不同水分条件天鹅湖高寒湿地土壤线虫群落结构对氮沉降的响应
2023-11-09陈康怡韩耀光王家力申志博杨贝贝赵一朱新萍贾宏涛
陈康怡,韩耀光,王家力,申志博,杨贝贝,赵一,朱新萍,2*,贾宏涛,3
(1.新疆农业大学资源与环境学院,乌鲁木齐 830052;2.北京农学院生物与资源环境学院,北京 102206;3.新疆土壤与植物生态过程重点实验室,乌鲁木齐 830052;4.新疆农业大学草业学院,乌鲁木齐 830052)
大气氮沉降及降水作为全球变化的重要现象,其格局的改变带来了一系列生态问题[1],湿地作为一种介于水生和陆地的特殊生态系统,对大气氮沉降以及水分变化等全球变化响应敏感[2]。氮沉降会通过影响土壤环境因子、微生物、植物生物量等,进而影响地下食物网的功能[3],对土壤动物产生直接或间接的影响[4]。
土壤线虫作为土壤动物的重要功能类群,处于土壤食物网的多个营养级[5],是碳氮等重要营养元素矿化的重要参与者和生态环境变化的重要指示生物,可用来评价土壤受干扰程度,其群落组成和结构的变化对维持湿地生态系统的地球化学循环和功能服务具有极其重要的意义[6]。近年来,土壤线虫在土壤生态环境中的作用日益受到重视,例如:在农业施肥对土壤线虫群落结构影响的研究中发现,减施化肥和配施黄腐酸后食细菌类线虫的数量和小麦各生育期平均丰度降低[7];草地生态系统土壤线虫群落的丰度随着放牧干扰强度的增加而增加[8];不同土地利用方式会对线虫群落有显著影响,自然农田线虫数量和种属数均高于退耕农田[9]。湿地生态系统作为生态系统的重要组成部分,由于气候变化、植被类型以及人为干扰等作用,土壤线虫群落组成、结构以及生态指数呈现不同的差异。在气候变化下,氮沉降增加和水分格局改变对我国干旱区巴音布鲁克高寒湿地同样产生一定的影响,据报道巴音布鲁克高寒草原氮沉降(以N 计)已达到8 kg·hm-2·a-1[10],该区域目前已开展了关于氮沉降及水分条件对巴音布鲁克高寒湿地土壤有机碳组分和温室气体的影响研究[11],然而在土壤线虫对氮沉降和水分条件的响应方面还处于空白。巴音布鲁克高寒湿地位于我国西北干旱区的新疆维吾尔自治区天山中部,具有独特的内陆高寒湿地生态系统特征。因此开展不同水分条件下氮添加对湿地土壤线虫群落结构特征影响的研究,有利于加深氮沉降增加对干旱区高寒湿地地下生态环境变化的认识,为科学评价氮沉降对高寒湿地土壤生态系统健康的干扰性及维持草地生态系统健康提供理论依据。
