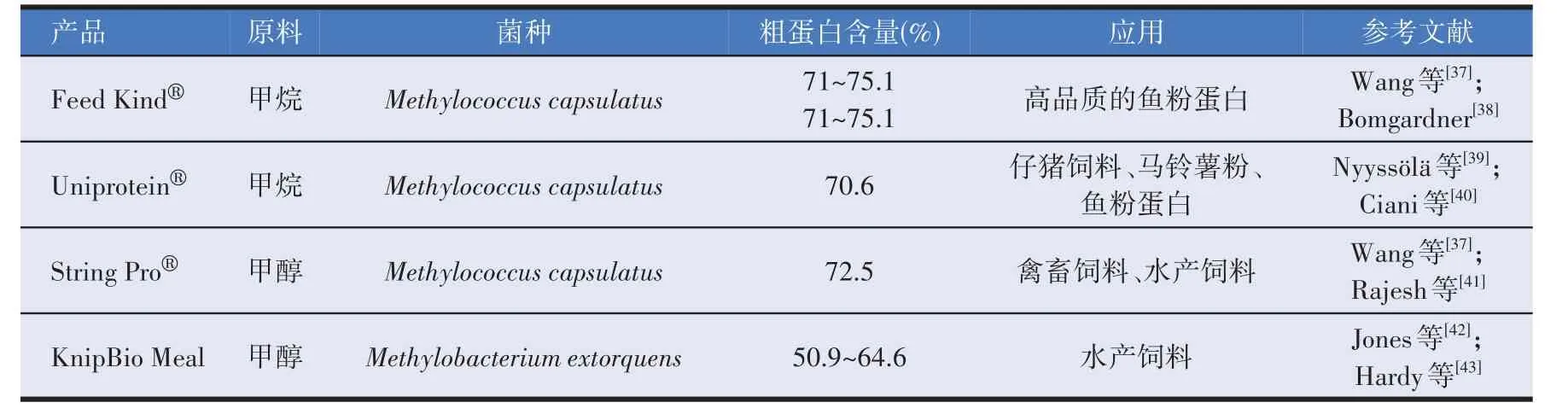
表2 甲烷氧化菌生产的单细胞蛋白商业化产品
2.2 甲烷氧化菌与其他菌共培养生产SCP
甲烷氧化菌来生产单细胞蛋白主要分为单一菌种和两种及两种以上混合菌进行培养生产,单一菌种生长较慢,而混合菌培养中非甲烷菌可以为甲烷菌提供所需的影响因子,可以减少代谢过程中的副产物对甲烷氧化菌的抑制。早在2002年,Bothek等[44]将甲基球菌Bath(Methylococcus capsulatusBath)分别和动脉瘤杆菌属(Aneurinibacillus)、农业短芽孢杆菌(Brevibacillus agri)、罗尔斯顿属(Ralstonia)进行共培养,研究发现非甲烷菌可以除去生物反应器中的有机碳、乙酸等,从而促进甲烷氧化菌的生长。Nunes 等[45]选用了产碱杆菌、动脉瘤芽孢杆菌、农业短芽孢杆菌三种不同的异养细菌,分别进行和甲烷氧化菌共培养。研究发现异养细菌不会利用会在甲烷进行代谢,但是可以消耗反应器中的有机碳,进而增加了生物量的积累浓度,提高单细胞蛋白的产量,为甲烷氧化菌和其他微生物共培养提供了新思路。
Rasouli 等[46]把小球藻(Chlorella sorokiniana)和荚膜甲基球菌(Methylococcus capsulatus)共培养生产SCP,发现可以把工业废水中的营养物质转换为SCP,蛋白质组成与传统的蛋白质来源相似,表明藻类或细菌的生物质可以替代它们作为不同动物的饲料成分。Hill 等[47]把嗜碱性甲基微球菌20z(Methylomicrobium alcaliphilum 20z)和蓝藻聚球菌PCC 7002(SynechococcusPCC 7002)进行共培养,和单一菌种生长组做对比,发现混合菌生长的时间更久,获得更高的生物量,可以为生产SCP 提供新思路。这种共培养技术在利用不同气体流的能力方面是可扩展的,其生物成分由模型细菌构建,可以通过代谢定制来生产一系列生物燃料和生物产品。
