荸荠球茎膨大相关基因COL5的克隆及表达分析
2023-11-11赵若男玉万国陈振林宋慕波刘英健
赵若男,玉万国,陈振林,宋慕波,刘英健*
1. 贺州学院食品科学与工程技术研究院,广西贺州 542899;2. 广西科技大学生物与化学工程学院,广西柳州 545616
荸荠(Eleocharis dulcis)也称马蹄,是禾本目莎草科荸荠属的一种浅水植物[1]。作为中国特色水生蔬菜,荸荠在广西、福建、海南等地均有大面积种植[2]。荸荠的球茎与莲藕的根茎、马铃薯的块茎、洋葱的鳞茎等都属于地下变态茎[3]。膨大的荸荠球茎可供食用,口感清脆、汁多味甜,富含碳水化合物、蛋白质、维生素等多种营养物质,具有很高的营养价值[1,4]。荸荠球茎的膨大过程是一个复杂的发育过程,需要经历4个阶段[5]。球茎膨大的第一个阶段是匍匐茎阶段,此阶段球茎顶端开始膨大,淀粉积累较少;之后是膨大初期,此时球茎开始生长,淀粉逐渐积累;接下来是膨大中期,球茎生长速度减缓;最后是膨大后期,此阶段球茎生长速度又加快,淀粉快速积累[6-8]。球茎的发育直接关系到荸荠的产量。因此,开展荸荠球茎膨大的分子机理研究对优化荸荠育种有重要意义。
近期研究中发现CONSTANS-Like(COL)基因家族成员可能在变态茎发育过程中起主要的调控作用。COL属于锌指蛋白转录因子家族,已知其家族成员在植物生长发育过程中扮演重要角色[9-10]。COL基因包含2~3个保守结构域,靠近N端有1个或者2个B-box Zinc finger结构域,另外一个是靠近C端的由约43个氨基酸组成的CCT保守结构域[5,11]。目前,COL基因家族已经在许多物种中相继报道。在拟南芥中发现了17个COL基因家族成员[12],在水稻(Oryza sativa)中至少有16个[13],大麦(Hordeum vulgare)中有9个[13],烟草(Nicotiana tabacum)中有15个[14],杜仲(Eucommia ulmoides)有8个COL基因[9]。以往研究发现,COL基因家族不同成员的功能各不相同。在拟南芥中的一系列研究发现,AtCOL4通过脱落酸依赖性信号参与植物非生物胁迫反应,增强植物的耐受性[15]。过表达AtCOL9可以延迟拟南芥开花[16]。AtCOL3是侧根发育和形成的正调控因子,另外在短日光条件下,抑制芽的伸长,促进分支芽的形成[17]。AtCOL7在拟南芥分支形成和下胚轴的伸长中发挥重要作用[18]。烟草中有3个COL基因在低温环境中表现出较大的差异,NtCOL13b在叶片中有较高的表达量,而NtCOL16c和NtCOL16d刚好相反[14]。最近研究发现,COL基因在植物贮藏器官发育中扮演着重要角色。莲藕NnCOL5基因参与根茎的膨大,尤其在根茎膨大中期(S3时期)表达量最高,并且在马铃薯中过表达NnCOL5可明显促进块茎单薯重和淀粉显著升高[19]。通过反义基因技术证实马铃薯(Solanum tuberosum)中的1个COL家族成员在块茎膨大发育过程中起负调控作用[20]。虽然在部分植物变态茎的发育过程中证实了COL基因的重要作用,但荸荠球茎中仅对淀粉合成过程中关键酶编码基因进行了克隆[21-24],膨大过程中COL基因的作用尚未有相关研究。
