不同带宽“带状皆伐”淡竹林的叶片生理特性及生产力比较研究
2023-11-22王枕戈祖星兰王海霞施建敏
苏 田,梁 宽,王枕戈,祖星兰,孟 遂,王海霞,申 展,邹 娜,施建敏*
(1.江西农业大学 林学院,江西 南昌 330045;2.江西省竹子种质资源与利用重点实验室,江西 南昌 330045;3.江西省林业科学院,江西 南昌 330013)
【研究意义】竹林作为我国南方重要的经济林,具有生长快、效益高等特点,是我国竹乡经济发展与山区人民脱贫致富的重要支柱[1-2]。但是,目前以择伐、皆伐为主的采伐方式分别面临采伐成本高或竹林衰退、经营不可持续等问题[3-5]。为此,王龙凤等[6]基于竹类植物的克隆整合特性和采伐作业的便捷性,于2016年提出了竹林“带状皆伐”方式,旨在提高竹林经营可持续性、降低采伐人工成本。
【前人研究进展】“带状皆伐”方式是将待采伐林分划为等宽的若干条带,每次隔条皆伐,皆伐条带和不采伐条带分别称为“皆伐带”和“留竹带”,两类条带按采伐周期进行往复交替[6-7]。按照“带状皆伐”的设计,带状皆伐降低了采伐人工成本,而“留竹带”保证了相邻“皆伐带”竹子更新的养分需求,从而克服了皆伐和择伐的弊端,达到竹林高效、可持续经营目的[6-7]。梁宽[8]针对淡竹的“带状皆伐”研究发现,在5,10,20 m 3 种带宽的采伐实验中10 m 宽度的竹材产量和经济效益最佳。詹美春[9]在毛竹林开展的“带状皆伐”研究也同样发现带宽适中的8~12 m 竹林采伐后恢复状况最优。现有竹林“带状皆伐”研究主要集中在采伐方式优化、竹材产量以及生态效益等方面[7-11],而对于皆伐带宽适中的竹林产量形成生理机制研究尚未涉及。竹子是克隆植物,竹株间可以通过相连的地下竹鞭进行资源传输、共享,该过程称为克隆整合[12-13]。资源传输一般从植物的“源”到“汇”,“源”是资源供应方,“汇”是资源需求方[14]。传输距离是影响“汇”从“源”获取资源量的重要因素,距离越远则资源供给的能力越弱,同时外界干扰,如修剪等会改变原来的“源-汇”关系[15]。“带状皆伐”基于克隆整合特性设计,但对于不同皆伐带宽的产量差异尚未进行机理阐释[8-11]。众所周知,植物生物量高低由光合作用所驱动和决定[16-17]。叶片光合作用强弱除了通过净光合速率(Net photosynthetic rate,Pn)量化描述外,比叶面积(Specific leaf area,SLA)也是作为一个表征叶片光获取能力的功能性指标[18]。同时,光合作用也受叶片养分因素影响,有研究认为叶片氮(Nitrogen,N)、磷(Phosphorus,P)含量与光合速率显著正相关[19-20]。据此,不同带宽竹林“带状皆伐”导致的“源—汇”关系变化会在叶片生理特性差异上反映出来。