肖扎等[48]用甲基单胞菌和嗜甲基单胞菌属进行共培养研究,以污水污泥厌氧消化产生的沼气和废弃废水作为碳氮源来生产SCP,甲基单胞菌的作用是将甲烷氧化为甲醇,而嗜甲基菌的作用是同化积累的甲醇,以避免过量的甲醇对甲基单胞菌生长的抑制[49],所生产的蛋白质可以达到细胞干重的41%以上,其中决定了动物生长情况和鳃结构完整性的组氨酸的含量分别是豆粕和鱼粕的1.67 倍和2.09 倍[50]。孙立瑞等[51]把甲烷氧化菌和光合菌进行共培养,把光合菌和甲烷氧化菌离心后的上清液和超声破碎的沉淀分别加入甲烷氧化菌和光合菌中,进行培养,探究细胞内外物质对彼此生长的促进或抑制作用,发现他们之间存在着一定的促进作用进一步研究发现光合菌在光照条件下吸收甲烷氧化菌同化甲烷产生的CO2,并且产生同化甲烷所需的O2,减少高含量CO2对甲烷氧化菌的抑制,缩短了发酵周期,增加了单细胞蛋白的产量[52]。
3 甲烷氧化菌产SCP 的营养特性及其在水产饲料中的应用
3.1 甲烷氧化菌产SCP的营养特性
用于生产单细胞蛋白的微生物有细菌、真菌、藻类等,微藻生产的单细胞蛋白主要作为营养补充剂的使用[53];真菌生产的单细胞蛋白可以应用在人类饮食中,如香肠汉堡中所添加的是一种由丝状真菌生产的具有类似肉制品质地的单细胞蛋白[54];细菌生产的单细胞蛋白可以应用于畜禽饲料,近些年在水产领域应用较多[55]。表3汇总了不同来源单细胞蛋白和动植物蛋白的优缺点。甲烷氧化菌干燥后蛋白含量占总量的70%左右,并且还含有动物机体所需的多种氨基酸、碳水化合物、多种维生素、核酸、脂肪、无机盐等,含量较高的例如K+、Mg2+、Fe3+和Fe2+等,因此可以添加到饲料里为水产动物、畜禽等补充微量元素[59]。从营养成分组成的角度上说,甲烷氧化菌生产的单细胞蛋白与天然蛋白一致。从必需氨基酸含量上说,与鱼粉相比,甲烷氧化菌生产单细胞蛋白的色氨酸含量较高,赖氨酸含量较低。与棉籽相比,甲烷氧化菌生产SCP 的含有更多的苏氨酸和蛋氨酸。表4 为甲烷氧化菌产的单细胞蛋白和其他饲料中氨基酸含量。