1 材料与方法
1.1 试验设计
在巴音布鲁克良好的原生稳定苔草湿地(42°40′~43°00′N,83°40′~84°35′E)选择2 个自然水分梯度作为研究区域:地表季节性积水条件区域(B),土壤体积含水率在43%~48%之间,植被优势种有黑花苔草(Carex melanantha)、水麦冬(Triglochin palustre);地表常年干燥条件区域(C),土壤体积含水率在30%~35%之间,植被优势种有冰草(Agropyron cristatum)、黑花苔草(Carex melanantha)[12]。本研究在每个水分条件区域选取植被盖度相对一致的3 块长7.5 m、宽7.5 m的正方形样地,围栏后进行氮添加试验。以全球草地平均氮沉降水平约为10 kg·hm-2·a-1[13]为氮添加量依据,设置3个氮添加处理(以N 计):N0(不添加氮),N10(添加氮10 kg·hm-2·a-1),N20(添加氮20 kg·hm-2·a-1)。每处理3个重复。氮素由硝酸铵(NH4NO3)和尿素[CO(NH2)2]按照7∶3的比例混合而成[14],以喷洒方式于2020 年6 月和9 月分两次进行添加,添加氮素后次年5月对不同氮处理土壤进行采样。
1.2 样品采集与测定方法
1.2.1 土壤样品采集
每个处理随机采3个土壤混合样,取样深度为0~5、5~10、10~20 cm。土壤样品尽量去除石头和可见根后混合均匀,分成2 份带回实验室。1 份用于分离土壤线虫(4 ℃冰箱保存)和测定含水量,1 份土壤自然风干后研磨,过1 mm 和0.25 mm 网筛,用于土壤基本理化性质的测定。
1.2.2 土壤环境因子测定
参考《土壤农业化学分析方法》[15],pH 使用pH 计测定;电导率(EC)使用电导率仪测定;含水量(SM)采用烘干法测定;碱解氮(AN)采用碱解扩散法测定;土壤有机碳(SOC)采用浓硫酸-重铬酸钾外加热法测定;全氮(TN)和全碳(TC)采用元素分析仪(EA3000EA3000,Euro Vector,Italy)测定。
1.2.3 土壤线虫的分离与鉴定