【本研究切入点】根据“源—汇”理论,“带状皆伐”留竹带是资源供给的“源”,而皆伐带是接受资源的“汇”。基于此,笔者认为“带状皆伐”改变了竹林养分供给的“源-汇”关系,从而影响叶片生理特性,最终在竹林生产力上形成差异。【拟解决的问题】为此,本研究以“源—汇”理论为指导,通过探究不同带宽留竹带和皆伐带的淡竹叶片生理特征、竹子生长和生物量等特征差异及其关系,为不同带宽采伐的竹林生产力提供理论解释,同时为竹林科学采伐提供理论依据。
1 试验材料与方法
1.1 试验材料
淡竹(Phyllostachys glauca)属禾本科刚竹属植物,茎竿高6~18 cm,胸径可达2~6 cm,是集食用、材用、药用、观赏为一体的多用途竹种。本研究区位于江西省西北部瑞昌市(29°23′~29°51′N,115°06′~115°44′E),区内淡竹资源丰富,天然淡竹林面积达5 600 hm2,多分布于石灰岩山地[21]。
1.2 试验方法
1.2.1 试验设计 采用随机区组设计在实验区选择长势良好且立竹度近似的淡竹林,设置5,10,20 m 3种带宽开展带状皆伐。采伐前对各带宽处理的林分进行了本底抽样调查(表1),结果显示不同处理的林分结构相近,无显著差异(P>0.05)。

表1 试验淡竹林本底特征Tab.1 Initial characteristic of Phyllostachys glauca forests
于2016 年12 月对待采伐林地沿山坡纵向划分等宽条带,隔行皆伐。带状皆伐实施后,在各带宽处理的留竹带和皆伐带分别设置5 m 长的实验调查样地,即3 种皆伐带宽处理有5 m×5 m、5 m×10 m 和5 m×20 m 3 种规格样地,样地数量分别为8 个、12 个、10 个。在采伐新竹长成后,于2017 年和2018 年分别对样地的新竹进行生长特征和叶片生理特征观测。
1.2.2 新竹生长特征测定 对各带宽处理下“皆伐带”和“留竹带”样地中分别在2017 年和2018 年对新竹进行每株检尺,测定其胸径(Diameter at breast height,DBH)、株高。立竹度(株∕m2)为样方内2017 和2018年新竹总数除以样方面积。
1.2.3 叶片生理指标测定 叶片光合作用试验在2018 年9 月晴天09:00—11:00 进行,每个样方中选5株淡竹,在每株冠层中部南面枝条上选取3枚正常的功能叶为试验对象,用LI-6400便携式光合仪(Li-Cor,美国)在红蓝光源下(光强1 500 mmol∕(m2·s)、相对湿度43%、温度30 ℃、流速500 mmol∕s)测定光合指标,包括净光合速率(Pn)、蒸腾速率(Transpiration rate,Tr)、水分利用效率(Water use efficiency,WUE)等,重复测定3次。