鉴于此,本研究从前期转录组数据中筛选并克隆荸荠球茎膨大相关COL基因,并对该基因序列进行生物信息学分析,同时利用荧光定量PCR分析该基因在球茎膨大过程中以及在不同组织中的表达模式。为进一步研究COL家族基因在荸荠膨大这一过程的分子机制和基因功能研究提供理论参考。
1 材料与方法
1.1 材料
供试材料选取广西地方品种桂林荸荠,采自广西贺州市八步区荸荠种植田,根据荸荠球茎不同发育时期(S1~S4时期,球茎最宽处直径分别约为10、20、35、50 mm)进行取样。同时采集球茎膨大后期的根、叶片、荸荠皮和荸荠肉样品。液氮速冻于-80 ℃超低温冰箱备用。植物RNA提取试剂盒购自华越洋生物科技(北京)有限公司;植物基因组DNA提取试剂盒、DNA凝胶回收试剂盒购自天根生化科技(北京)有限公司;反转录试剂盒、载体PMD18-T、DH5α大肠杆菌感受态均购自TaKaRa公司。
1.2 方法
1.2.1 荸荠基因组DNA、总RNA和cDNA的合成 从-80 ℃超低温冰箱中取出步骤1.1所储存的样品,按照植物基因组DNA提取试剂盒的使用说明书分别进行荸荠样品DNA、总RNA的提取。用超微量紫外分光光度计检测总RNA的纯度和含量。对合格的RNA进行反转录从而获得cDNA第一链,-20 ℃储存。
1.2.2 荸荠COL5基因的克隆 前期取荸荠叶、茎、根和球茎不同膨大时期的组织采用Pacbio平台进行全长转录组测序(Pacbio full-length cDNA sequencing),测序共获得154 398个转录本序列信息。在此荸荠转录组中筛选得到的CwCOL5基因序列信息设计引物(CwCOL5-F:5′-ATGGGAATAGAAAAAGGAGCCAAGT-3′,CwCOL5- R:5′-TCAAAATGTCGGCACCACACTGTAC-3′)。
分别以荸荠球茎组织的DNA、cDNA为模板进行PCR扩增,具体反应体系及反应程序见表1和表2。通过1%琼脂糖凝胶电泳对扩增产物进行检测,切胶,使用DNA凝胶回收试剂盒进行纯化回收,将回收产物连接至PMD18-T载体后转入DN5α大肠杆菌感受态细胞中,经过菌液PCR后,挑选阳性克隆进行送测。
表2 PCR扩增反应程序Tab. 2 PCR amplification reaction procedure
1.2.3 荸荠COL5基因生物信息学分析 生物信息学分析参考董伟清等[24]和宋慕波等[25]的方法进行。利用ORF finder、BLASTN、BLASTP、DNAMAN、ProtParam Tool、ExPaSy-SOPMA、SWISS-MODEL、SignalP 4.1 Server、Conserved Domains、TMHMM Server v.2.0以及NetPhos 2.0 Server软件进行生物信息学分析;利用ClustaW 1.83和Mega 5软件构建系统进化树。
