中全新世海南潭门滨珊瑚的生长率特征及其气候意义
2023-11-13李悦儿余克服颜廷礼江蕾蕾
李悦儿,余克服,2,颜廷礼,江蕾蕾
[1.a.广西大学 海洋学院;b.广西南海珊瑚礁研究重点实验室;c.珊瑚礁研究中心,南宁 530004;2.南方海洋科学与工程广东省实验室(珠海),广东 珠海 519080]
造礁珊瑚作为理想的重建高分辨率气候记录的海洋资料,近数十年来珊瑚重建历史时期月-年分辨率气候环境变化的研究取得丰硕成果。其中,对于珊瑚骨骼生长条带的认识和理解是将珊瑚作为高分辨率气候重建的重要基础。中国珊瑚生长率的研究,最早始于民国时期,马廷英(1936;1937)于1936 年4—9 月赴东沙群岛调查研究现代造礁石珊瑚成长率,并指出造礁珊瑚成长率确不失为研究古气候的一个可靠工具;之后由聂宝符于1984年开始使用“珊瑚生长率”这一术语(聂宝符,1984),并系统介绍了珊瑚生长率的研究地点、珊瑚种类的选择、采样要求及生长率获取、测量、拼接方法等(聂宝符 等,1997)。珊瑚生长率是造礁珊瑚最基本的生长参数,表示珊瑚骨骼生长速率,即珊瑚骨骼沿最大生长轴每年所增长的量,以cm/a或mm/a为单位,在X光片上珊瑚生长率表现为一组黑白条带之间的间隔距离(Knutson et al., 1972)。此外,珊瑚生长率不受后期成岩作用改变(Reuter et al.,2005),能提供准确的年分辨率的气候环境变化信息,并且珊瑚骨骼的地球化学指标分析也需在其构建的年代框架下进行(Tudhope, 1994),因此珊瑚生长率的研究发展至今仍具有不可或缺的意义。
珊瑚生长率是诸多因素的综合反映,表层海水温度(Sea Surface Temperature, SST)、表层海水盐度(Sea Surface Salinity, SSS)、光照、水动力条件等都会对珊瑚的生长造成影响(Lough et al., 1997,2000, 2011, 2014;施祺 等,2002;苏瑞侠 等,2003;Wang et al., 2010)。研究表明,SST 是珊瑚骨骼生长率的关键环境控制因素(Lough et al.,1997, 2000, 2011, 2014;施祺 等,2002;苏瑞侠等,2003;Cooper et al., 2012;陈雪霏 等,2023),并且珊瑚生长率和SST存在显著的线性关系,基于这种关系可以重建有器测资料之前的历史时期SST变化(Nie et al., 1997, 1999; Dang et al., 2020)。南海珊瑚礁星罗棋布、种类众多、发育历史久,自20世纪90年代以来,南海珊瑚生长率及其与海表温度变化关系的研究迅速发展。如Nie等(1997;1999)最早利用滨珊瑚生长率重建西沙海域过去220 a 以来的SST 变化;黄博津(2013)和张会领(2014;2017)等也根据西沙海域的滨珊瑚生长率,重建了罗马暖期和中晚全新世特征时段的SST变化,其结果较好地响应了同期全球的气候变化;陶士臣等(2021)重建过去500 a(1520—2007 AD)西沙海域连续的SST序列显示,西沙海温存在1520—1825 AD的相对低温期和1826—2007 AD的快速升温期2个主要时段,其对赤道西太平洋暖池环境变化响应敏感。此外,部分学者通过珊瑚生长率的年际和年代际变化重建了南海北部厄尔尼诺-南方涛动(El Niño-Southern Oscillation, ENSO)和太平洋多年代际涛动(Pacific Decadal Oscillation, PDO)活动的历史(林丽芳 等,2018;Dang et al., 2020)。