取土壤鲜样50 g,利用改良浅盘分离-蔗糖离心浮选的连续提取法[16-17]进行线虫分离,将分离静置后的线虫溶液中加入固定液(9%福尔马林、1%甘油)保存,通过体视显微镜进行计数,根据土壤含水率计算每100 g干土中含有的线虫数量。将分离出的线虫随机选取200 条制成临时玻片,参照土壤线虫分类书籍[18-19],利用形态学方法,使用生物显微镜鉴定到科属。根据线虫取食类型的不同将其分为食细菌类群(BA,Bacterivores)、食真菌类群(FU,Fungivores)、植物寄生类群(PP,Plant-parasites)、杂食/捕食类群(OP,Omnivores/Predators)[20]。
1.2.4 土壤线虫群落生态指数计算
土壤线虫群落生态指数根据以下公式计算[21]:
(1)香农威尔指数(H)
式中:pi为第i个分类单元中线虫个体占总个体数量的比例。
(2)优势度指数(λ)
(3)成熟度指数(MI)
式中:vi为根据自由生活线虫在生态演替中的不同生活策略分别赋予的c-p 值;fi为某一科/属线虫(i)在自由生活线虫(不包括植物寄生类线虫)总数中所占的比例。
(4)植物寄生类线虫的成熟度指数(PPI)
式中:PP为寄生类线虫数量。
(5)富集指数(EI)
式中:e代表食物网中的富集成分,主要指食细菌线虫中c-p 值为1 和食真菌类线虫中c-p 值为2 的类群;b代表食物网中的基础成分,主要指食细菌类线虫和食真菌类线虫中c-p值为2的类群。
(6)结构指数(SI)
式中:s代表食物网中的结构成分类群,分别为食细菌类线虫、食真菌类线虫和植物寄生类线虫中c-p值为3~5 的类群以及杂食/捕食类线虫中c-p 值为2~5 的类群。
(7)线虫通道指数(NCR)
式中:BA和FU分别代表食细菌类线虫和食真菌类线虫的数量。
1.3 数据处理
使用Origin 2018 和R 4.1.1 制图,用SPSS 26.0 和Excel 2019 软件对数据进行方差分析。采用单因素方差分析法研究氮添加对土壤线虫群落的影响、氮添加对土壤环境因子的影响。采用冗余分析(RDA)和蒙特卡洛检验估算各土壤环境因子对线虫群落的影响。统计分析及图表制作于Excel、Origin 和R Studio 中完成。
2 结果与分析
2.1 氮添加对高寒湿地土壤理化因子的影响
如表1所示,地表季节性积水条件土壤含水量、有机碳和全碳均显著高于地表常年干燥条件土壤(P<0.05)。湿地土壤pH 均呈碱性(7.73~8.33),地表常年干燥条件土壤pH高于地表季节性积水条件土壤。地表季节性积水条件下N20 处理电导率显著高于N10和地表常年干燥条件土壤各氮处理(P<0.05)。地表季节性积水条件N20 处理的土壤有机碳含量显著低于N0 和N10 处理。相同水分条件下,氮添加对湿地土壤pH、全氮和全碳均无显著影响,但显著增加土壤碱解氮含量(P<0.05)。
