基于线粒体Cyt b基因的洪泽湖翘嘴鲌群体遗传多样性分析
2023-11-21李大命杨子萍刘燕山谷先坤唐晟凯杨家新
李大命 杨子萍, 刘燕山 谷先坤 唐晟凯 杨家新
(1 江苏省淡水水产研究所,江苏省内陆水域渔业资源重点实验室,江苏南京 210017;2 南京师范大学海洋科学与工程学院,江苏南京 210023)
洪泽湖(118°10′~118°52′E,33°06′~33°40′N)位于江苏省西部淮河下游,水域面积约1 597 km2,是我国五大淡水湖之一。洪泽湖具有相对稳定的水位、宽广的水域和丰富的水生生物资源,为渔业发展提供了良好条件,是我国主要淡水渔业基地之一[1]。近几十年来,由于过度捕捞、水体环境质量下降、水利工程建设等多重因素影响,洪泽湖鱼类物种数量减少,群落结构趋于单一化,鱼类小型化趋势严重,大型肉食性鱼类如翘嘴鲌等资源衰退严重[1-2]。
翘嘴鲌(Culteralburnus)隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲌亚科(Culterinae)、鲌属(Culter),俗称白鱼、翘嘴白丝、大白鱼等,广泛分布于我国各个水系[3]。翘嘴鲌为凶猛肉食性鱼类,是水域生态系统中的顶级消费者,能有效控制小型鱼类种群数量,减轻小型鱼类对浮游生物的压力,在稳定生态系统结构和功能中发挥重要作用[4-5]。近年来,渔业主管部门在洪泽湖开展了翘嘴鲌人工放流,对于恢复翘嘴鲌资源发挥了重要作用,但盲目的增殖放流可能会对鱼类资源遗传结构产生不利影响[6]。因此,迫切需要了解洪泽湖的翘嘴鲌种质资源状况和遗传多样性水平,用于指导洪泽湖翘嘴鲌增殖放流、种质资源保护和管理工作。
目前,分子标记技术被广泛用于研究物种群体的遗传多样性和遗传结构。由于线粒体DNA (mitochondrial DNA, mtDNA)具有分子小、结构简单、进化快、严格的母系遗传等特点,因此成为群体遗传学和分子系统学的重要分子标记[6]。Cytb基因(线粒体细胞色素b基因)是mtDNA的13个蛋白编码基因之一,其结构和功能较为清晰,进化速率适中,包含着从种内到种间的较强进化信号,故被广泛用于物种多样性和系统进化等方面的研究[7-9]。王利华等[10]应用COⅠ和Cytb基因分析了6种鲌属鱼类(达氏鲌、海南鲌、尖头鲌、蒙古鲌、拟尖头鲌和翘嘴鲌)的遗传关系,认为线粒体COⅠ和Cytb基因可以作为DNA条形码鉴定鲌属鱼类。李大命等[11]采用Cytb基因序列研究了滆湖3种鲌属鱼类(翘嘴鲌、蒙古鲌、达氏鲌)的遗传多样性,发现3种鲌鱼的遗传结构趋于单一化。张桂宁等[12]通过Cytb基因序列分析发现,长江下游4个翘嘴鲌群体的遗传多样性较高。2009年,伊西庆[13]采用线粒体ND2基因序列对采集的16尾洪泽湖翘嘴鲌进行了遗传分析,检测到较高的单倍型多样性和较低的核苷酸多样性,但该研究所分析的样本量较少,其结果无法准确反映洪泽湖翘嘴鲌的遗传状况。近十多年,尚未有关于洪泽湖翘嘴鲌遗传多样性的研究报道。本研究拟采用线粒体Cytb基因序列,对6个洪泽湖翘嘴鲌群体的遗传多样性和遗传结构进行分析,以期了解当前洪泽湖翘嘴鲌群体的遗传多样性状况,并为翘嘴鲌增殖放流、种质资源保护及利用提供科学的理论依据。