综上,珊瑚生长率是重建区域SST变化的重要代用指标,同时其年际周期还能揭示历史时期ENSO 等变化。然而,受珊瑚自身生长时段和样品采集等因素的限制,目前基于珊瑚生长率的气候记录主要集中在工业革命以来的时段(Yu, 2012; Tierney et al., 2015),极少有年代更早的珊瑚生长率研究。而中全新世时期作为全球变暖背景下气候环境状况的地质历史相似型,重建中全新世时期的气候变化,对揭示气候变化机制和提高未来气候预测的准确性具有重要意义。中全新世(8200—4200 a BP)是全新世以来最为温暖湿润的时期,又称为全新世大暖期,在其最盛期时(8000—6000 a BP)全球许多地方都比现在温暖(施雅风,1992;de Menocal et al., 2000)。目前,基于代用资料记录和古气候模拟重建对于中全新世起止时间、总体气候特征以及大暖期的定义已有丰富的研究成果。来自全球不同地区、不同类型的地质记录,总体都揭示了中全新世温暖湿润、期间存在气候突变事件等气候特征(Bond et al., 1997; Chen et al., 2016; Ramos-Román et al., 2016; van Beynen et al., 2017; Badino et al., 2018; Novenko et al., 2019)。而冰心、孢粉、石笋、沉积物等地质记录由于空间分布不均、时间分辨率低及连续性差等特点,对中全新世期间影响气候的关键大气环流系统和ENSO等海气相互作用的研究较为有限(姜大膀 等,2022)。鉴于热带海洋在气候系统中的独特地位,中全新世气候变化的研究还需要更多来自热带海区的高分辨率气候记录(汪品先 等,1999)。
因此,本文以海南潭门珊瑚岸礁的滨珊瑚为材料,利用X射线照相技术与图像处理方法对珊瑚生长率进行测量与分析,获得中全新世(6143—4356 a BP)海南潭门滨珊瑚的生长率特征,并基于该区域现代(2005—2021 AD)滨珊瑚生长率与海表温度的关系,重建中全新世期间共406 a 的年均SST序列,旨在为全面理解中全新世的热带气候特征提供新的信息。
1 材料与方法
1.1 研究区域及采样点
本文所使用的澄黄滨珊瑚(Porites lutea)岩心样品采自南海北部海南岛东部琼海市潭门镇(19°15´N、110°38´E)珊瑚岸礁(图1)。该区属热带季风气候,分明显的旱雨两季,年均温24.1°C,最高气温出现在7月,年均值为28.3°C,最低气温出现在1 月,年均值为17.8°C。多年平均降水量为1 653.4 mm,全年降水量主要集中在每年夏季季风盛行的雨季(5—10 月),占全年降雨量的76%~81%。潭门海域的年平均表层水温约25.6°C,表层盐度约32.6‰,海水透明度良好,这些条件非常适合珊瑚群落的生长发育(Yan et al., 2019)。
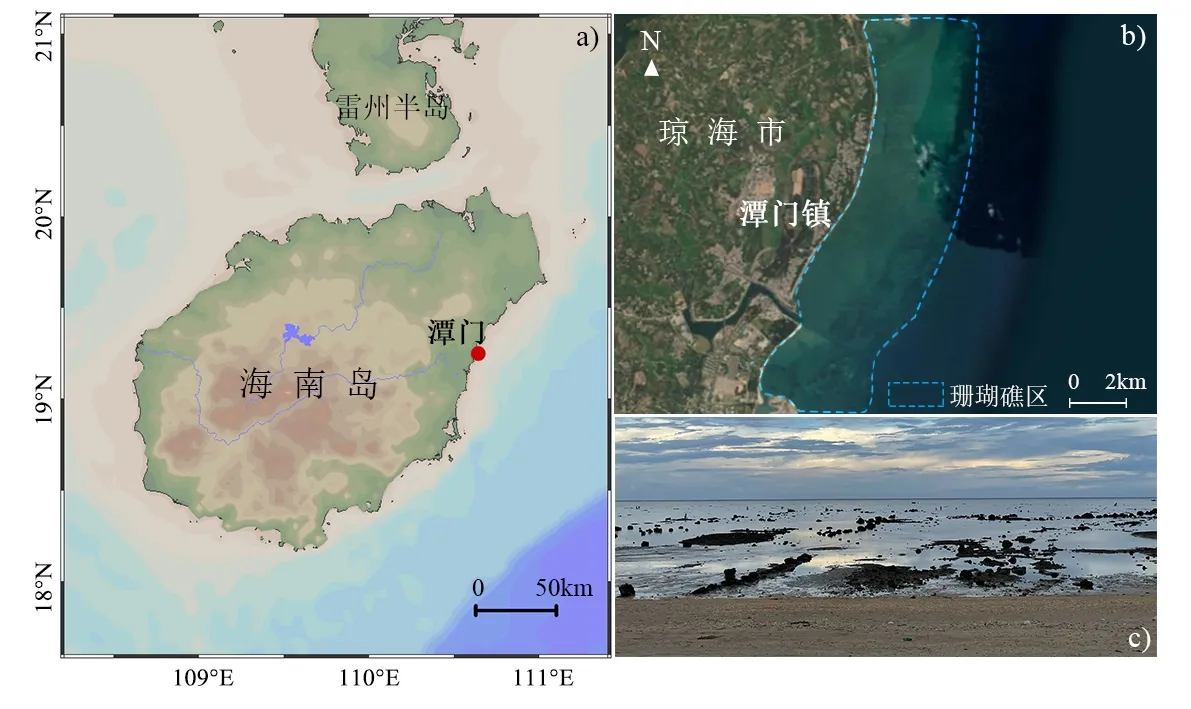
图1 研究区域(a)和采样位置(b.潭门珊瑚岸礁的分布;c.潭门礁坪实景)Fig.1 Study area(a) and locational distribution of coral core samples ( b.the distribution of Tanmen coral reefs; c.the photo of Tanmen sea area)
2021年7和9月于潭门珊瑚岸礁礁坪处水深5 m左右的位置使用手持式液压钻机从顶部钻取珊瑚岩心,岩心整体呈圆柱状,长度约为20~30 cm,直径为6 cm。在实验室内用切片机将上述柱状珊瑚岩心切成厚度约为7~8 mm 的薄板。将切割完成的珊瑚薄板放入质量分数为5%的H2O2溶液中浸泡48 h,再用超声波清洗4次,每次20 min以去除残留物质,最后将珊瑚薄板放入40°C的恒温烘箱中烘干48 h以供后续使用。
1.2 定年及生长率测量
珊瑚样品的高精度铀系年代由澳大利亚昆士兰大学放射性同位素实验室测定,年代介于6143—4356 a BP(颜廷礼,2022)。对珊瑚生长率序列分析需以珊瑚薄板上的定年点为基准,再计数珊瑚骨骼年层建立起生长纹层的年代框架。H1 心为2021年7月采集的现代活珊瑚样品,其年代范围以顶面年龄(2021 AD)为基准,再根据生长纹层计年获得。
使用X光机对处理后的珊瑚薄板拍摄等比例的X光照片(图2)。通过X光照片可以清晰看出珊瑚骨骼由明暗相间的条纹组成,这些条纹即珊瑚在不同季节形成的高/低密度生长条带,相邻的一条亮条带的宽度加上一条暗条带的宽度就是珊瑚一年的生长量。采用图片处理软件作为珊瑚生长率测量的主要工具,使用医学软件RadiAnt DICOM Viewer将X光负片灰度图像转化为X光正片,然后使用该软件中的测距工具测量珊瑚年生长率(cm/a)。在X 光正片上,以白色条带中间为起止点,一对黑-白色条带的宽度作为珊瑚骨骼一年的生长量进行测量。