1.2.4 荸荠COL5基因实时荧光定量PCR(RT-qPCR)分析 依据获得的CwCOL5基因序列设计荧光定量引物(CwCOL5-F: 5′-ACGCTAAGTCTGGCTATAGCTCTCT-3′,CwCOL5-R: (5′-AT GCATAGCGGATTGTCTTCTC-3′),该引物位于COL家族序列保守度低的区域,以避免该家族其它成员的干扰。以荸荠不同膨大时期(S1~S4时期)和不同组织(荸荠皮、荸荠根、荸荠肉和荸荠叶片)的cDNA为模板,以18s rRNA(登录号:MG742686)为内参,进行RT-qPCR扩增。测定方法参考何芳练等[22]的方法进行,采用2-ΔΔCT法计算相对表达量。
1.3 数据处理
采用Microsoft Office 2019软件进行数据处理和分析,使用Origin 2018和Adobe Photoshop 2021软件进行图片处理及美化。
2 结果与分析
2.1 荸荠COL5基因的克隆
在荸荠转录组数据库中筛选被注释为COL家族成员的转录本片段,其中编号为isoform_7928的转录本预测蛋白与同为莎草科的蒿草(Carex littledalei)COL家族成员同源性高达87%。在NCBI中的序列比对结果显示,该基因片段与其他物种的COL基因核苷酸序列同源性较高,其中与油棕(Elaeis guineensis)和生姜(Zingiber officinale)的核苷酸同源性分别为78.82%、77.97%。以该片段设计特异性引物,分别以荸荠球茎组织的DNA、cDNA为模板,通过PCR扩增克隆分别得到长度约为1000 bp和1200 bp的片段(图1)。经回收测序结果表明,以cDNA为模板扩增得到的cDNA片段长为1017 bp,编码338个氨基酸(图2),将其命名为CwCOL5,登录号为ON934922。以荸荠基因组DNA为模板扩增得到的DNA片段长为1275 bp,其含有1个长度为257 bp的内含子(图3)。

图1 CwCOL5基因PCR扩增产物电泳图Fig. 1 Electrophoresis of PCR amplification products of CwCOL5 gene

图2 CwCOL5基因ORF序列及其编码的氨基酸序列Fig. 2 CwCOL5 gene ORF sequence and its encoded amino acid sequence

图3 CwCOL5基因结构图Fig. 3 Structure diagram of the CwCOL5 gene
2.2 CwCOL5基因的生物信息学分析
2.2.1 CwCOL5蛋白理化性质分析 利用ExPaSy-SOPMA软件对CwCOL5氨基酸序列的理化性质进行预测。结果表明,CwCOL5蛋白质由338个氨基酸组成,其中丙氨酸(Ala)和丝氨酸(Ser)占比最高,分别为9.8%和9.5%,其次是精氨酸(Arg),为8.3%,不含吡咯赖氨酸(Pyl)、晒半胱氨酸(Sec)。分子式为C1592H2515N477O508S17,总原子数5109,理论等电点为5.89,相对分子质量为37 010.39,有43个带正电荷(Asp+Lys)的氨基酸残基,有48个带负电荷(Asp+Glu)的氨基酸残基。亲疏水性分析发现(图4),CwCOL5蛋白质第218位氨基酸残基疏水性最强(Score=1.467),第279位氨基酸残基亲水性最强(Score=-3.789),总平均亲水值(GRAVY)为-0.462,不稳定系数为45.63,推测为亲水性不稳定蛋白质。