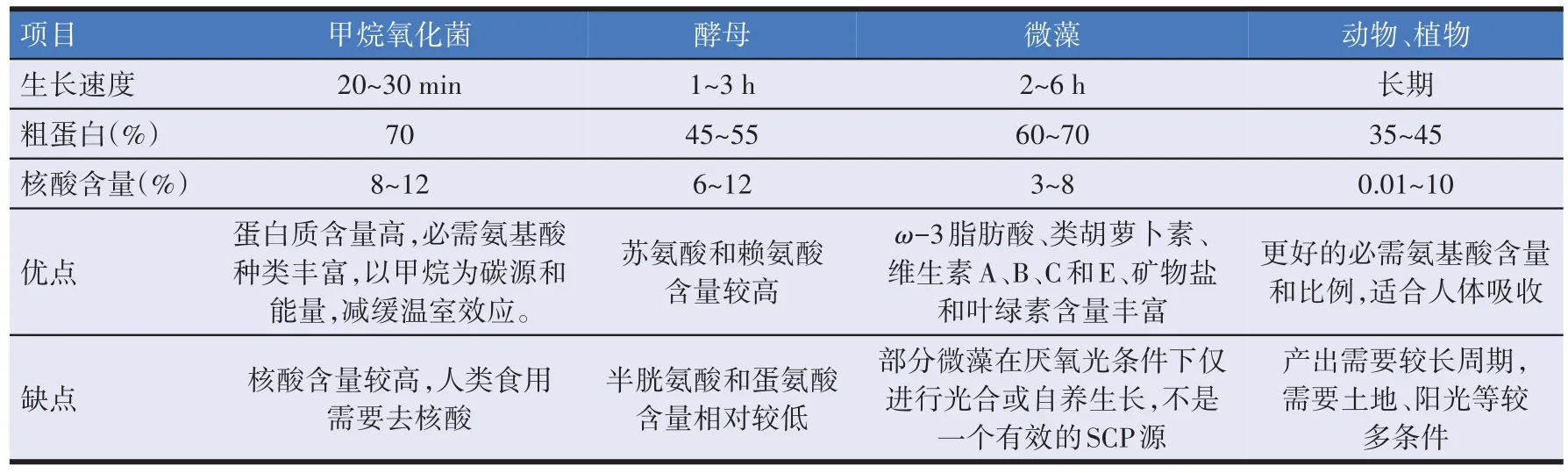
表3 不同来源蛋白的优缺点[56-58]
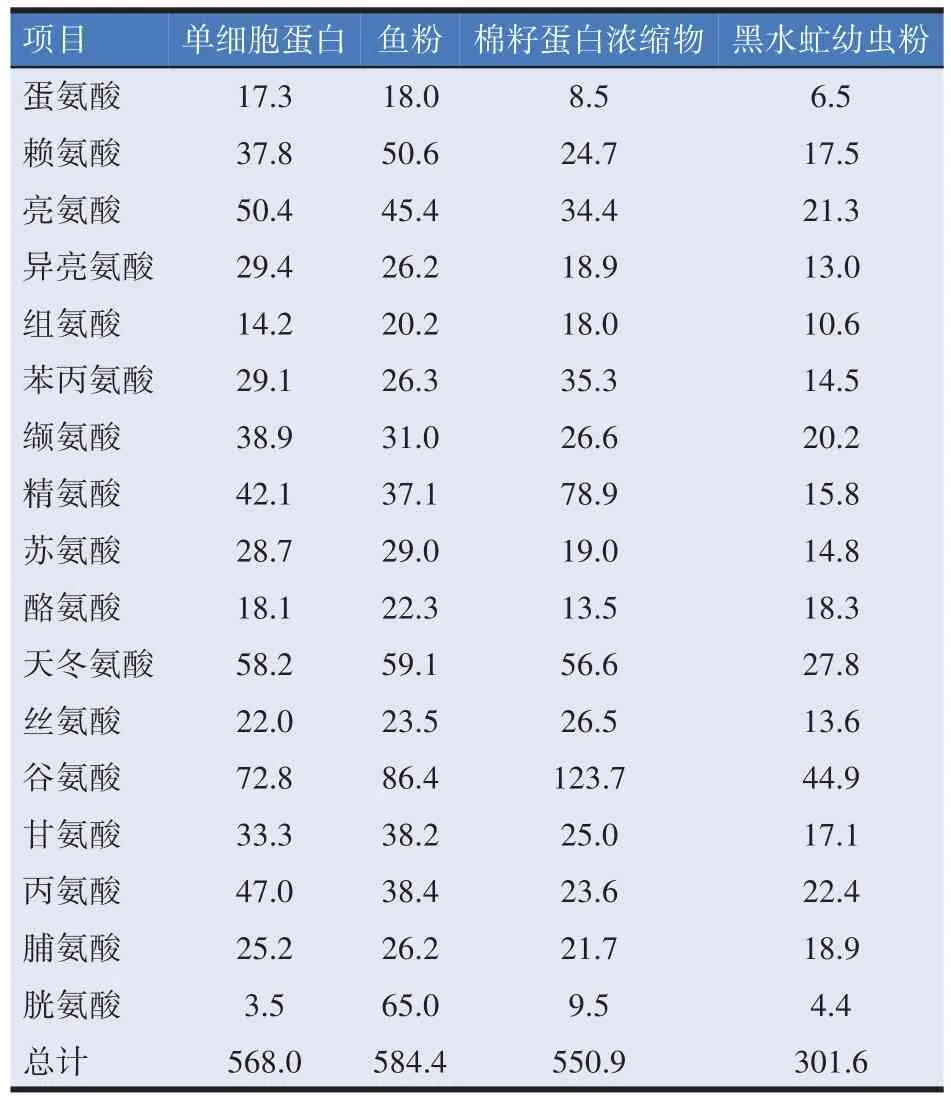
表4 甲烷氧化菌产SCP和其他饲料氨基酸组成(g/kg干物质)[60-61]
3.2 甲烷氧化菌产SCP在水产饲料中的应用
经过国内外专家进行不同物种的饲养实验,已经明确甲烷氧化菌生产的SCP 可以部分替代鱼粉蛋白,缓解饲料蛋白紧缺的现状。饲喂加入SCP 的饲料,会改善鱼、虾等肠道问题,提高成活率等[62]。还有一些专家直接采用成品的甲烷氧化菌生产的蛋白饲料,证实了甲烷氧化菌生产的单细胞蛋白应用在水产领域是安全无害的,并且具有一定优势。
3.2.1 改变肠道形态,降低死亡率
近年来,许多专家发现用甲烷氧化菌生产的SCP替代鱼粉饲料,会降低鱼、虾等水产动物的死亡率。经研究发现,给水产动物饲喂含有SCP 的饲料,可以提高肠道微绒毛高度、增加了肠道黏膜褶皱深度,从而改善了肠道菌群,进而提高了虾的抗病率。陈永康等[63]进行了12 d的副溶血性弧菌挑战,对白虾注射副溶血弧菌,饲喂SCP 与饲喂普通鱼粉蛋白的白虾作对比,发现用SCP替代15%的鱼料蛋白饲喂的白虾死亡率显著下降,对白虾的肠道进行分析,发现饲喂含有SCP 的饲料,改善了肠道微绒毛长度、增加了肠道黏膜褶皱高度。Jintasataporn 等[64]也通过饲喂试验发现,对白虾饲喂含有SCP 的饲料,会减少肝胰腺弧菌属的负荷,从而提高虾的存活率。张启乐等[65]用荚膜甲基球菌蛋白代替鱼粉对幼年大口鲈鱼进行饲喂,发现适量的荚膜甲基球菌蛋白可以提高肠壁总长度和肠壁绒毛长度,进而提高了鲈鱼的成活率。徐冰英等[66]直接采用荚膜甲基球菌蛋白替代基础饲料中的鱼粉蛋白来饲喂幼年黑鲷鱼,经过70 d 投喂试验,发现用荚膜甲基球菌蛋白替代24.80%的鱼粉蛋白,可以显著增加鱼的中肠微绒毛密度,改变肠道。梁伟兴等[67]用SCP 替代鱼粉蛋白饲喂养珍珠龙胆石斑鱼,在一定范围内,随着SCP 替代鱼粉蛋白含量增加,肠道绒毛高度呈变大趋势,肠道绒毛密集程度增加。