图2 海南潭门珊瑚样品X光片及定年点位置Fig.2 Positive X-ray images of the coral skeletons from Tanmen,and the positions of subsamples used for U-Th dating
需要指出的是,在采样和后期切片过程中,如果珊瑚岩心样品不是垂直于珊瑚年生长层钻取,切片方向不垂直于珊瑚年生长层等,均可能引起生长率测量的人为偏差(Saenger et al., 2009)。笔者已经充分认识到取岩心和切片对生长率的可能影响,在取样过程中已尽量降低人为因素对生长率的影响。与此同时,为了尽可能减小误差,对23段化石珊瑚的生长率进行10次测量,去除一个最大值和一个最小值后,求算数平均作为珊瑚骨骼生长率结果。对于取到的唯一的现代珊瑚H1,X光片中显示其中分布有若干孔洞,表明现代珊瑚受生物侵蚀,相对于其他样品X 光片的生长条带而言不那么清晰,但仍能识别出明暗相间的条带。为了保证测量结果的可靠性,首先对现代珊瑚样品薄板上明显的生长轴进行直接测量,并重复10次,再对X光片使用DICOM explorer 软件进行10 次测量,同时结合Acycle软件对图像灰度变化进行分析。
1.3 数据分析方法
通过上述测量与计算获得1段现代珊瑚和23段中全新世化石珊瑚的生长率数据,并且利用器测资料获取现代潭门海域的气候环境参数,其中月均及年均海温数据来自英国气象局哈德利中心提供的1°×1°网格全球海温数据集HadISST①https://www.metoffice.gov.uk/hadobs/hadisst/(Rayner et al., 2003),年均海表盐度数据来自中国科学院海洋科学数据中心提供的1°×1°网格全球海洋温盐现场观测数据集②http://msdc.qdio.ac.cn,年均太阳辐射数据来自美国国家海洋和大气管理局国家气候数据中心提供的NCEPNCAR再分析资料的1°×1°网格净短波辐射通量③https://psl.noaa.gov/data/gridded/index.html。
为探究潭门珊瑚生长率与气候环境参数的关联,对现代珊瑚生长率与气候环境参数进行Pearson偏相关性分析,在控制其他变量影响的条件下分析变量间的线性相关性;再通过建立一元线性回归模型进一步确定变量之间的因果关系,考虑到样本数量不够大,且数据中存在若干明显异常值,因此,在对比各回归方法并对拟合结果比较后,选择使用稳健的鲁棒回归(Robust Regression)建立模型。Pearson偏相关性分析和鲁棒回归的一元线性回归分析均使用SPSS软件完成。
为探究潭门珊瑚生长率序列与气候环境参数的周期成分及变化规律,对现代珊瑚生长率序列和器测海温进行傅里叶频谱分析,以分离其显著的年际周期。中全新世化石珊瑚生长率序列中有16 a的重叠部分,重叠年代为4381—4385 a BP(共计5 a)、4475—4477 a BP(3 a)、5235—5237 a BP(3 a)和5410—5414 a BP(5 a),重叠部分的珊瑚生长率结果不完全一致,相差范围为0~0.2 cm/a,但生长率变化趋势相对一致,对重叠部分的生长率结果取算数平均值,拼接形成4段连续时间较长的生长率序列:5427—5394 a BP (34 a)、5243—5209 a BP(35 a)、4515—4456 a BP(60 a)和4404—4356 a BP(49 a);再对以上序列进行连续小波变换,分析这些时段的时频变化特征,以进一步探讨珊瑚生长率在各周期波动的趋势变化。傅里叶频谱分析和连续小波分析均使用Past软件完成。
2 结果分析