选取长势良好的竹株,在冠层上、中、下随机摘取40~50 枚完整叶片,用于测定比叶面积(SLA)。叶片面积用CI-203 便携式激光叶面积仪测定;将叶片放入烘箱105 ℃杀青30 min,后65 ℃烘48 h 至恒重,并称量得到叶片生物量。将烘干叶片粉碎过100目筛,采用浓硫酸-双氧水进行消煮,制备待测样液,利用流动分析仪(SEAL Analytical Chem.Tray AA3)测定叶片的氮、磷含量[19]。
1.2.4 指标计算
式3中,生物量为带状皆伐后样地内新长竹子(2017和2018年竹子)的单株生物量总和,单株生物量根据邹凯等[22]的淡竹生物量一元二次函数模型进行计算。本研究以采伐后的新竹林生物量作为竹林生产力的表征指标。
1.3 统计分析
采用Excel 2010和SPSS 17.0进行整理分析。利用单因素方差分析比较净光合速率、蒸腾速率、水分利用率、比叶面积、叶片氮磷养分、新竹胸径、株高、生物量等指标在不同带宽处理间差异,利用Pearson相关性分析检验上述指标之间的相关关系。利用双因素方差分析确定影响采伐处理、采伐宽度及其交互作用对上述指标的影响,使用Origin 8.0 软件作图。利用Canoco 5 进行竹林生产力与生理特性的RDA分析并作图。
2 研究结果
2.1 叶片光合特性
从表2可看出:随着带宽增加,留竹带竹子的净光合速率(Pn)逐渐下降,20 m 带宽Pn比5 m 带宽的降低2.5 µmol CO2m2∕s;水分利用效率(WUE)以5 m带宽值最高;但是蒸腾速率(Tr)和比叶面积(SLA)以10 m带宽最高。与留竹带叶片光合特性随带宽变化趋势有所不同,皆伐带的竹子Pn和Tr均以10 m带宽最高,但WUE 仍以5 m 带宽最高;其中10 m 带宽的WUE 显著小于5 m 带宽(P<0.05),而SLA 随着带宽增加逐渐降低。从不同带宽的整体数据(同带宽留竹带与皆伐带平均值,下同)来看,其随皆伐带宽的变化趋势与留竹带的一致。
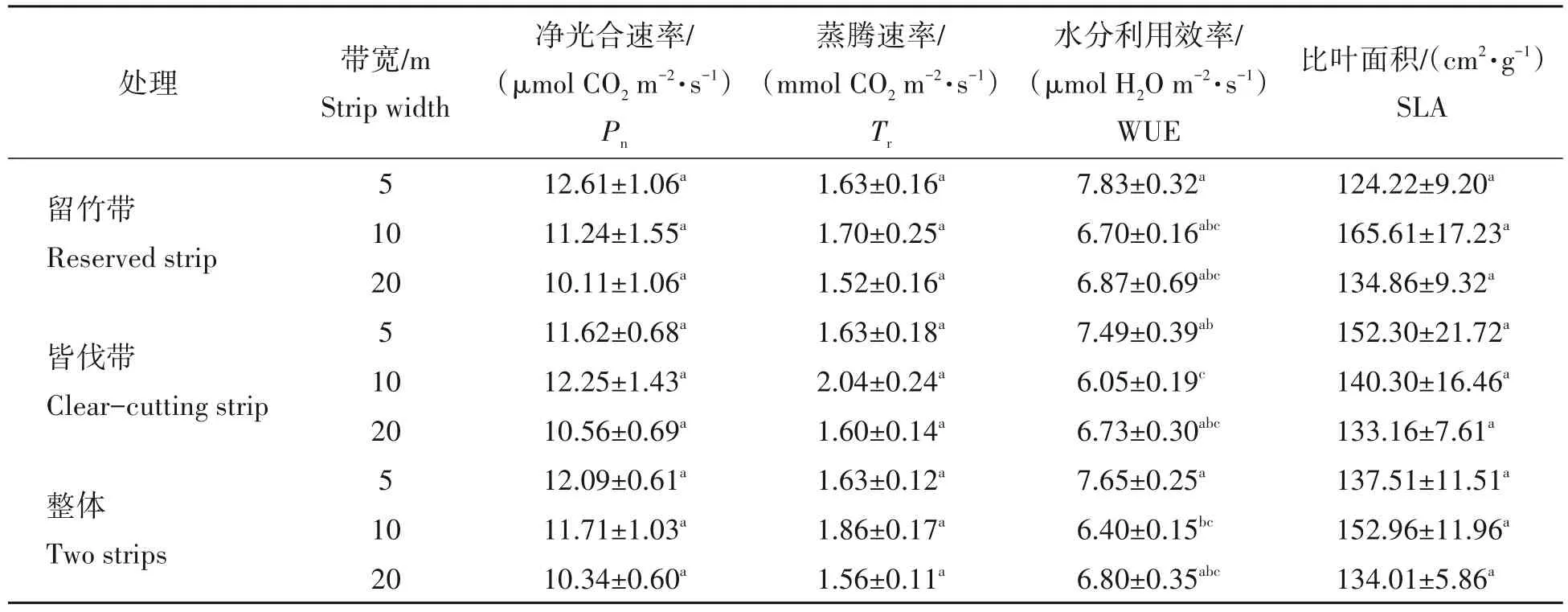
表2 带状皆伐处理下不同带宽之间叶片光合生理特征Tab.2 Leaf photosynthetic traits from different strip-width stands after strip clear-cutting
相同带宽的留竹带和皆伐带叶片光合特性比较发现,10 m 和20 m 带宽的皆伐带Pn、Tr高于留竹带,而10 m 带宽的皆伐带WUE 显著低于留竹带(P<0.05);对于SLA 值,5 m 的带宽皆伐带高于留竹带,10 m 带宽留竹带高于皆伐带。总体上,3 种带宽的皆伐带与留竹带叶片光合特性差异以10 m 带宽的最大。
