杉木家系苗期性状的遗传变异、相关性及综合评价研究
2023-11-22朱科帆
曾 路,朱科帆,安 波,朱 勇,张 芸*
(1.江西农业大学 林学院∕国家林业与草原局鄱阳湖流域森林生态系统保护与修复重点实验室,江西 南昌 330045;2.江西省信丰县林木良种繁育中心,江西 信丰 341600)
【研究意义】杉木(Cunninghamia lanceolata),是我国特有的速生用材树种,其遗传改良研究一直倍受重视,已在中心产区完成了第三代育种[1],筛选出了大批优良种源和优良家系应用于生产造林,江西省在杉木的优良种质选育工作相对滞后,在生产上能推广的优良种质相对不足[2]。林木育种普遍存在世代间隔长、育种周期较长和遗传改良过程进展缓慢等问题,苗期选择具有降低田间试验费用、缩短育种周期、加快遗传改良进程等优势[3],研究[4]表明杉木大多数苗期生长性状能稳定遗传,性状早晚的相关性明显。因此,开展赣南地区的杉木家系苗期生长监测、遗传变异分析和科学评价,为挖掘适应赣南地区的优良造林家系提供材料,对加速江西特色优良新品种创制的步伐具有现实意义。【前人研究进展】国内外学者采用苗期表型测定、全基因组选择和田间长时间观测相结合的方法研究林木种内性状及遗传变异特征,表明性状表型的早期选择在提高林木选育效率、缩短世代间隔、提高林木的遗传增益上仍具有显著效果。经过子代遗传测定,不仅能够从亲本中进行后向选择,而且还能从优良家系中挑选出最佳单株,更好管理和改善现有种子园,为高世代遗传改良提供良好的育种材料[5]。【本研究切入点】苗高、地径、材积等地上部生长性状直接体现用材树种的生长状况,目前也主要将地上生长性状作为早期选择优良家系的衡量指标[6]。根系是植物水分和养分的吸收和转运的主要器官,直接参与协调植株地上部分与土壤环境之间的交互,其性状不仅反映了植物对土壤资源探索能力及对逆境的适应性[7],对地上部分表现和产量也有重要指示作用,根系性状作为选择指标在育种中应用对选育高产、抗逆性强的品种有实际应用价值[8-9]。由于根系表型分析的困难以及对根系性状遗传学的理解有限,根系性状在林木品种选育中并没有被广泛应用。研究发现不同种源或家系杉木在南方红壤低磷环境下表现生长上的差异[10],优良的杉木品种可通过调控根系生长[11]、提高有机酸分泌[12]、协调根系菌根真菌[13]等适应机制提高养分利用效率,因此将根系性状纳入优良家系筛选的评价体系,对选育丰产、低投入、高效率的多功能优质杉木品种具有一定意义,有望解决林业上高产、稳产、高效之间的矛盾。【拟解决的关键问题】本研究以江西省信丰县林木良种繁育中心选育的15个1年生杉木家系为试验材料,对8个地上部表型性状、7个叶片性状和8个根系性状进行测量,比较不同杉木家系苗木差异性及相关性,估算性状的遗传参数和变异情况,通过聚类分析和隶属函数分析对15 个杉木家系的表型性状进行了评价鉴定,为赣南地区杉木营建高品质的高世代种子园提供优质遗传材料提供参考资料。
1 材料与方法
1.1 试验材料