1 材料和方法
1.1 样品采集和处理
2021年在洪泽湖开展渔业资源监测,采用三重刺网在6个采样点采集翘嘴鲌210尾,其中洪泽群体(HZ)35尾,高渡群体(GD)34尾,顾勒河群体(GLH)34尾,新开河群体(XKH)33尾、临淮群体(LH)36尾,马浪岗群体(MLG)38尾。各群体样品采样点位置见表1。测量每尾鱼的体长,称取体质量,并剪取尾鳍组织放入无水乙醇中保存。

表1 洪泽湖翘嘴鲌各群体采样点位置
1.2 DNA提取、PCR扩增和测序
采用TaKaRa公司试剂盒提取翘嘴鲌的基因组DNA,将DNA溶于TE溶液,于-20 ℃保存备用。用琼脂糖凝胶电泳和紫外分光光度计检测DNA质量和浓度。
线粒体Cytb基因扩增引物为L14724(序列为5’-CGTCAGTCCTTTACTTCGCA-3’)和H15915(序列为5’-AGGGCATACTCACGGGGTTG-3’)[14],引物由生工生物工程(上海)股份有限公司合成。PCR反应体系共50 μL,其中包括:2×Premix TaqTM25 μL(TaKaRa Taq 1.25 U/25 μL、0.2 mM dNTP、1.5 mM Mg2+、色素Marker、比重增加物和稳定剂)、上、下游引物各2 μL (10 μmol/L)、DNA模板2 μL (40 ng/μL )和ddH2O 19 μL。PCR扩增程序为:94 ℃预变性4 min;94 ℃变性40 s,55 ℃复性30 s,72 ℃延伸1 min,35个循环;72 ℃延伸5 min。PCR反应完成后,取4 μL产物经1%琼脂糖凝胶电泳检测,剩余的PCR产物送生工生物工程(上海)股份有限公司进行纯化和双向测序,测序引物与扩增引物一致。
1.3 数据处理和分析
使用BioEdit 7.0.9.0软件[15]读取序列,并参照测序图谱人工校对序列中可能的错误。使用CLUSTALX 2.0软件[16]对序列进行多重比对和排序。使用MEGA 7.0.26软件[17]统计序列的碱基组成、突变位点数和突变类型,计算群体间的Kimura双参数遗传距离,利用邻接法(neighbor-joining, NJ)构建单倍型系统发育树,系统树中节点的自举置信水平应用自引导估计,循环次数为1 000次。采用Network软件构建单倍型网络图,用以展示单倍型之间的进化关系。使用DanSP 5软件[18]计算群体的单倍型数量、单倍型多样性(haplotye diversity,Hd)、核苷酸多样性(nucleotide diversity,Pi)、平均核苷酸差异数(K)等遗传多样性参数。采用Arlequin 3.5软件[19]进行分子方差分析(AMOVA),计算群体间的遗传分化指数(Fst),开展Tajima’sD及Fu’sFs中性检验和构建核酸错配分布曲线,评估翘嘴鲌群体的历史动态。
2 结果
2.1 Cyt b基因序列变异及遗传多样性