2.2 叶片养分特性
随着带宽的增加,留竹带和皆伐带叶片氮(N)和磷(P)的含量未呈现出一致的变化规律(图1)。留竹带中,10 m带宽叶片N含量最低((20.65±0.38)mg∕g),相比5 m和20 m显著降低12.37%和11.76%;5 m带宽叶片P 含量最高((1.70±0.08)mg∕g),相比10 m 和20 m 带宽显著提高26.91%和47.46%。皆伐带中,10 m 带宽N 含量最高((23.22±0.55)mg∕g),相比5 m 和20 m 带宽分别提升11.45%和12.80%(P<0.05);但叶片P含量与N含量关系相反,以10 m带宽最低(图1)。整体上,叶片N含量在不同带宽间无明显差异,叶片P含量5 m带宽最高,相比20 m带宽显著提高21.71%(P<0.05)。

图1 带状皆伐处理下不同带宽淡竹叶片养分含量比较Fig.1 Nitrogen(A)and phosphorus(B)contents of leaves of different strip widths under strip clear-cutting
相同带宽的留竹带与皆伐带叶片养分含量比较发现,叶片N 含量在5 m 和20 m 带宽的留竹带高于皆伐带,在10 m带宽留竹带低于皆伐带(图1A,P<0.05);叶片P含量在5 m带宽的留竹带高于皆伐带(图1B,P<0.05)。从皆伐带与留竹带的竹子叶片养分特性差异来看,5 m带宽的差别大于10 m和20 m带宽。
2.3 竹林生长特征
随带宽逐渐增加,胸径(DBH)和株高降低,总生物量先增后降,立竹度在留竹带呈先增后降,而在皆伐带和整体呈增加趋势(图2)。留竹带竹子DBH 和株高以5 m 带宽最高,10 m 带宽次之,相比20 m 带宽显著增加(DBH 增加39.90%、31.03%,株高增加36.63%、28.48%,P<0.05);立竹度以5 m 带宽的最低;生物量以10 m 带宽的最高,相比20 m 带宽显著增加77.80%。皆伐带竹子不同带宽的DBH、株高和生物量数据规律与留竹带规律相似,立竹度以5 m带宽的最低,且显著低于10 m和20 m带宽(P<0.05)。不同带宽的整体数据随带宽变化规律与留竹带和伐竹带的规律基本一致。
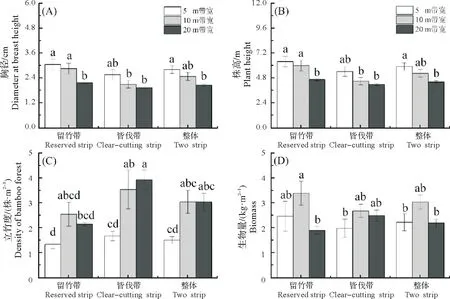
图2 不同带宽皆伐后淡竹更新生长特征Fig.2 Bamboos growth characteristics between different strip widths after strip-clear cutting