各段样品的编号及生长率测量结果见表1所示。现代珊瑚平均生长率为1.260 cm/a,最大生长率为1.400 cm/a,最小生长率为1.150 cm/a;中全新世化石珊瑚的平均生长率为1.079 cm/a,最大生长率为1.670 cm/a,最小生长率为0.607 cm/a;中全新世时期珊瑚生长率总体略低于现代珊瑚0.181 cm/a,而生长率的变化幅度大于现代。中全新世珊瑚生长率序列显示其具有明显的波动变化:在6143—5855 a BP 珊瑚生长率波动下降,而后在5774—5691 a BP迅速上升;在5672—5647 a BP 珊瑚生长率保持较低水平,随后在5576—5410 a BP 逐渐上升并在5414—5235 a BP 保持较高水平;在5237—5209 a BP 生长率又下降,直到5179—5149 a BP 期间降至最低,接着在5116—4356 a BP 生长率又大幅度波动上升。同时,在序列中可见3段连续时间较长的低生长率特征时期,还明显可见4515 a BP 之后珊瑚生长率变率显著增大,呈现更为复杂的波动变化。此外,频谱分析显示现代和中全新世珊瑚生长率均存在显著的3~7 a周期信号,但在5427—5394、5243—5209、4515—4456、4404—4356 a BP 和 现代,期间显著的主周期及频率不断转变。

表1 海南潭门珊瑚生长率测量结果Table 1 Results of growth rates of corals from Tanmen,Hainan Island
3 讨论
3.1 海南潭门现代珊瑚生长率与SST的关系
大量研究通过统计学方法以及模型分析法业已表明,南海北部珊瑚生长率主要受温度控制,并据此建立珊瑚生长率与SST 的关系(Nie et al., 1997,1999;施棋 等,2002;苏瑞侠 等,2003;黄博津等,2013;陈天然 等,2013;张会领 等,2014)。目前采用一元线性回归已建立的南海北部珊瑚生长率与SST 线性关系有:Nie 等(1997;1999)建立海南岛南部滨珊瑚生长率与SST的线性回归方程为SST=2.52×L+24.22(r=0.85),西沙群岛滨珊瑚生长率与SST 的关系为SST=2.25×L+24.772(r=0.876);张会领等(2017)建立西沙永兴岛滨珊瑚生长率与SST 的 线 性 回 归 方 程 为SST=2.0×L+24.91 (R2=0.61);林丽芳等(2018)建立黄岩岛澄黄滨珊瑚生长 率 与SST 的 关 系 为SST=3.49×L+23.058 (r=0.697);陶士臣等(2021)建立西沙永兴岛滨珊瑚生长率与SST 的线性回归方程为SST=2.25×L+24.767(R2=0.61)。但因为珊瑚对环境变化的敏感程度在不同种类、不同生长地点存在显著差异,所以珊瑚生长率与温度的回归方程并不完全一致(何学贤 等,1999)。
海南潭门海域尚缺乏珊瑚生长率与SST关系的构建研究,因此,首先需探究现代海南潭门珊瑚岸礁区珊瑚生长率是受单因子SST 影响还是受SSS、光照的联合控制。对现代珊瑚生长率数据与区域气候环境参数采用偏相关分析(表2),一阶偏相关中,在控制SSS不变的条件下,SST和光照均与珊瑚生长率有显著的正相关关系,但二阶偏相关中,在控制了SSS 和SST 后,光照与生长率不显著相关。在所有不同控制变量情况下,SST与珊瑚骨骼生长率始终保持显著正相关关系,因此SST是海南潭门海域滨珊瑚骨骼生长参数变化的主要控制因素。图3 中a、b、c 分别为2005—2021 年潭门海域珊瑚生长率与SSS、SST 和光照的变化曲线,可知珊瑚生长率变化与SST的变化高度相关且通过显著性检验。此外,虽然光照和盐度与珊瑚生长率的相关性不显著,但在变化曲线上存在一定程度的谐调性,所以光照和盐度是海南潭门滨珊瑚骨骼生长的次要影响因素。

表2 海南潭门珊瑚生长率与环境参数的偏相关分析结果Table 2 Partial correlation analysis results of coral growth rate and environmental parameters in Tanmen
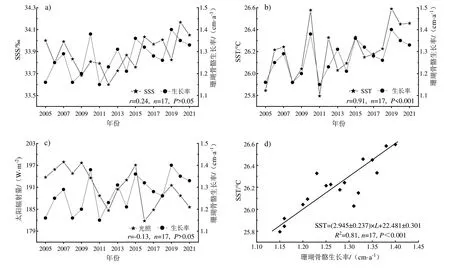