试验材料为江西省赣州市信丰县林木良种场国家杉木良种基地(25°38′N,114°93′E)的15 个家系1 年生杉木实生苗,种子分别来自信丰县林木良种场杉木2 代种子园(混系)、92 年全同胞子代林2 代种子园(3×5、7×8、7×12、8×10、11×12、13×18、20×23、20×24)、安远县森源科技林木良种场杉木1 代种子园(sy18、sy21、sy22、sy27)、贵州种源(天柱18)以及广西种源(广西那么3)。
试验家系于2020 年12 月由林场播种,苗圃整地作床,宽1.0~1.2 m,高25 cm,苗床间留有25 cm 步道。播种前用0.5%的高锰酸钾溶液浸种消毒,成块播种育苗,期间追肥两次,有效成分45%(N-P-K:15-15-15)。试验设计采用完全随机区组设计,组内重复5次,每个重复10株。
1.2 调查性状
本研究共调查21 个性状,分别为X1=苗高(cm),X2=地径(cm),X3=冠幅(cm),X4=坐生密度(片),X5=一级侧枝数(个),X6=枝盘数(个),X7=当年生节间距(cm),X8=地上生物量(g),X9=叶片面积(cm2),X10=叶片体积(cm3),X11=叶片干重(g),X12=比叶面积(cm2∕g),X13=叶片含水量(%),X14=叶片组织密度(g∕cm3),X15=总根长(cm),X16=根直径(mm),X17=根体积(cm3),X18=根系生物量(g),X19=比根长(cm∕g),X20=比根表面积(cm∕g),X21=根组织密度(g∕cm3)。其中X1—X8 为地上部生长性状,X9—X14为叶片性状,X15—X21为根系性状。
1.3 生长性状的测定
2021 年11 月对15 个家系1 年生苗木进行地上表型性状测定。利用钢卷尺(精确度0.01 cm)及游标卡尺(精确度0.01 cm)对15 个家系苗木的苗高(cm)、地径(cm)和冠幅(cm)进行测量,每个家系调查80 株苗,同时每个家系随机选取20株幼苗测定一级侧枝数(个)、枝盘数(个)、当年生节间距(cm)和坐生密度(片)[14]。随后各家系选取10~15株带回实验室测定叶片性状和根系形态指标。每株选取10片叶片用扫描仪(V700,Seiko Epson Co.,Nagano,Japan)扫描,使用WinRhizo 软件(Pro 2007d,Regent Instruments Inc.,Canada)获得叶面积(cm2)、叶体积(cm3);用游标卡尺(精确度0.01 cm)在叶片上部、中部、下部依次进行测量读数,取平均值作为叶片实际厚度(mm)。电子天平测量叶片鲜重(g),并将叶片放入烘箱中100 ℃杀青,60 ℃烘干至恒重,为叶片干重(g)。叶片含水量(%)、比叶面积(cm2∕g)、叶片组织密度(g∕cm3)分别用公式(1)~(3)计算。
称取单株地上部分和根系鲜重(g),随即在60 ℃下烘干至恒重,并分别称量地上部分和根系的干重(精确度0.01 g)。使用WinRhizo软件分析根系形态参数:总根长(cm)、根直径(cm)、根体积(cm3)和根表面积(cm2)。比根长、比根表面积和根系组织密度分别通过下式计算:
1.4 数据处理与统计分析