相同带宽的留竹带和伐竹带新竹生长特性来看,5,10,20 m带宽的DBH和株高在留竹带大于皆伐带;立竹度在3种带宽下均呈现皆伐带大于留竹带;生物量在5 m和10 m带宽的留竹带大于皆伐带,20 m带宽规律相反。整体上,除立竹度外,10 m带宽新竹生长在留竹带和伐竹带的差异大于5 m和20 m带宽。
2.4 采伐处理综合分析
双因素方差分析发现,采伐处理(保留、皆伐)对叶片氮含量、胸径、株高和立竹度存在显著影响,采伐带宽对水分利用效率、叶片磷含量、胸径、株高、立竹度和生物量有显著影响,采伐处理和采伐带宽的交互作用对叶片氮、磷含量具有显著影响(表3)。由分析可知,采伐处理和采伐带宽均会对竹子的生理特性和生产力产生显著影响,而且两者的交互作用显著影响叶片养分。
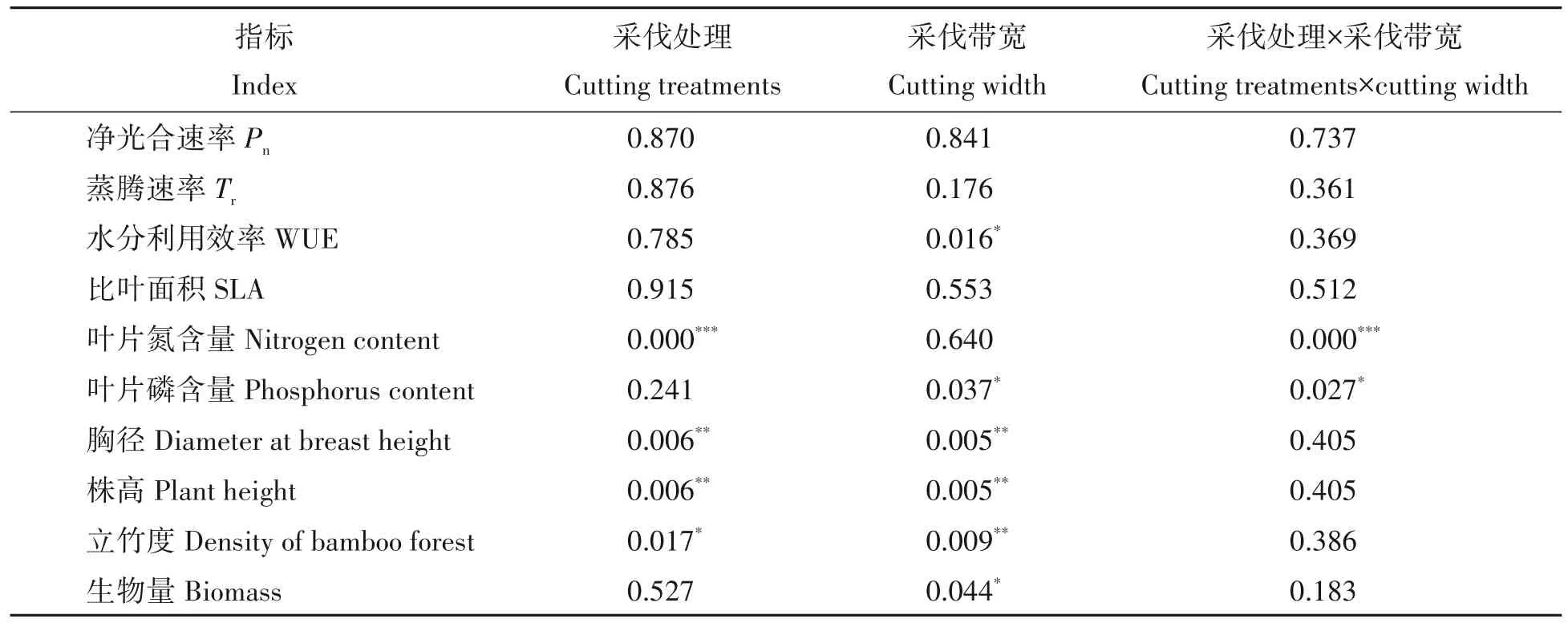
表3 带状皆伐方式对叶片生理特征和竹林生产力的双因素方差分析Tab.3 Two-way ANOVA shows the effect of strip clear-cutting on leaf physiological traits and productivity
运用RDA分析留竹带和皆伐带竹子叶片生理特性、竹林结构和生产力的关系,结果显示:(1)留竹带新竹个体大小(胸径和株高)与皆伐带宽、光合速率、蒸腾速率负相关,叶片养分(磷、氮)对新竹大小有一定正影响,竹林生产力(生物量)则与立竹度有一定正相关(图3A);(2)皆伐带新竹个体大小主要受比叶面积和水分利用效率的正影响,同时一定程度上受净光合速率和蒸腾速率负影响;叶片养分与立竹度则对竹林生产力有正向影响(图3B)。从RDA 分析结果可知,留竹带和皆伐带的新竹生长影响因素既有相同也有差别之处:相同的是两类采伐处理的新竹个体生长均受带宽的负影响,即皆伐带越宽两类新竹均个体变小;同时两类采伐处理的竹林生产力均受立竹度的影响,立竹度越大竹林生产力越高。不同的是比叶面积与留竹带新竹个体大小负相关,而与皆伐带新竹个体大小正相关;叶片养分与竹林生产力的关系在皆伐带比留竹带密切。
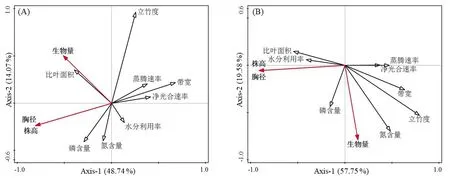
图3 留竹带(A)和皆伐带(B)竹林生产力与叶片生理特征、采伐带宽和竹林结构的RDA分析Fig.3 RDA analysis of bamboo forest productivity with cutting strip width and leaf physiological characteristics.(A)Reserved strip;(B)Clear cutting strip
3 讨论
3.1 留竹带与皆伐带的叶片生理特性和竹林生产力差异