2.2 氮添加对高寒湿地土壤线虫数量的影响
如图1 所示,地表常年干燥条件土壤线虫在100 g 风干土中的数量显著高于地表季节性积水条件土壤(P<0.05)。地表常年干燥条件土壤线虫主要分布在0~10 cm 土层中;地表季节性积水条件土壤线虫主要分布在0~5 cm 土层中,且氮添加处理土壤线虫数量显著低于N0 处理(P<0.05)。相同水分条件和施氮处理下,随土层深度的增加,土壤线虫的数量逐渐减少。

图1 土壤线虫个体密度的垂直分布Figure 1 Vertical distribution of individual density of soil nematodes
2.3 氮添加对高寒湿地土壤线虫群落结构的影响
所有处理共分离线虫9 737 条,线虫个体密度平均为每100 g 干土1 061 条,分属39 属,其中食细菌类线虫10 属,食真菌类线虫5 属,植物寄生类线虫10属,杂食/捕食类线虫14 属。地表季节性积水土壤线虫优势属是前矛线属(Prodorylamus),占比为29.30%,除此之外N0 处理土壤线虫优势属还有Laimydorus(21.53%)和真头叶属(Eucephalobus,11.19%),N10 处理的优势属还有Laimydorus(20.89%)和矛线属(Dorylaimus,13.24%)。随氮添加量的增加前矛线属丰度逐渐增加,在N0、N10 和N20处理分别达到了21.61%、23.03%和43.26%。地表常年干燥条件土壤线虫优势属是前矛线属(18.69%)、Laimydorus(20.21%)和螺旋属(Helicotylenchus,33.48%)。
在属水平上,相同水分条件的不同氮添加量下,土壤线虫群落组成具有相似性,但在丰度上具有差异(图2)。地表季节性积水条件下,N20处理与N0处理比较,前矛线属丰度上升了21.65%;Laimydorus和真头叶属丰度分别下降了12.5%和9.81%,并从优势属转变为常见属;地表常年干燥条件下,随着氮添加量的增大前矛线属和Laimydorus丰度分别上升了2.19%和12.28%。
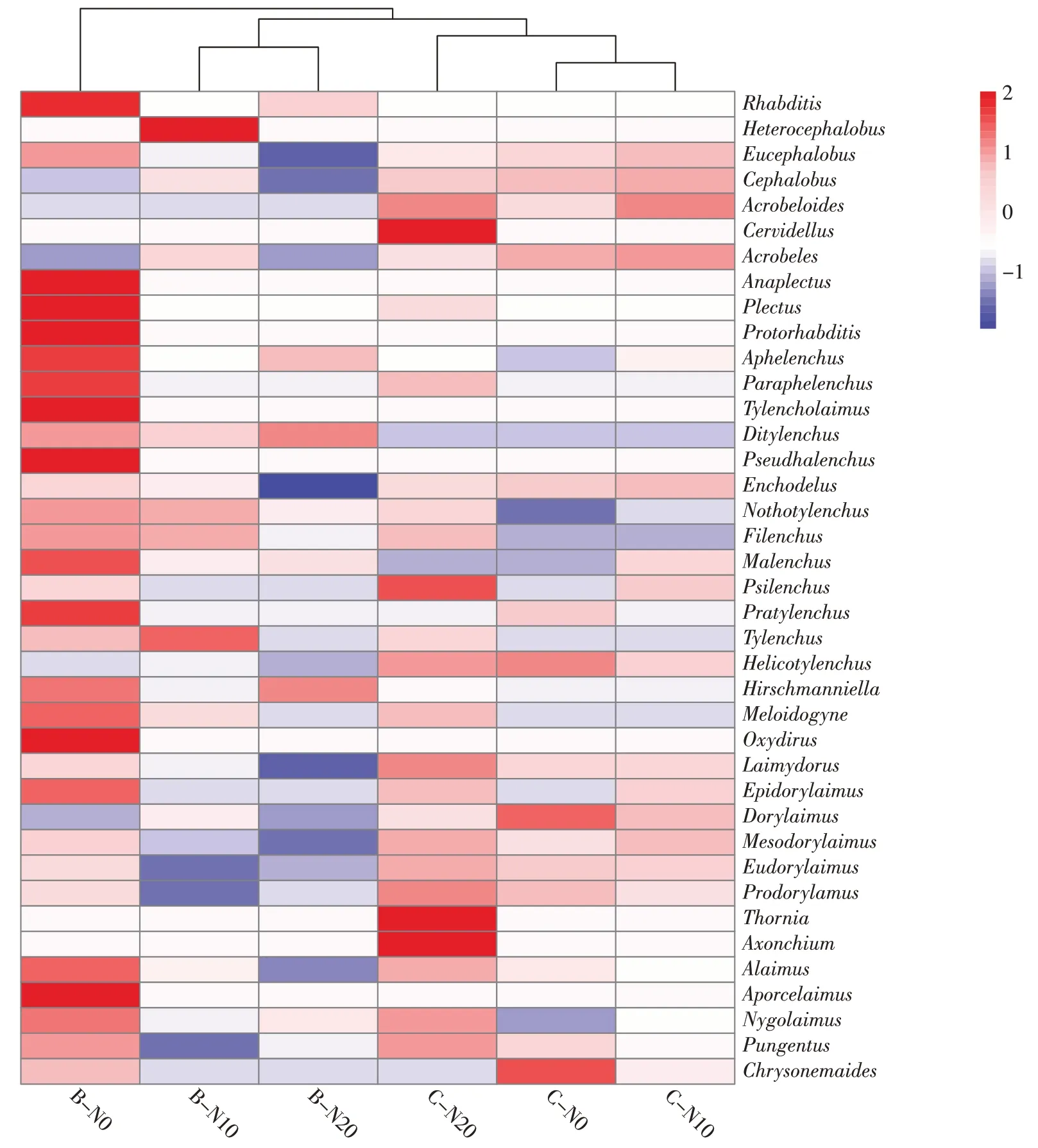