本研究获得翘嘴鲌的Cytb基因全序列,其长度为1 141 bp。4种碱基的平均含量分别为A(29.4%)、T(26.6%)、C(29.4%)和G(14.6%),碱基A+T含量(56.0%)大于G+C含量(44.0%),具有明显的碱基组成偏倚性。210条Cytb序列中共发现31个变异位点,其中有11个单突变位点,20个简约信息位点。
6个群体210尾翘嘴鲌共定义了26个单倍型,整个群体的单倍型多样性和核苷酸多样性分别为0.735±0.032和0.001 9±0.000 1,平均核苷酸差异数为2.118(见表2)。6个群体中,马浪岗群体(MLG)的遗传多样性最高(Hd为0.788±0.064,Pi为0.0021±0.0002),高渡群体(GD)的遗传多样最低(Hd为0.618±0.096,Pi为0.001 5±0.000 3)。洪泽湖翘嘴鲌的遗传多样性具有典型的高单倍型多样性和低核苷酸多样性模式。

表2 洪泽湖翘嘴鲌群体遗传多样性参数
2.2 群体单倍型组成及系统发育
6个群体定义了26个单倍型,其中洪泽群体拥有14个单倍型,高渡群体拥有11个单倍型,顾勒河和马浪岗群体各拥有12个单倍型,新开河和临淮群体各有10个单倍型。26个单倍型中,共享单倍型有18个,独享单倍型有8个。其中,共享单倍型Hap3和Hap6为6个群体共有,个体数量分别有17个和105个,占整个群体的比例合计为58.1%;共享单倍型Hap1、Hap2、Hap4、Hap8、Hap11~12、Hap14~17和Hap19~24为部分群体共有,个体数量共有80个,占整个群体的比例合计为38.1%;独享单倍型Hap5、Hap7、Hap9、Hap10、Hap13、Hap18、Hap25和Hap26为各自群体独有,个体数量均只有1个,占整个群体的比例合计为3.8%(见表3)。整体来看,共享单倍型的个体数量多,占群体的比例高,暗示6个群体间存在广泛的基因交流。

表3 洪泽湖6个翘嘴鲌群体单倍型组成
利用红鳍原鲌(Cultrichthyserythropterus)和蒙古鲌(Cultermongolicus)作为外类群,基于邻接法构建单倍型系统发育树(见图1)。结果显示,所有单倍型聚为一个进化分支,分支的置信限水平为100%。单倍型在6个群体中呈混杂分布模式,没有形成与群体地理位置相对应的谱系关系。单倍型网络结构图也得出类似结果(见图2),说明6个群体间没有明显的遗传分化。

图1 洪泽湖翘嘴鲌群体单倍型NJ系统树

图2 洪泽湖翘嘴鲌群体单倍型网络结构图
2.3 群体间遗传结构
6个群体间的遗传距离为0.001 6~0.002 0,遗传距离均值为0.001 9,表明群体间遗传差异较小,亲缘关系较近(见表4)。分子方差分析结果显示,群体内个体间的分子变异占比为100.33%,群体间遗传变异占比为-0.33%,遗传分化指数Fst值为-0.003 3(P=0.624 6)(见表5),表明整个翘嘴鲌群体没有出现遗传分化。两两群体间的Fst值为-0.017 7~0.018 2,且统计检验结果均不显著(见表4),显示群体间没有形成显著的遗传分化。

表4 洪泽湖翘嘴鲌6个群体间的遗传距离(对角线下)和遗传分化指数(对角线上)
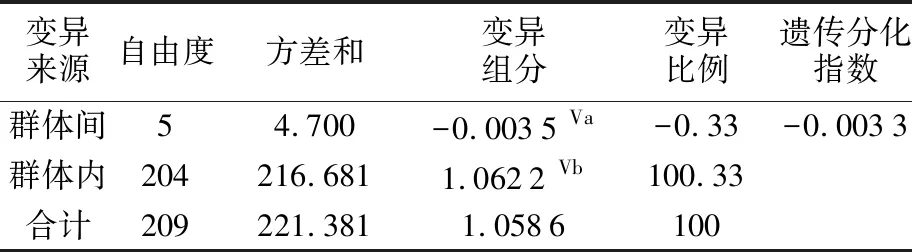
表5 洪泽湖翘嘴鲌群体分子方差分析
2.4 群体历史动态