图3 海南潭门现代珊瑚生长率与环境参数(a.SSS;b.SST;c.光照)的变化关系及与SST的回归模型(d)Fig.3 Relationship between modern coral growth rate and environmental parameters (a.SSS; b.SST; c.Solar radiation) in Tanmen, and L-SST linear regression model (d)
由于其他环境因素与珊瑚生长率变化不显著相关,在建立海南潭门现代珊瑚生长率与SST的关系时予以忽略。使用鲁棒回归建立的海南潭门海域LSST一元线性回归模型如图3-d所示:
式中:SST为潭门海域年均海表温度(℃);L为珊瑚年生长速率(cm/a);s.e.为标准误差。根据两者的函数关系,也即珊瑚生长率每增加1 cm,代表海温上升约2.7~3.2℃。江巧文等(2016)已证实琼东海域澄黄滨珊瑚骨骼生长率与SST表现显著正相关关系,这与本文结论一致。因此,海南潭门现代滨珊瑚的生长率能较为准确地反映该区域SST变化。
3.2 海南潭门中全新世SST 总体呈波动变化特征,并存在3个显著的SST偏低期
考虑到水深对珊瑚生长率也有所影响,南海中全新世的海平面比现代高,在7.0—5.5 ka BP 时期高出现代海平面2~3 m(时小军 等,2007)。颜廷礼(2022)根据现代珊瑚和中全新世化石珊瑚的铀系年代,并结合珊瑚生长与海平面的关系,重建了潭门海域珊瑚岸礁记录的中全新世海平面波动历史;大型块状滨珊瑚的生长上限充其量只能达到大潮低潮面,因此礁坪面成古高海面的极好标志。在采样过程中采集的样品均为原位、未受外力搬运的珊瑚,现代珊瑚与化石珊瑚均采自于礁坪处;再根据海平面重建结果,中全新世化石珊瑚生长位于平均大潮低潮面和平均小潮低潮面之间,古今珊瑚生长时期的水深基本相似。因此,中全新世与现代的珊瑚生长率具有可比性。
将化石珊瑚的生长率数据带入以上建立的潭门珊瑚岸礁区海表温度与珊瑚生长率的转换方程中,获得海南潭门中全新世(6143—4356 a BP)期间406 a的年均SST,图4为定量重建的海南潭门中全新世SST变化及化石珊瑚生长率序列。中全新世平均SST 为25.7±0.54℃,与现代(2005—2021 AD)平均SST基本接近。中全新世年均SST变化范围为24.7~26.8℃,在重建时段内存在明显的波动变化,存在3个维持时间较长的低海温特征时期——5860、5660、5160 a BP,珊瑚生长率重建的平均SST 为25.0±0.54℃。

图4 定量重建的海南潭门中全新世SST变化及化石珊瑚生长率序列Fig.4 Coral growth rate-SST in Tanmen during Mid-Holocene and sub-fossil coral growth rate sequence
珊瑚生长率重建的SST 显示,中全新世6—4 ka BP 南海北部SST 与目前全球变暖程度加剧下的现代海温十分接近,这可能表明当时气候温暖程度与现代相当。目前一系列全新世温度集成序列显示,中全新世期间8.0—6.0 ka BP时为暖期鼎盛期,而6.0—4.0 ka BP 总体温度开始呈现下降趋势(Marcott et al., 2013; Osman et al., 2021; Rao et al.,2022)。南海北部6 ka BP之前的珊瑚记录显示中全新世早中期SST 明显高于现代,如Yu 等(2005)利用雷州半岛全新世不同时期发育的珊瑚月分辨率的Sr/Ca和δ18O 记录重建显示,在6.8—5.0 ka BP 期间SST比现代(20世纪90年代,该区最暖的10年)高0.9~0.5°C;Wei等(2007)利用海南岛三亚滨珊瑚的月分辨率Sr/Ca 记录重建中全新世海温记录显示,在6.5—6.1 ka BP 期间年均SST 比现代高0.2~1.5°C。与上述研究相比,本文重建时段的总体SST并没有显著高于现代,显示中全新世6—4 ka BP南海北部SST从鼎盛温暖期开始下降的过程。南海黄岩岛海域珍贝海山底部沉积物中的浮游有孔虫记录也支持该结果,由有孔虫壳体内部Mg/Ca比重建的全新世SST 变化显示,在9.1—6.3 ka BP,SST 自24.5°C迅速升至29.3°C,达到全新世最大值,温暖程度明显高于现代;而在6.3—5.3 ka BP,SST 自27.7°C降至24.8°C(杨俊 等,2020)。