图2 氮添加对不同水分条件高寒湿地土壤线虫群落结构的影响Figure 2 Effects of nitrogen additions on soil nematode community structure in alpine wetland under different water conditions
水分条件和氮添加对土壤线虫营养类群分布特征具有一定影响。由图3 可知,地表季节性积水条件下,土壤线虫营养类群主要为植物寄生类线虫(PP)和杂食/捕食类线虫(OP),食真菌类线虫(FU)占比最小(4.19%~8.19%)。随着土层深度的增加,植物寄生类线虫在10~20 cm 土层比0~5 cm 土层上升了57.01个百分点,杂食/捕食类线虫占比在10~20 cm 比表层降低了46.75个百分点,食细菌类线虫(BA)占比逐渐下降。地表常年干燥条件土壤线虫营养类群分布特征与地表季节性积水条件类似,但杂食/捕食类线虫和食真菌类线虫占比低于地表季节性积水条件,而植物寄生类线虫占比高于地表季节性积水条件。

图3 不同水分条件下土壤线虫营养类群空间分布特征Figure 3 Spatial distribution characteristics of soil nematode communities with different trophic groups under different water conditions
由图4 可见,不同水分条件土壤下,氮添加量对土壤线虫营养类群产生一定影响。在地表季节性积水条件下,随着氮含量上升,土壤杂食/捕食类线虫占比N20比N0处理上升了11.32个百分点,这主要是由前矛线属等丰度上升造成的。真头叶属等食细菌类线虫占比迅速下降,N20 比N0 处理下降了13.32 个百分点;植物寄生类线虫和食真菌类线虫占比变化不明显。地表常年干燥条件下,N20 较N0 处理杂食/捕食类线虫占比上升了17.29 个百分点,这主要是由前矛线属和Laimydorus 等丰度上升造成的。与N0 比较,N10处理下植物寄生类线虫(PP)的占比显著下降,其中螺旋属显著下降了29.1 个百分点。食细菌类线虫占比有所增加,食真菌类线虫占比变化不明显。
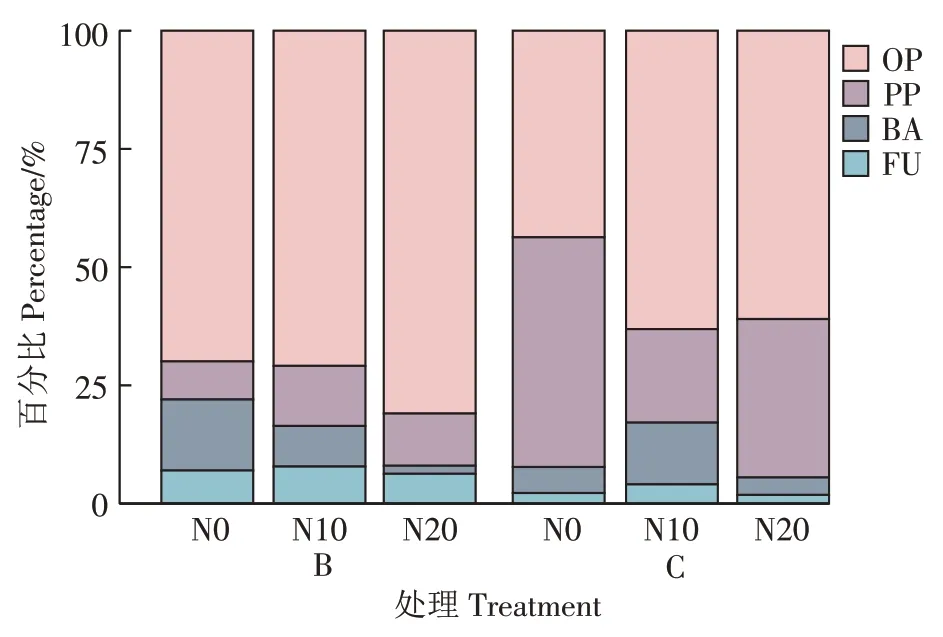
图4 不同氮添加量下土壤线虫营养类群分布特征Figure 4 Distribution characteristics of soil nematode communities with different trophic groups under different nitrogen additions