“源-汇”假说认为,克隆植物在分株间会沿着“源-汇”梯度进行光合产物和营养物质从“源”到“汇”的转移[23],转移程度和速率取决于“汇”的需求和“源”的容量[24]。前人已研究发现相连的分株中,幼株是强“汇”,它从成年“源”株获取足够资源以满足生长所需[25];对于成年分株之间,如果受外部干扰如修剪或采伐等,其“源-汇”梯度发生改变,会显著增加光合产物或营养物质向受干扰的“汇”株转移[26-27]。
本研究中淡竹林经带状皆伐处理后,皆伐带作为强“汇”需要从留竹带“源”获取资源以保证皆伐后的竹林生长更新,所以皆伐带和留竹带竹子叶片生理特性和竹林生长特征的差异是对该“源—汇”梯度的响应。研究结果显示,留竹带和伐竹带竹子的光合速率差异因带宽而异,5 m带宽留竹带高于皆伐带,而10 m 和20 m 带宽留竹带低于皆伐带(表2)。10 m 和20 m 带宽下皆伐带淡竹的高净光合速率和高比叶面积,可能是皆伐后新生竹子适应生长的调节机制,因为除了从留竹带“源”处获取资源外,仍需自身加强光合作用提高生物量积累以加速竹林恢复[28-30]。
研究发现不同皆伐宽度的留竹带和皆伐带竹子叶片养分含量未有一致性变化规律,多数情况下留竹带的竹子叶片N 和P 含量高于皆伐带,但在10 m 带皆伐带宽叶片N 和20 m 带宽皆伐带叶片P 高于同宽度的留竹带(图1B);而从留竹带和皆伐带的3 种皆伐宽度平均值来看(N 含量为(22.32±0.83)g∕kg 和(21.55±0.84)g∕kg,P 含量为(1.39±0.16)g∕kg 和(1.31±0.04)g∕kg),它们N 和P 含量相近,没有显著差别(P=0.69 和P=0.66)。苏文会等[11]在毛竹带状采伐研究中发现,采伐带新竹各器官的非结构性碳(NSC)低于对照的保留带竹子NSC,并认为新竹生长的养分来自于保留带母竹是NSC 含量低的主要原因。本研究结果与此类似,皆伐带竹子养分多低于留竹带,但10 m 和20 m 带皆伐带叶片养分反而高于留竹带,这可能是本研究的新竹包括了2个年度的新生竹,经过两年适应性生长后皆伐带竹子加强了根系养分吸收,较大程度减少了对留竹带养分供应的依赖[25,31]。
采伐处理对新竹的大小具有显著影响(表3),留竹带的新竹胸径、株高和生物量等生长特征普遍优于皆伐带(图2),这与张洋洋等[10]和曾宪礼等[32]在毛竹带状采伐中得到的结果一致,他们也发现皆伐带内的新竹胸径普遍低于保留带。对于皆伐带竹子质量弱于留竹带的原因,一方面是因为皆伐带新竹数量多,从留竹带“源”株获取资源分配至各新竹的量有限,难以满足竹株的优势生长[33];另一方面可能是留竹带新竹离“源”更近,获得的生长支持大于皆伐带新竹,进而提高了单株大小。