采用Excel2021 软件进行数据整理。运用SPSS 的描述性统计功能计算苗期表型的均值、变异系数、最大值、最小值等基础数据,采用单因素方差分析法(One-way ANOVA)检验家系之间差异的显著性,采用Duncan 多重比较比较不同家系各个性状平均值的差异;运用SPSS 计算地上生长性状、根系和叶片性状指标之间的Pearson 相关性,采用系统聚类法进行聚类分析。利用Origin2021 软件绘制相关性热图和遗传增益柱状图。
遗传参数表型变异系数(PVC)[15]、家系遗传力(H2)[16]、现实增益(ΔGr)、遗传增益(ΔGg)[17]分别通过下式计算:
式(8)中:Mf为家系均方,Me为环境方差。
用隶属函数法对家系性状进行综合评价[18],公式为:
式(11)、(12)中,R(Xij)是i家系j性状的隶属函数值,Xij是i家系j性状的数值,Xmax、Xmin、分别是j性状中最大值、最小值和平均值,D为综合评价值,ω(j)是i家系j性状的权重,S(Xj)是j性状的数值标准差。
2 结果与分析
2.1 性状的遗传变异分析
方差分析结果(表1)显示,除X4(坐生密度)、X9(叶片面积)、X11(叶片干重)、X12(比叶面积)、X16(根直径)外,其他地上部生长性状和叶片性状在家系间均呈现极显著差异(P<0.05)。根系性状中除平均根径外,各性状在杉木家系间均存在显著差异(P<0.05),表明15 个家系存在显著的遗传和表型变异,具有较大的选择潜力。

表1 15个杉木家系各性状遗传变异参数分析Tab.1 Genetic variation parameters of different traits in 15 Chinese fir families
15 个家系各性状遗传变异系数差异较大,变化范围为2.86%~65.06%,其中叶含水量最低,遗传稳定性高,根直径、地上生物量的变异系数超过60%。各指标遗传力变化范围为0.07~0.96,各性状指标具有较高的遗传力,除X9(叶片面积)、X11(叶片干重)、X12(比叶面积)、X16(根直径)外,均超过0.40。
家系7×8的苗高、冠幅、当年生节间距的平均值最大,地上部分生物量位列第二位,具有速生丰产的潜力,而sy27和天柱18的苗高、地径、冠幅、当年生节间距、地上生物量远低于各指标的平均水平(图1)。叶表面积、叶体积、叶含水量、叶组织密度、比叶面积平均值最大家系为7×8、20×24、20×23、sy18、3×5(表2);最小的家系为sy18、sy22(sy18)、11×12、20×24、sy21;各家系间叶干重平均值无明显差异。根系部分性状变幅较大,其中广西那么3在15个家系中总根长、根表面积、根体积的平均值最小,各家系间性状表现差异明显(表3)。

图1 15个杉木家系地上生长性状Fig.1 Aboveground growth traits of 15 Chinese fir families

表2 15个杉木家系叶片性状均值Tab.2 The mean value of leaf traits of 15 Chinese fir families