2.2.2 CwCOL5蛋白质结构预测分析 利用Plant-mPLoc server在线软件对CwCOL5蛋白质序列进行亚细胞定位分析,发现该蛋白定位于细胞核。对CwCOL5蛋白质进行磷酸化位点预测,结果发现,该蛋白有31个磷酸化位点,丝氨酸(Ser)23个,苏氨酸(Thr)5个,酪氨酸(Tyr)3个,其中第292位丝氨酸预测得分最高,为0.996。利用SignalP 4.1 Server软件预测蛋白质信号肽,并利用TMHMM Server v.2.0软件预测蛋白质跨膜结构。结果显示,CwCOL5编码的蛋白质无信号肽,且不含跨膜结构域。
CwCOL5蛋白质的二级结构显示,该蛋白具有113个(33.43%)α-螺旋、13个(3.85%)β-折叠、178个(52.66%)无规则卷曲和34个(10.06%)延伸链。使用Swiss-Model软件对CwCOL5蛋白的三级结构进行同源建模,以系统自动匹配度最高的7wsj.2.A为参考建模时发现,其三级结构模型中无规则卷曲所占比例最高,与二级结构预测结果一致。
2.2.3 CwCOL5保守结构域分析 将CwCOL5基因编码的蛋白氨基酸序列提交至Pfam网站在NCBI进行蛋白结构域预测,结果发现(图5),CwCOL5编码的蛋白序列有2个高度保守的B-box Zinc finger和1个CCT结构域。从结构域位置来分析,CCT结构域位于CwCOL5蛋白的C端,基序长为43个氨基酸,2个B-box Zinc finger位于N端且紧密相邻。

图5 CwCOL5蛋白结构域预测Fig. 5 CwCOL5 protein domain prediction
2.2.4 多序列比对分析和系统进化树构建 使用GENEDOC对CwCOL5蛋白质序列同在NCBI数据库中下载的19个其他植物COL蛋白质序列进行多序列比对分析(图6)。结果中黑色部分表示蛋白序列同源性最高,其次是深灰色、浅灰色。荸荠COL5与其他植物同源蛋白氨基酸有较高的一致性,在C端和N端的序列都相对保守。采用MEGA 5.0软件中的Neighbor-Joining法构建CwCOL5氨基酸序列与拟南芥、烟草、番茄(Solanum lycopersicum)、高粱(Sorghum bicolor)、小米(Setaria italica)、大麦等19个物种COL氨基酸序列的系统进化树(图7),结果发现,依据拟南芥COL蛋白可以分为2个不同的分支,其中荸荠CwCOL5蛋白与ATCOL5等物种COL蛋白亲缘关系较近。

图6 CwCOL5氨基酸序列比对Fig. 6 CwCOL5 amino acid sequence alignment
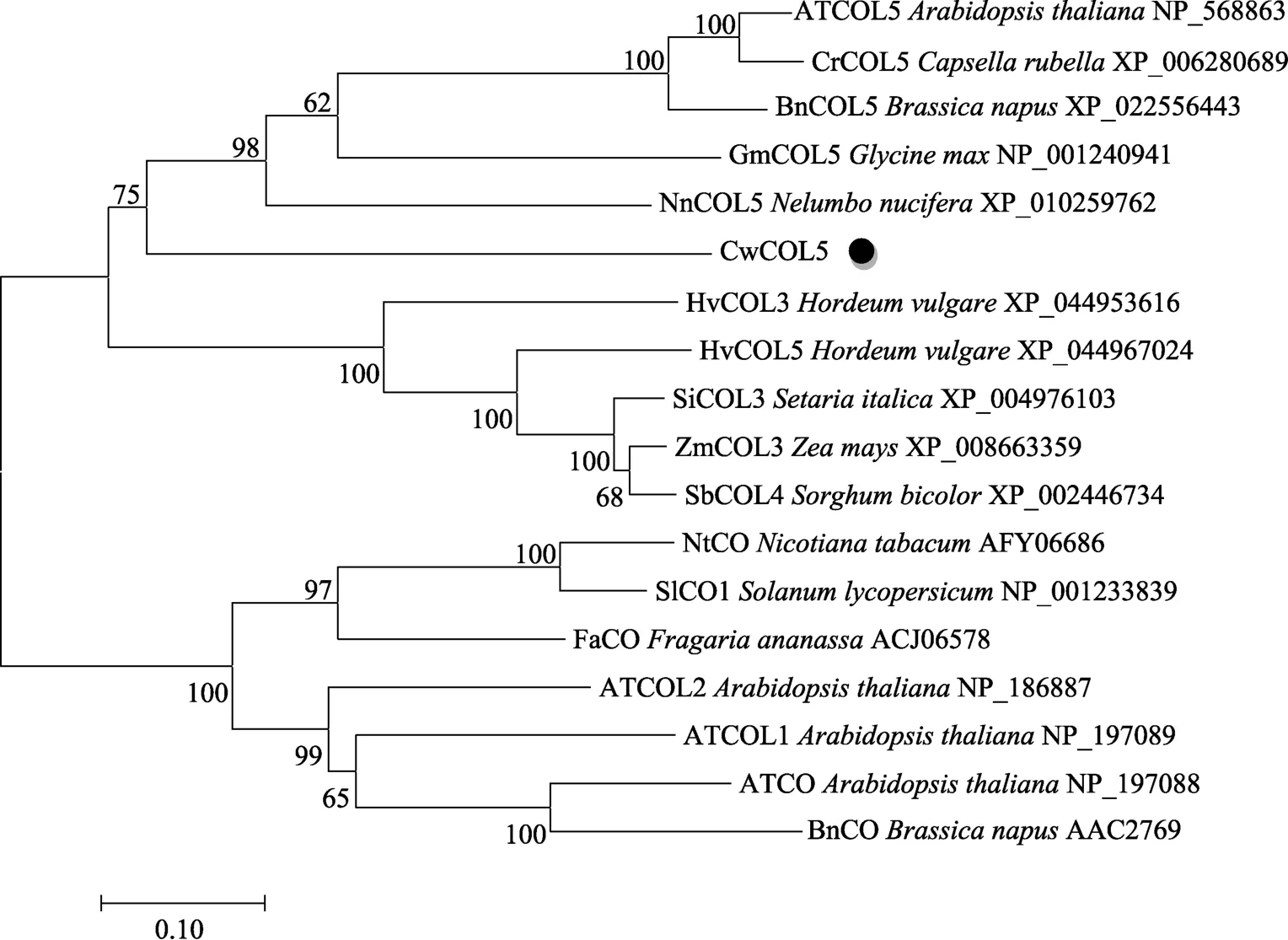
图7 CwCOL5蛋白系统发育进化树Fig. 7 CwCOL5 protein phylogenetic evolutionary tree
2.3 CwCOL5基因在球茎膨大过程中的表达分析
为了解CwCOL5基因在荸荠球茎膨大4个时期的表达情况,根据序列结果设计引物进行RT-qPCR检测,结果显示,CwCOL5基因S4时期的表达量显著低于S1、S2、S3时期(P<0.05),S3时期的表达量最高(图8)。