采用Tajima’sD、Fu’Fs中性检验和核苷酸错配分布曲线分析翘嘴鲌群体的历史动态。结果显示,6个群体的Tajima’sD检验值统计均无显著性差异(P>0.05),除临淮群体(LH)外,Fu’Fs检验值统计均有显著性差异(P<0.05)(见表6),且6个群体的核苷酸错配分布曲线呈现出单峰模式(见图3),表明6个群体显著偏离中性进化。从整体来看,其Tajima’sD和Fu’Fs的检测值分别为-1.683 2和-13.532 9,统计分析具有显著性差异(P<0.05),且核苷酸分布曲线也具有典型的单峰模式(见图3),表明翘嘴鲌在进化过程中发生了群体扩张事件。
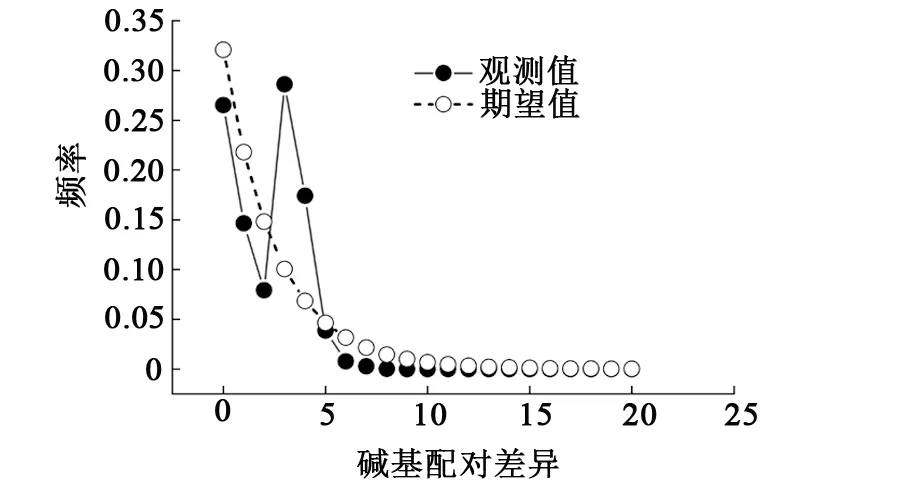
图3 洪泽湖翘嘴鲌群体核苷酸错配分布曲线

表6 洪泽湖翘嘴鲌群体的中性检验
3 讨论
3.1 翘嘴鲌的遗传多样性
遗传多样性是生物多样性的基础,也是评价生物种质资源状况及制定种质资源保护措施的重要依据。单倍型多样性(Hd)和核苷酸多样性(Pi)是衡量群体遗传多样性的重要指标,其数值越大,表明群体遗传变异越丰富,遗传多样性也越高[20-21]。相比较而言,核苷酸多样性考虑各种单倍型在群体中所占的比例,能更为精确地反映群体多态性程度[22]。本研究结果显示,洪泽湖翘嘴鲌的平均单倍型多样性和核苷酸多样性分别为0.735±0.032和0.001 9±0.000 1,6个群体的单倍型多样性在(0.618±0.096)~(0.788±0.064),核苷酸多样性为(0.001 5±0.000 3)~(0.002 1±0.000 2)。Grant等[23]根据单倍型多样性和核苷酸多样性大小,将单倍型多样性和核苷酸多样性之间的关系分为4种类型:高Hd高Pi、高Hd低Pi、低Hd高Pi和低Hd低Pi。据此判定,洪泽湖翘嘴鲌的遗传多样性属于高Hd(Hd≥0.5)低Pi(Pi<0.005)类型,这种遗传多样性模式暗示翘嘴鲌的遗传多样性较低[23]。多年来,受过度捕捞、环境污染、非法采砂等不利因素的影响,洪泽湖环境生态质量下降,翘嘴鲌渔业资源量明显减少,其适宜的栖息环境大幅缩减,个体小型化和种群低龄化趋势加剧,遗传多样性丧失。已有研究结果显示,我国许多水体的野生翘嘴鲌资源遗传多样性具有高Hd低Pi共存现象[11,13,24-26],表明翘嘴鲌遗传多样性较低是一个普遍现象,需要采取措施恢复翘嘴鲌资源量及提高其遗传多样性。
3.2 翘嘴鲌的历史动态
鱼类群体的遗传多样性模式与群体的历史动态有密切的联系。Grant等[23]提出了一个较简单的模式,即利用Hd、Pi来推测群体的历史动态:当Hd≥0.5、Pi≥0.005时,表示群体稳定,具有较悠久的进化历史;当Hd≥0.5、Pi<0.005时,表示群体是经历瓶颈效应后迅速扩张而来的;当Hd<0.5、Pi≥0.005时,表明群体经历了轻微的瓶颈效应,几乎没有影响到核苷酸变异;当Hd<0.5、Pi<0.005时,表明群体近期经历了瓶颈效应。本研究结果显示,翘嘴鲌群体的单倍型多样性较高(Hd≥0.5),而核苷酸多样性较低(Pi<0.005),由于积累核苷酸多样性所需的时间比积累单倍型所需的时间漫长,表明洪泽湖翘嘴鲌群体是从一个有效种群数量较小的种群经快速扩张而来的,但是仍然没有达到积累核苷酸变异所需要的时间。