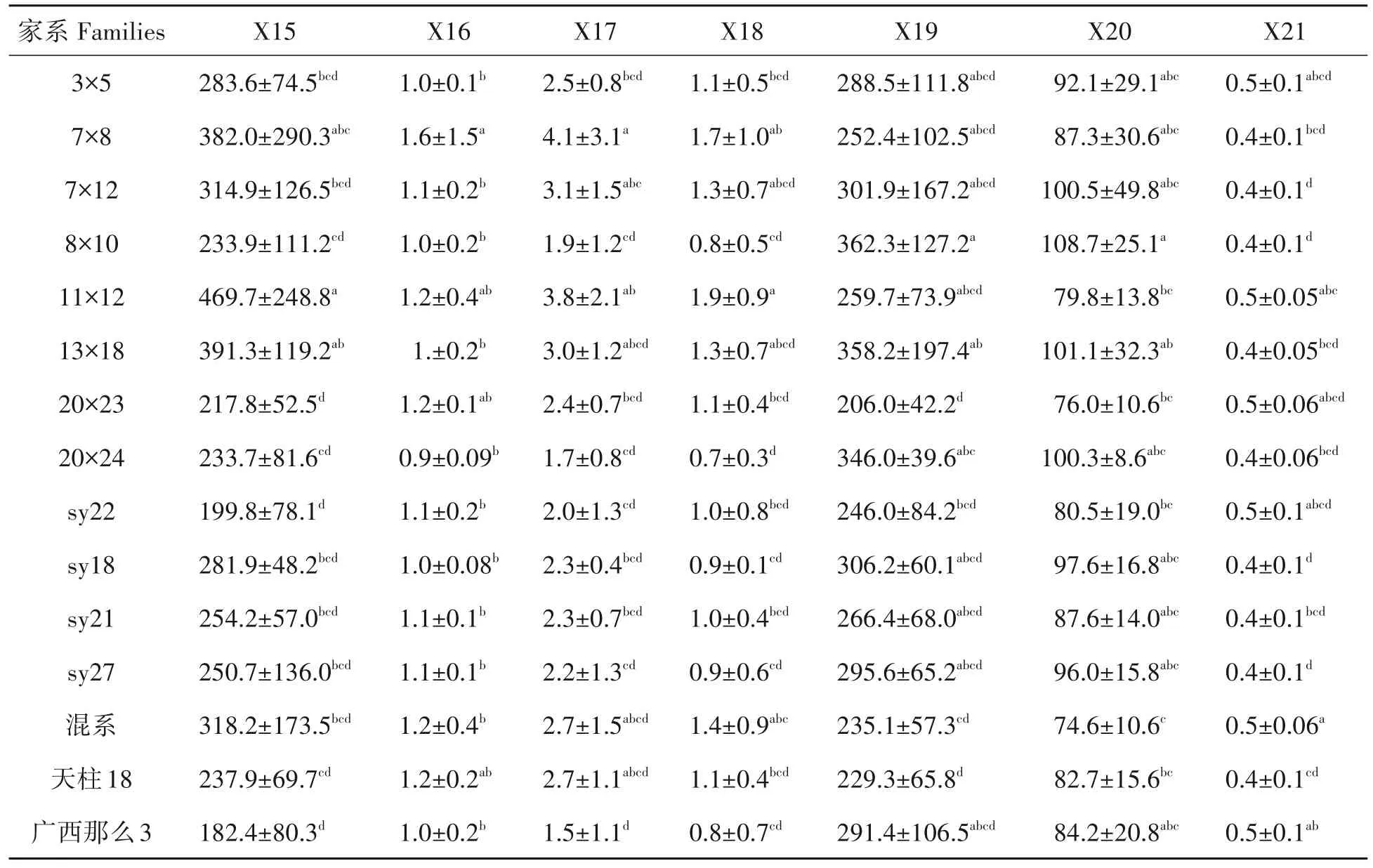
表3 15个杉木家系根系性状均值Tab.3 The mean value of root characters of 15 Chinese fir families
2.2 性状的相关性分析
大多数地上部分性状之间的相关性呈极显著相关性,苗高、地径、冠幅、坐生密度、一级侧枝数、地上生物量两两之间呈显著或极显著的正相关(P<0.01)(图2)。地上生长性状中仅地径、冠幅和地上生物量与超过4个以上的叶片性状呈显著的相关关系,其中地上生物量与叶片面积、叶片体积、叶片干重之间存在显著正相关(P<0.01),与比叶面积、叶片含水量呈显著的负相关关系(P<0.05)。根系性状与除当年生节间距外的大部分地上部生长性状均呈显著的相关关系(P<0.05),其中根长、根直径、根体积、根生物量、根组织密度与苗高、地径、冠幅、一级侧枝数、地上生物量呈极显著正相关(P<0.001),比根长和比根表面积与地上性状呈负相关关系,地上部生长性状与根性状的相关系数普遍低于与叶性状之间的相关系数,说明杉木根系性状与地上生长存在紧密关系。

图2 各性状的相关性热图Fig.2 Correlation heatmap of all traits
2.3 综合评价与优良家系的选择
由于叶片性状的变异系数较低,因此以变异系数大于50%和遗传力大于0.5 较大的地上生长和根系性状为评价指标:苗高、地径、冠幅、当年生节间距、地上生物量、总根长、根体积、根系生物量共8 个性状作为评价指标。由图3 可知,当欧式距离达到15 时,可将15 个家系划分为3 个类:其中第Ⅰ类包含sy21、天柱18、8×10等11个家系;第 Ⅱ 类则包含7×8、13×18、混系等3个家系;第Ⅲ类则包含11×12 1个家系。第Ⅰ类家系8个性状的现实增益和遗传增益均为负值,因此将其进一步归为苗期慢生家系(图4);第Ⅱ类家系除苗高冠幅的现实增益和遗传增益为负值,其余14 个性状的现实增益为2.31%~29.64%,遗传增益为2.01%~21.64%,为苗期速生家系;第Ⅲ类家系苗高、地径、冠幅的现实增益和遗传增益为负数,而当年生节间距、地上生物量、根长、根体积和根系生物量的现实增益和遗传增益在2.42%~65.42%,为速生中等家系。
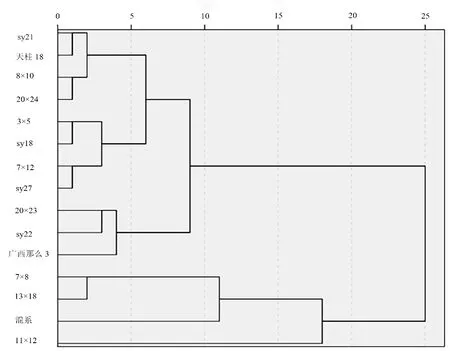
图3 15个杉木家系表型性状的聚类分析Fig.3 Cluster map of phenotypic traits of 15 Chinese fir families
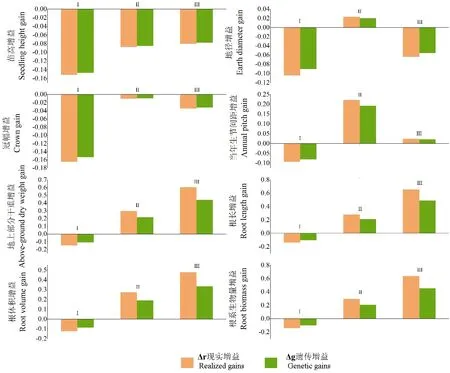
图4 3类杉木家系的现实增益和遗传增益Fig.4 Realistic gain and genetic gain of three types of Chinese fir families