3.2.2 提高饲料转化率
试验表明,饲喂适量的SCP 饲料,可以提高水产动物的饲料转化率,这可能是饲喂甲烷氧化菌生产的SCP 可以提高部分酶的活性来帮助水产动物进食消化。Jintasataporn 等[64]饲喂SCP 的饲料和饲喂普通鱼粉饲料的白虾做对比,发现饲喂SCP 组的虾饲料转化率要远远高于普通饲料组;徐冰英等[66]发现用8.27%的荚膜甲基球菌蛋白替代鱼粉蛋白会增强黑鲷前肠的胰蛋白酶和中肠的淀粉酶活性,从而提高生长性能和饲料转化率;恒宇等[68]把荚膜甲基球菌蛋白添加到幼年鲤鱼的日粮中,发现在日粮中添加6%、8%和10%荚膜甲基球菌蛋白可改善鲤鱼的生长性能和饲料利用率。
3.2.3 改善肠炎,保护肠道
饲料中的豆粕蛋白容易引起鲑科鱼类患肠炎,它们对溶剂提取的豆粕特别敏感,主要是由豆粕中一种或多种醇溶性成分引起,而饲喂SCP 替代鱼粉的饲料可以改善鱼患肠炎的几率,这可能与甲烷氧化菌的细胞壁中的脂多糖有关。Romarheim 等[69]在饲喂大西洋鲑鱼的饲料中添加了荚膜甲基球菌细胞粉,证明了SCP 可以缓解因为饲喂豆粕引起的肠炎,起着保护肠道的作用。Romarheim 等[70]一项研究表明,SCP 可防止豆粕诱发的肠炎的发展,并且饲喂大西洋鲑鱼的饲料中含有SCP,可以更多地使用豆粕,而不会对肠道健康产生不利影响。Kleiveland 等[71]为了验证这一观点,用小鼠进行实验,在水中添加3.5%葡聚糖硫酸钠诱发小鼠患结肠炎,然后用SCP 饲喂小鼠,最后证实SCP 有可能通过增强结肠屏障功能来减弱葡聚糖硫酸钠诱导的结肠炎。
3.2.4 其他功能优势
添加一定量的SCP 替代鱼粉蛋白,可以一定程度上改变鱼肝脏对于脂类的合成。梁伟兴等[67]用SCP替代鱼粉蛋白饲喂珍珠龙胆石斑鱼,发现肝脏组织的空泡化面积显著增加,促进了肝脏组织对脂类物质的合成和积累,但肝细胞膜界限清晰,表明SCP 未对石斑鱼肝脏组织造成损伤。