2.4 氮添加对高寒湿地土壤线虫群落生态指数的影响
由表2 可知不同水分条件下,氮添加增大了MI指数,对土壤线虫多样性无显著影响,地表季节性积水条件的H指数和λ指数均高于地表常年干燥条件,且不同氮添加处理间差异不显著。地表常年干燥条件下,NCR趋近于1,显示该水分条件土壤有机质分解途径主要是细菌分解。地表季节性积水条件下,随着氮添加量上升,EI指数和PPI指数先下降后上升;N20 处理NCR指数为0.15,说明土壤有机质分解途径主要为真菌分解。EI小于50 但SI大于50,说明不同水分条件下氮添加对土壤环境无明显干扰,养分富集状况中等。
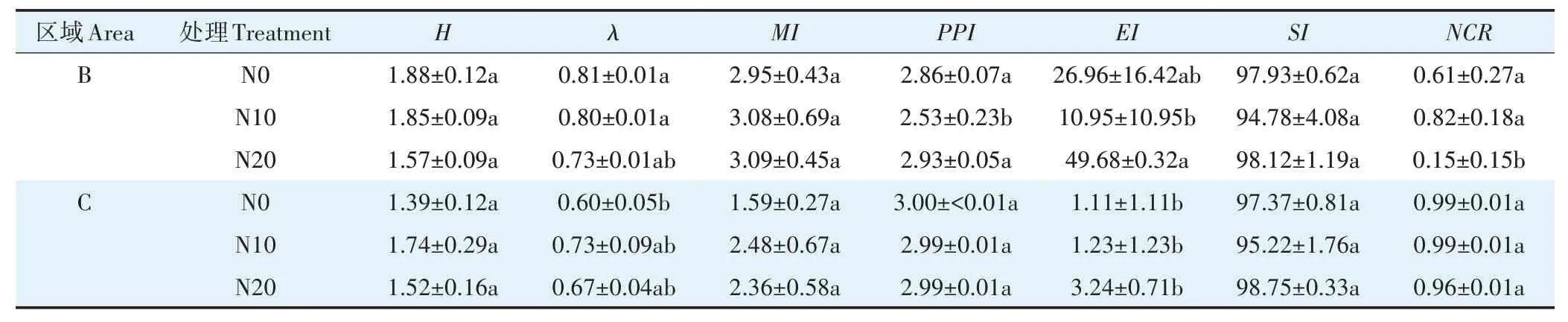
表2 氮添加对高寒湿地土壤线虫群落生态指数的影响Table 2 Effects of nitrogen addition on the ecological index of soil nematode communities in alpine wetlands
2.5 氮添加下高寒湿地土壤环境因子和土壤线虫群落之间的关系
图5、图6 是不同水分条件下氮添加对土壤环境因子与土壤线虫群落常见属的联系进行冗余分析后得到的RDA 排序图。环境因子用黑色箭头表示,线虫属用蓝色箭头表示,样本分组用不同色块表示。箭头延申越长代表环境因子与线虫属相关性越大,反之越小。箭头延长线和排序轴夹角代表土壤环境因子与排序轴相关性大小,夹角越小相关性越高,反之越低。
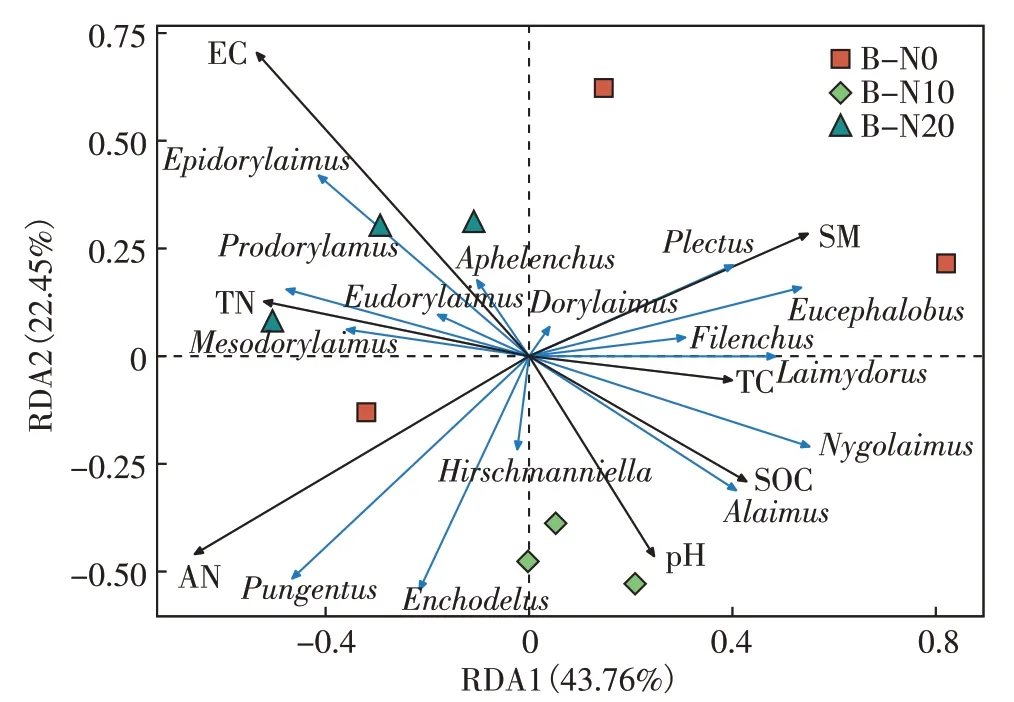
图5 地表季节性积水条件土壤理化因子和土壤线虫群落数量的RDA分析Figure 5 RDA analysis of soil physicochemical factors and soil nematode community quantity under surface seasonal waterlogging conditions
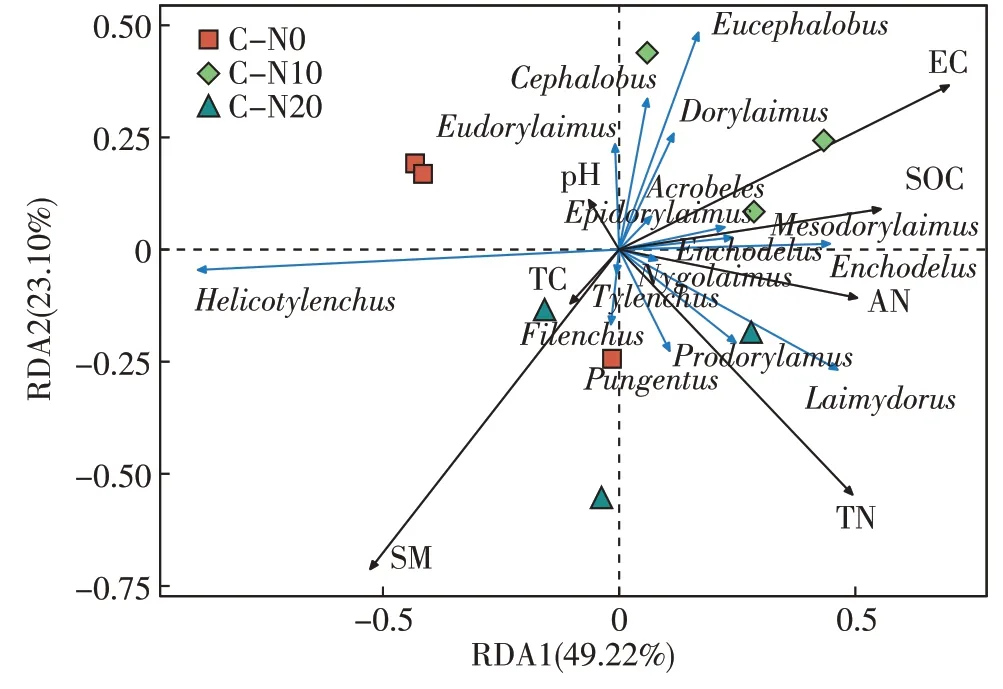