以苗高、地径、冠幅、当年生节间距、地上生物量、总根长、根体积、根系生物量作为评价指标,通过加权隶属函数法对15 个杉木家系的表型生长性状进行综合评价,总隶属函数值按从小到大的顺序进行排列(表4)。在入选率为20%的情况下,3个家系11×12、7×8、13×18 入选,它们的地径、冠幅、地上生物量、总根长、根体积、根生物量平均值分别为0.58 cm、24.87 cm、7.66 g、414.34 cm、3.61 cm3、1.62 g,现实增益分别为2.42%、5.67%、46.29%、45.92%、41.53%、42.22%,遗传增益分别为2.08%、5.27%、29.16%、30.77%、23.67%、24.91%。

表4 15个杉木家系21个性状的隶属函数值分析Tab.4 Membership function analysis of 21 characters in 15 Chinese fir families
3 讨论与结论
遗传力体现性状遗传的稳定程度,高遗传力说明受环境影响小,评价选择的效果更好[19],根据遗传力大小大致划分遗传控制程度:大于0.5 为高遗传力;0.2~0.5 为中等遗传力;小于0.2 为低遗传力。本研究1 年生杉木幼苗地上生长性状的家系遗传力为0.49~0.96,除坐生密度(0.49)外,其余地上生长性状属于高遗传力,其中苗高、冠幅,其遗传力达到了0.9 以上,其他研究也发现苗期和幼龄林杉木的树高和地径具有中等以上的遗传力[1,4,20],表明地上性状受到强烈的遗传控制,是杉木优良家系评价选择的重要依据。根叶家系遗传力,低于地上生长性状,在0.07~0.75,其中叶片体积、叶片含水量、叶片组织密度、总根长、根体积、根系生物量和根组织密度的遗传高于0.5,说明这些性状的遗传稳定程度高,具有评价和选择的潜力,值得进一步研究。
变异系数大小可反映群体的变异程度,是衡量性状遗传变异潜力的有效指标[21]。本研究中15 个1 年生杉木家系各表型性状的变异系数变化范围为2.86%~65.06%,大部分性状在不同家系间存在显著的差异(P<0.05),表明杉木生长和性状分化明显,变异较丰富,具有较高的遗传改良潜力,这与杉木在我国有广泛的分布区这一事实相符,长期的自然选择造成杉木在种源间、林分间、个体间甚至个体内存在遗传变异。在3 类性状中,地上部分生长性状(13.45%~65.06%)和根系性状(19.38%~60.12%)的变异系数较大,表现出较大的表型可塑性,叶片性状的变异系数为2.86%~20.93%,其中叶干重、比叶面积、叶表面积3 个叶片性状在家系间无显著差异,表现出内稳定性。庾庐山等[22]研究发现33 个杉木无性系叶片变异叶片的变异系数变化范围为2.88%~17.92%,何智英等[23]采集所有杉木自然分布区的叶片和其他组织器官,发现叶片厚度的变异系数为5.1%,在所有研究的20 个性状中最低,叶片宽度的变幅也仅为10.5%,结合本研究结果和前人探究说明杉木叶片性状在遗传变异上较为稳定,且受到环境调控的影响较小。