通常情况下,有2种方法可用于推测群体的历史动态:(1)核苷酸错配分布曲线。若分布曲线呈单峰分布,表明群体经历了种群扩张;若分布曲线呈双峰分布,表明群体没有经历过种群扩张,较为稳定[27-28]。(2)Tajima’sD和Fu’Fs中性检验。若中性检验值为负值且统计分析结果为显著,表明群体可能经历了种群扩张[29-30]。本研究结果显示,洪泽湖翘嘴鲌群体的Tajima’sD和Fu’Fs中性检验值均为负值,统计检验结果具有显著性差异(P<0.05);核苷酸错配分布曲线呈典型的单峰型,均说明翘嘴鲌进化过程中偏离了中性选择,经历了显著的种群扩张,形成了高Hd和低Pi的遗传多样性模式,这与已有的研究结果[11,13]一致。
3.3 翘嘴鲌的遗传结构
通常采用遗传分化指数(Fst)来评估群体间的遗传分化程度。Wright[31]指出,若Fst值为0~0.05,表明群体间不存在分化;若Fst值为0.05~0.15,表明群体间存在中度分化;若Fst值为0.15~0.25,则为高度分化。分子方差分析结果显示,遗传变异主要发生在群体内个体间,整个群体的Fst值为-0.003 3(P=0.624 6),两两群体间的Fst值为-0.017 7~0.018 2,P>0.05,均表明翘嘴鲌群体间没有出现遗传分化,群体的分子系统发育树和单倍型网络结构图也支持上述观点。从群体的单倍型组成来看,群体间有多个共享单倍型,且共享单倍型的个体数量占比较高,说明群体间存在广泛的基因交流。对于鱼类而言,地理阻隔或同一水域中存在不同的栖息环境可能会限制基因交流,从而使群体间产生遗传分化现象[32]。本研究中的6个群体虽处于洪泽湖的不同水域,水域生态环境也存在一定的差异[33-34],但群体所在的地理位置较近,没有形成有效的地理隔离,而翘嘴鲌游泳能力强,活动范围广,也助推了群体间的基因交流。此外,近年来渔业主管部门在洪泽湖开展了翘嘴鲌增殖放流活动,大量人工养殖群体的加入容易导致群体遗传结构的单一和均质化,这一现象在其他水域的增殖放流种类中均有发现[35-37]。根据翘嘴鲌的遗传结构特点,应将6个群体作为一个进化单位进行管理和保护。
3.4 翘嘴鲌资源保护对策
对鱼类群体遗传多样性和遗传结构的研究可以为渔业资源管理措施及物种保护策略的制定提供重要的数据支撑。研究结果显示,洪泽湖翘嘴鲌的遗传多样性具有高单倍型多样性和低核苷酸多样性共存现象,表明翘嘴鲌遗传资源较为贫乏。本研究洪泽湖翘嘴鲌6个群体间没有显著的遗传分化,因此可将洪泽湖不同水域的翘嘴鲌群体作为一个整体进行管理和保护。针对洪泽湖翘嘴鲌的遗传多样性现状,应当采取措施,加大对翘嘴鲌的保护力度:(1)强化渔业资源管理,加大执法力度,严厉打击非法捕捞,禁止电、毒、炸等作业方式;(2)控制环境污染,开展生态环境治理,恢复水生植被,构建人工鱼巢,为翘嘴鲌生存和繁殖提供良好的生态环境;(3)科学规范地进行翘嘴鲌增殖放流,提高翘嘴鲌的成活率,开展放流群体和野生群体的遗传评估,跟踪监测翘嘴鲌资源变化,逐步提高翘嘴鲌的遗传多样性。