图6 地表常年干燥条件土壤理化因子和土壤线虫群落数量的RDA分析Figure 6 RDA analysis of soil physicochemical factors and soil nematode community quantity under perennial dry conditions
地表季节性积水条件的典型轴1 的解释量为43.76%,典型轴2 解释量为22.45%。根据箭头延伸长、夹角小的特点可知,土壤线虫Epidorylaimus、真滑刃属(Aphelenchus)与EC有较大相关性;螯属(Pungentus)、垫咽属(Tylencholaimus)分别与AN、TC 和SOC 有较强相关性;拟绕线属(Plectus)、丝尾垫刃属(Filenchus)和真头叶属与SM相关性较高;前矛线属、中矛线属(Mesodorylaimus)和真矛线属(Eudorylaimus)与pH和TN相关性较高。由蒙特卡洛置换检验的结果可知(表3),EC和AN 与土壤线虫群落显著相关(P<0.05),是影响线虫群落结构变化的主要驱动因子。
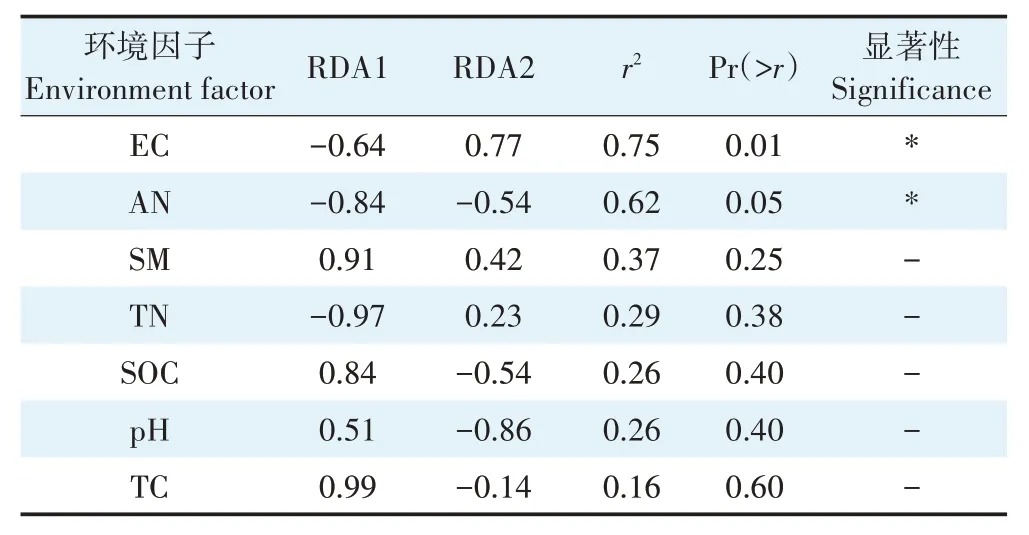
表3 地表季节性积水条件土壤理化因子和土壤线虫群落的蒙特卡洛检验Table 3 Monte Carlo test of soil physicochemical factors and soil nematode communities under surface seasonal waterlogging conditions
地表常年干燥条件的典型轴1 的解释量为49.22%,典型轴2解释量为23.10%。土壤线虫螺旋属与SM 有较强相关性;前矛线属、穿咽属(Nygolaimus)、螯属分别与pH 和AN 相关性较高;矛线属、真矛线属、头叶属(Cephalobus)、丽突属(Acrobeles)分别与TN、TC 和SOC 的相关性较高。由蒙特卡洛置换检验的结果可知(表4),线虫群落结构的变化受SM 的显著影响(P<0.05),SM 是影响线虫群落结构变化的主要驱动因子。

表4 地表常年干燥条件土壤理化因子和土壤线虫群落的蒙特卡洛检验Table 4 Monte Carlo test of soil physicochemical factors and soil nematode communities under perennial dry conditions