叶片是植物光合作用的主要场所,其表面积及持绿时间影响植物的光合作用效率[24],而根系作为林木养分和水分的吸收器官,其性状(如根系生物量、形态和空间构型)反映植物对土壤资源的利用策略和竞争能力[7],植物生长很大程度取决于叶片和根系发育。植物在应对环境变化时并非调节某个单一性状,而是通过各性状间的协同与权衡作用来适应环境和生长变化[25],所以在林木的育种策略和遗传选择上,应做到多类型性状综合评价,以提高选择优良家系的几率。本研究对15个1年生杉木家系21个表型性状进行相关性分析发现:7个根系性状与地上生物量、树高和地径等地上生长指标呈极显著相关性,高于叶片性状与地上生长指标之间的相关性,说明根系的功能在杉木早期生长中起到更重要的作用,由于叶片性状的变异相对稳定,在杉木的早期选择中可以将根系性状作为重要的参考指标。另一方面,地上生物量与根系直径呈极显著正比,与比根长和比表面积表现出显著负相关关系,这一结果与杉木的根系功能属性相符。根据细根经济谱理论,与资源进取型树种相比,杉木通过增加根系生物量、提高根系直径和维持根系寿命去获取养分和水分[11,26],表现出低比根长和高组织密度[27-28],研究结果也为速生型杉木家系的早期选择提供了依据,即在生长初期,粗壮的根系有利于土壤水肥的吸收,可支撑杉木的快速生长,相同根系干重量情况下,应倾向于选择粗壮的根系而不是细长的根系。
遗传增益受到遗传力、变异系数的影响,是反映林木遗传改良效果的直接指标[21]。本研究中苗高、地径、冠幅、当年生节间距、地上生物量、总根长、根体积和根系生物量均受较强的遗传控制,变异系数处于较高水平,各性状间相关性均达到了极显著水平,因此选择以上指标可作为优良家系评价和筛选的参考因子。结合聚类分析和隶属函数综合评价法,将15个杉木家系分为苗期速生家系、速生中等家系和慢生家系3 个群体,筛选出3 个优良家系,遗传增益较高,选择效果较为显著。然而隶属于较速生类群的11×12入选,可能是因为该家系地上生物量和总根长较大,促使隶属函数值高从而入选。因此,在苗期选择时,也可适当考虑中等速生家系。此外,在家系综合评价中发现,遗传增益值较高的Ⅱ、Ⅲ类群的家系均来自于本地种子园,广西那么3、天柱18排名靠后,由此可见,本地种源的家系相较于外源家系在生长上占较大优势。
本研究通过聚类分析和加权隶属函数法将15 个杉木家系分为3 类,并初选出3 个优良家系,入选家系在地径、生物量、冠幅等方面均可获得正向增益,但考虑到林木生长周期长,生长受到遗传和环境的共同调控,在不同发育阶段林木的遗传和表型参数是动态变化的[17],基于苗期的优良家系选择存在一定的漏选和错选风险。饶显生等[4]研究发现杉木无性系大多数苗期高度、地径和材积等生长性状能稳定遗传,1 年生苗木与6 年生幼龄林性状之间的早晚相关性明显,说明通过苗期评价来进行杉木优良家系筛选具有一定的可靠性,但考虑到后期应结合当地试验林进行持续的观测,探究杉木家系间的遗传变异规律,分析不同性状苗期与成熟林的早晚相关性和稳定性,以保证苗期选择的可靠性,为赣南地区杉木遗传改良和林分营建提供了较好的种质材料。
致谢:江西省研究生创新专项资金项目(YC2022-s424)同时对本研究给予了资助,谨致谢意!