还有研究表明,随着饲料中SCP 水平的增加,抗炎基因的表达量增加,这证明了SCP有利于提高鱼类抵抗疾病的能力。张启乐等[65]发现关于炎性细胞因子基因的变化,增加荚膜甲基球菌蛋白代替鱼粉蛋白含量,提高了大口鲈鱼的Toll 样受体2(TLR2)、肿瘤坏死因子α(TNF-α)、白细胞介素-1β(IL-1β)、白细胞介素-1β(IL-10)和转化生长因子β(TGF-β)的表达水平,表明饲喂甲烷氧化菌生产的SCP的水产动物可以在一定程度上控制炎症和阻止肠炎。
不同的水产动物,对于SCP 的适应能力是有限的,在不影响鱼的生长性能和健康的前提下,可以确定添加多少SCP 替代鱼粉蛋白。徐冰英等[66]用荚膜甲基球菌蛋白替代24.80%的鱼粉蛋白,对黑鲷的生长无不良影响,而替代8.27%的,可提高黑鲷的生长性能和饲料利用率。Biswas 等[72]在不影响黄尾鱼的生长性能和饲料利用率的前提下,添加荚膜甲基球菌蛋白来替代30%以上的鱼粉蛋白。郑继昌等[73]验证了用SCP替代30%鱼粉蛋白,不会对大菱鲆幼鱼的生长性能和健康产生不良影响。
4 展望
甲烷氧化菌生产的SCP 作为水产饲料的应用前景广阔,有力推动水产养殖可持续发展。主要优点之一是甲烷氧化菌生产的SCP 蛋白含量高,且与鱼粉的必需氨基酸组成及含量相近,使其成为鱼粉的替代品;优点之二是甲烷氧化菌把甲烷作为唯一能量和碳源,不像其他微生物生产的SCP 易染沙门氏菌等其他致病菌,更加安全。然而目前甲烷氧化菌在生产及应用SCP 时也存在一些问题,生产时如何最大限度提高气液传质的效率,这是甲烷生物转化的最主要的挑战;在应用时,需要考虑不同水产物种对于单细胞蛋白的适应性,不建议以甲烷氧化菌生产的SCP 作为单一蛋白来源,因其高浓度的蛋白含量可能会降低水产动物的饲料利用率,从而对生长性能造成影响,因此,仍需从营养调控和遗传育种等多方面开展研究以消除高水平添加单细胞蛋白的不利影响。
目前国内对于甲烷氧化菌生产的单细胞蛋白饲料的应用还较少,一些商业化的产品也被应用在动物中,相信不久的将来应该就能看到甲烷氧化菌产的SCP出现在我国水产饲料行业。