值得注意的是,来自全球的不同代用资料总体都揭示了中全新世8—6 ka BP比现代更加温暖湿润的气候特征,但对于6—4 ka BP区域气候与现代相比如何还存在争议。一些研究认为部分区域在6—4 ka BP期间延续了暖期鼎盛期温暖湿润的气候特征,气候温暖程度仍超过现代,如黎广钊等(1996)根据孢粉分析、有孔虫氧同位素分析、历史记载及现代气温观测资料综合探讨广西沿海全新世以来的气候变化,结果显示中全新世中期(6—4 ka BP)时,气候炎热潮湿,平均气温高于现今3~5℃;Kong等(2014)利用珠江口HKUV16 岩心的长链烯酮不饱和指数重建了香港沿海地区8 ka BP 以来的SST 变化,结果显示7.4—2.6 ka BP SST 保持长期稳定且高于现代约2°C。而也有越来越多的证据表明,全球大部分区域在该时期气候转向寒冷干燥,与现代相比温度可能偏低,如Magny等(2004)综合了南北半球海洋和陆地的植被、海温、极地冰盖、热带冰心等44 个替代指标记录,证明5.6—5.0 ka BP 气候寒冷干燥,且具有全球性;Wanner等(2008)基于全球范围不同种类的气候代用资料,并结合一般环流模型(GCMs)和中等复杂性地球系统模型(EMICs)结果,建立了从全新世中期到前工业时代的气候变化框架,其中确定5.4—4.8 ka BP 为新冰期冰川推进的8个时期之一,当时气候突然转冷导致南北半球山地均有冰川前进出现(Thompson et al., 2006);侯光良(2009)建立的中国东南区200 a分辨率温度变化序列中,中全新世6—4 ka BP低于现代年均温约0.3℃;Abram 等(2009)利用西太平洋暖池边缘印度尼西亚和巴布亚新几内亚的珊瑚Sr/Ca值重建中全新世SST,结果显示5.5—4.3 ka BP 间年均SST 低于现代(1982—1993 AD)约1.2℃;Rodriguez等(2019)利用热带大西洋西部2个年代分别为6427 和5199 a BP 的化石珊瑚,通过骨骼Sr/Ca值和U/Ca值获得年分辨率的Sr-U记录并重建SST序列,结果显示重建的年均SST比现代器测记录低0.5~0.75°C。
高分辨率气候重建序列显示,即使是气候系统稳定性高的中全新世期间仍存在多次冷暖波动。如Yu等(2004)根据雷州半岛角孔珊瑚礁剖面的生长间断和高分辨率的珊瑚骨骼Sr/Ca重建表明,7.0—7.5 ka BP期间南海北部至少发生9次大振幅的冷事件;方修琦等(2011)重建的百年尺度全新世温度集成序列表明,全新世大暖期存在多次百年尺度的寒冷事件。本文基于海南潭门珊瑚生长率重建的中全新世温度也呈波动变化的特征:在6.1—5.7 ka BP,SST 整体偏暖,部分时段温暖程度超过现代,但5860 a BP 前后出现一个明显的低SST 时期;在5.6 ka BP又出现了低SST特征时期,此后数百年温度又小幅度波动回升;在5.2—4.6 ka BP,SST整体偏冷,部分时段SST比现代低1℃;在4.5 ka BP前后SST大幅度波动变化,而4.4 ka BP之后SST持续波动上升甚至超过现代1℃,推测之后发生的4.2 ka BP气候突变事件打断了这一变暖趋势。
在5860、5660 和5160 a BP 显著低海温特征时期,南海北部可能受到规模较大的冷事件影响,在其他代用资料重建的全新世气候变化中也记录了这几次冷事件。如瞿文川等(1997)利用长江下游西太湖沉积岩心的总有机碳含量、δ13C、磁化率、色素含量以及氢指数等记录重建全新世的气候变化,结果显示在7.2—5.7 ka BP 期间,该湖区普遍温暖湿润,而5.7 ka BP以来气候转向寒冷干燥。李秋子(2018)利用东海陆架泥质区D03 钻孔的底栖有孔虫组合重建全新世海洋沉积环境变化,结果显示5.8—5.6 ka BP 为明显的气候变冷时期。