3.2 带状皆伐宽度对叶片生理特性和竹林生产力影响
竹类植物属于克隆植物,克隆整合带来的资源共享为植物生长、繁殖,特别是应对不利环境提供了生理基础[12]。现有研究认为克隆分株间的克隆整合存在距离效应,“汇”株离“源”株距离越远,其获得的资源量越少[15]。本研究发现皆伐宽度与叶片的生理特性无明显关系,但对竹株生长有显著负影响,随着带宽逐渐增加留竹带和皆伐带竹子胸径、株高下降(表2、3)。詹美春[9]对毛竹开展“带状皆伐”研究也发现,随着皆伐宽度增加,新竹胸径、株高均下降,即采伐宽度过大会导致植株生长受到抑制。这与本研究发现的皆伐宽度过大造成淡竹林生产力下降的结果一致[9]。克隆整合特性进行物质传输的有效性会随着距离的增加而减弱[15],这是导致新竹生长随着采伐带宽增加而被抑制的主要原因。
对于淡竹的克隆整合有效距离,同位素实验和野外采伐观测均认为最远可达5 m[6,12],所以在带状皆伐方式下,皆伐带左右两侧的养分有效整合距离在10 m左右。本研究中,皆伐带竹子呈现净光合速率随采伐宽度的增加先增后降趋势,以10 m 带宽最高。前人发现净光合速率随比叶面积增加而增加[34-36],这与皆伐带中10 m 带宽比5 m 和20 m 带宽比叶面积更高,净光合速率增加的结果相印证。比叶面积在留竹带和皆伐带均呈现先增加后降低的趋势,10 m 和20 m 带宽之间差异显著。Cornelissen 等[34]发现,比叶面积越大,越有利于植物捕捉光源,会增加净光合速率,适度的干扰(5 m 和10 m 宽度)促进植物叶片功能性适应,提高资源累积,加速林分恢复。在生产力方面,留竹带和皆伐带的新竹大小均随皆伐距离增大而降低,而竹林立竹度大体上呈升高趋势,这使得最高10 m 带宽的生产力均最高(图2)。这与詹美春等[9]研究发现,过大的皆伐带宽会导致留竹带和皆伐带新竹胸径、株高和生物量下降结果一致。
4 结论
淡竹林进行不同带宽“带状皆伐”后,作为“源”的留竹带和作为“汇”的皆伐带在竹子叶片光合、养分生理特性、新竹大小特征和生产力上均有明显差异,同时皆伐宽度对留竹带和皆伐带的叶片生理、新竹大小和生产力亦有显著影响。20 m皆伐带宽超出了淡竹克隆整合有效距离,虽然新竹数量多但个体小,5 m 皆伐带宽虽然新竹个体大但萌发的新竹数量少,而10 m 皆伐带宽的新竹大小和立竹度均适中,其年生产力最高。
分析认为,10 m 带宽在淡竹最大有效整合距离范围内,留竹带的“源”供应能较好满足新竹生长需求,同时该带宽皆伐刺激了更多新竹萌发,并且新竹又通过提升竹子光合生理特性和养分生理指标进行适应生长,这些应该是10 m 带宽生产力高的主要原因。不同带宽“带状皆伐”叶片生理和生产力的比较研究结果为竹林高效采伐提供了理论参考。