图8 CwCOL5基因在荸荠球茎膨大过程中的表达分析Fig. 8 Expression analysis of CwCOL5 gene during corm enlargement in Chinese water-chestnut
2.4 荸荠CwCOL5基因组织表达模式分析
由图9可知,CwCOL5基因在荸荠肉、根、叶片和荸荠皮中均有并表达,其中,在叶片和荸荠皮中的表达非常显著且表达水平相当(P<0.05),在荸荠肉和根部位表达水平相当。

图9 CwCOL5基因在荸荠不同组织中的表达Fig. 9 Expression of CwCOL5 gene in different tissues of Chinese water-chestnut
3 讨论
荸荠在我国长江流域以及广西、广东等南方各省(区)都有大面积种植,被广泛用于食品、医药等领域,具有较高的营养价值、生态价值和经济价值[1,4]。研究发现COL(CONSTANS-Like)基因是植物储存器官发育的重要调控基因[26-27]。然而,荸荠球茎作为一种典型的贮藏器官和变态茎,其球茎膨大调控机制的相关研究较少。本研究克隆得到可能参与荸荠球茎膨大调控的CwCOL5基因,该序列ORF长度为1017 bp,可编码338个氨基酸,其DNA序列长1275 bp,包含1个内含子。CwCOL5基因结构特征以及长度均与莲藕COL5和马铃薯COL相似[20,28]。
目前研究认为CO-FT通路在植物变态茎发育的调控中起重要作用,而在该通路中COL基因家族扮演重要角色[29-30]。在拟南芥中,COL基因家族蛋白根据其结构可分为3组,第I组包含2个B-box结构域和1个CCT结构域;第II组只包含1个B-box域和1个CCT域;而第III组包含1个B-box结构域、1个次级锌指蛋白结构域和1个CCT结构域[13,19,25]。CwCOL5与拟南芥COL基因家族第I组的结构相同。马铃薯COL蛋白包含2个B-box结构域和1个CCT结构域,也属于第I组[20]。COL蛋白在许多植物中参与光信号的感受与转导进而影响植物开花,例如过表达AtCOL5时,FT、SOC1相关开花基因表达上调,从而导致拟南芥可以在短时间内开花[31]。然而,有研究表明COL在马铃薯和莲藕中参与了马铃薯块茎膨大以及淀粉含量积累的调控[19]。因此,本课题组推测与AtCOL5和NnCOL5近缘的CwCOL5可能参与荸荠球茎的发育调控。
CwCOL5在荸荠不同组织和球茎不同发育阶段均有明显表达,CwCOL5在荸荠叶片和荸荠皮中表达量较高,在荸荠果肉中的表达量相对较低。马铃薯COL基因在叶片中的表达高于其他组织[32],水稻、大豆、杨树中的COL基因作用的主要部位也是在叶片[30,33-34],本研究发现CwCOL5的表达模式与上述结果大体一致。在球茎发育过程中CwCOL5在球茎膨大前期表达量快速上升,而在膨大后期显著下调,这一表达模式与马铃薯StCOL、莲藕NnCOL5相似[19,31]。基于上述结果,推测CwCOL5可能参与了荸荠球茎膨大过程,但球茎膨大过程存在复杂的调控机制,CwCOL5与CO-FT途径其它关键基因如FKF1、CDF1和SP6A等的相互影响有待进一步研究。