牛蕊等(2017)利用贵州草海南屯泥炭的腐殖化度和烧失量序列重建中全新世以来的气候变化显示,在6.0和5.0 ka BP 左右出现较为明显的降温事件。Sadler等(2016)利用热带太平洋西部大堡礁的滨珊瑚Sr/Ca值重建中全新世区域SST,结果显示5.2 ka BP为中全新世的冷事件时期,重建SST比现代(2009—2014 AD)器测平均SST 低1.31~2.76℃。Chang 等(2008)利用中国东海东北部沉积岩心中的放射虫重建全新世以来的SST,结果显示在5.1—4.6 ka BP发生SST 骤降的冷事件。与此同时,对Marcott 等(2013)根据73 个地质记录集成的全球全新世温度变化序列进行再分析,发现在5.8 ka BP增温短暂停滞,在5.5 ka BP 温度明显下降,在5.2 ka BP 温度大幅度降低,这与本文珊瑚记录中的3个低海温特征时期基本对应。
3.3 南海北部中全新世ENSO 活动总体比现代弱,但ENSO变率呈现逐渐增强的趋势
ENSO 是地球气候系统中最强烈的气候振荡,在年际时间尺度上(<10 a)主导着全球大部分地区的气候变化。ENSO 直接影响热带太平洋的SST、SSS、降水和热带气旋活动,还能通过大气遥相关影响南海北部海洋环境变化。不少研究表明,南海北部珊瑚对ENSO活动有良好的记录能力,如邓文峰(2007)利用海南三亚中全新世的珊瑚骨骼Sr/Ca重建了月分辨率SST序列,频谱分析显示SST存在显著的ENSO周期,表明在中全新世的南海气候受ENSO 活 动 影 响;Jiang 等(2021a;2021b;2023)利用西沙群岛过去2000 年的珊瑚骨骼Sr/Ca重建了罗马暖期、黑暗时代冷期以及中世纪气候异常期月分辨率SST 序列和ENSO 变率,结果显示ENSO 呈现波动性的演化过程。海南潭门海域现代器测SST资料与现代珊瑚生长率的频谱分析结果如图5-a 所示,现代观测和珊瑚记录在年际尺度均存在较为显著3~7 a 的ENSO 周期,说明海南潭门海域受到ENSO活动的显著影响,并且该海域的珊瑚生长率能响应ENSO活动。

图5 南海北部中全新世4个时段的ENSO变率变化分析(a.南海北部现代珊瑚与器测资料的频谱分析;b.中全新世4段拼接较长序列的小波变换;c.中全新世4个时段的概率密度统计)Fig.5 Analysis of ENSO variability in four periods during the Mid-Holocene in the northern South China Sea (a.Spectral analysis of modern coral and instrumental data in the northern South China Sea;b.The wavelet transform of 4 long sequences of the Mid-Holocene;c.Probability Density Statistics for 4 Periods of the Mid-Holocene)
中全新世期间4 段相对较长的生长率序列(5427—5394、5243—5209、4515—4456 和4404—4356 a BP)的小波变换结果如图5-b 所示,在4 个时段中均捕捉到ENSO周期,在5427—5394 a BP有一个显著的5.7~8.0 a的ENSO周期,在5243—5209 a BP 仅出现了一个低频的2.0~2.8 a 的ENSO 周期,在4515—4456 a BP有一个显著的2.8~5.5 a的ENSO周期,在4404—4356 a BP出现一个高频的5.7~8.0 a的ENSO周期和一个低频的3.2 a的ENSO周期。这表明南海北部中全新世存在ENSO活动,但与现代相比中全新世珊瑚生长率的ENSO主周期改变且频率显著降低,中全新世南海北部ENSO 信号较现在弱。
来自南海北部的其他珊瑚证据也显示中全新世ENSO活动总体偏弱。Yu等(2010)基于珊瑚骨骼的铀系年代和14C 年代对南海碳库年代与El Niño的关系研究表明,6498—5662 a BP的高碳库年代,指示ENSO 活动明显减弱。Wei 等(2007)利用采自海南岛年代为6500—6100 a BP 的3 个澄黄滨珊瑚,通过月分辨率的骨骼Sr/Ca 值重建中全新世南海SST 变化,发现序列中存在ENSO 活动周期,谐波分析后6324—6299 a BP 和6217—6175 a BP 的夏季SST 序列中分别表现出4.9 和7.8 a 的ENSO 周期,但与现代器测反映的ENSO相比总体偏弱。孙东怀等(2007)利用年代为5400 a BP 的澄黄滨珊瑚探究中全新世亚洲季风气候季节性与年际变率,对δ18O 序列的谱分析显示,冬季温度序列存在3.2 和2.7 a的ENSO周期,但对比现代冬季温度周期频率较低,而夏季降水序列不存在明显的ENSO周期。
以上4 个时段的ENSO 周期转变指示中全新世ENSO 活动不断发生变化,为了进一步量化ENSO变率,根据珊瑚序列较短的特点采用概率密度函数(Probability Density Function, PDF)统计方法,该方法已被证实在量化ENSO变率上具有可靠性,并用于重建西南太平洋中世纪气候异常的部分时期与20 世纪早期相似的弱ENSO 变率(Lawman et al.,2020)。PDF 统计结果显示(图5-c),4515—4456和4404—4356 a BP时段ENSO变率显著增强,中全新世以来(5427—4356 a BP)ENSO 变率呈现逐渐增强的态势,中全新晚期4.4 ka BP 较5.4 ka BP 时期ENSO变率增强40%~60%,表明ENSO系统在中全新世晚期活跃,并可能影响4.2 ka冷事件的发生。
南海北部其他高分辨率的海洋记录也揭示了中全新世晚期ENSO 变率逐渐增强的趋势。Sun 等(2005)利用采自海南岛东部年代为4.4 ka BP 的滨珊瑚骨骼δ18O 重建中全新世的SST和SSS记录,其中冬季SST 显示出显著的6.7 a 周期,这揭示了4.4 ka BP 期间强ENSO 对南海冬季SST 的影响。Zhou等(2022)利用采自西沙群岛的年代为5—3 ka BP的3个鳞砗磲,通过月分辨率的骨骼Sr/Ca值重建中全新世南海北部ENSO变率变化,结果显示全新世中晚期ENSO变率不断增强。
4 结论
本文利用海南潭门珊瑚岸礁采集的1段现代珊瑚岩心和23段中全新世化石珊瑚岩心,通过X射线照相技术与图像处理方法对珊瑚生长率进行测量与分析,获得了中全新世海南潭门滨珊瑚的生长率特征,并基于现代珊瑚生长率影响因素分析建立了海南潭门海域珊瑚生长率与海温的定量转换关系为:
SST =(2.945 ± 0.237)×L+ 22.481 ± 0.301(1 s.e.),在此基础上通过化石珊瑚的年分辨率生长率数据,定量重建了中全新世期间累计406 a 的海温序列,结果显示:
1)中全新世化石珊瑚的平均生长率为1.079 cm/a,变化于0.607~1.670 m/a之间,并且存在明显的波动过程。在序列中可见3段连续时间较长的低生长率特征时期,4515 a BP 之后年际变率显著增大,呈现更为复杂的波动变化。
2)重建中全新世(6143—4356 a BP)的平均SST为25.7±0.54℃,在误差范围内与现代(2005—2021 AD)平均SST 基本接近;中全新世年均SST变化于24.7~26.8℃之间,存在明显的冷暖变化,进一步揭示中全新世温度变化的细节;期间5860、5660和5160 a BP是3个显著的SST偏低期。
3)对比现代与中全新世(5427—5394、5243—5209、4515—4456 和4404—4356 a BP)珊瑚生长率的频谱周期,发现现代与中全新世珊瑚的生长率均存在显著3~7 a ENSO 周期,但中全新世珊瑚生长率的ENSO主周期改变且频率显著降低,表明中全新世ENSO活动总体偏弱;概率密度函数统计显示,中全新世期间ENSO 变率存在逐渐增强的趋势。
致谢:陈雪霏副研究员提供了海南龙湾现代珊瑚的生长率数据,党少华博士研究生帮助测量珊瑚生长率和分析数据,审稿专家和编辑对论文提出了宝贵的修改意见。谨表谢忱!