为继续探究土壤线虫生态指数与土壤环境因子的相关性,对二者进行相关性冗余分析得到图7。SI指数、PPI指数与pH 在同一象限具有相关性,MI指数与TN、AN、TC、SOC 和SM 有较强相关性。环境因子用黑色箭头表示,线虫生态指数用蓝色箭头表示,样本分组用不同色块表示。由蒙特卡洛置换检验的结果可知(表5),pH 极显著影响土壤线虫生态指数(P<0.01),SM和TC显著影响线虫生态指数(P<0.05),其他环境因子的影响不显著。
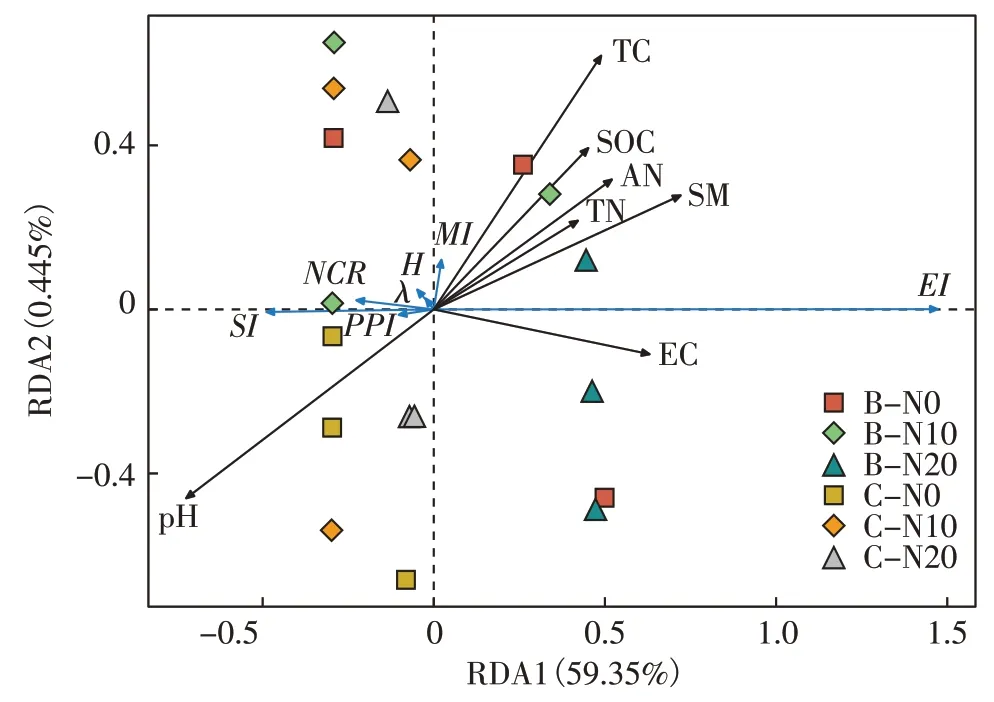
图7 土壤理化因子和土壤线虫群落生态指数的RDA分析Figure 7 RDA analysis of soil physicochemical factors and soil nematode community ecological index
3 讨论
3.1 水分条件与氮添加对土壤线虫群落结构的影响
土壤线虫群落的变化被认为是环境条件变化的标志,群落组成和结构变化对维持湿地生态系统的地球化学循环和功能服务具有极其重要的意义[6]。土壤线虫丰度对土壤含水量响应敏感[22],从土壤线虫数量上看,巴音布鲁克高寒湿地不同水分条件下不同氮添加量的样地中共获得土壤线虫9 737 条,线虫个体密度平均为每100 g 干土1 061 条,分属39 属。地表常年干燥条件土壤线虫每100 g风干土中的数量显著高于地表季节性积水条件土壤(P<0.05),这与杨贝贝[8]的研究不符,可能与采样季节和土壤水分有关,本研究采样在5 月,湿地处于冰雪融化时期,植物才开始生长,土壤水分增加的同时可能导致溶解氧减少[23],从而会对土壤线虫中垫刃属、某些矛线属等植物寄生性线虫的生长产生抑制[24],影响土壤线虫在土壤空隙中的运动及捕食情况,引起线虫数量改变[25]。
水分和氮素影响土壤线虫群落的不同营养类群,从而造成线虫生态指数变化,同时,生态指数可以反映水分和氮沉降对土壤线虫群落结构、多样性及土壤食物网稳定性的影响。季节性积水条件土壤中杂食/捕食类线虫和食真菌类线虫的群落占比要高于常年干燥条件土壤,而植物寄生类线虫则相反,这主要是因为随着土壤含水量增加,食真菌类线虫数量上升,说明土壤水分上升可以增加食真菌线虫数量[26]。随氮含量的增加,土壤食细菌类线虫数量下降,因而表现出NCR值接近0,土壤有机质的真菌分解途径占比增大[27],氮添加改变了土壤环境,抑制了微生物生长和有机质的转化,降低了能量流动速度[28-29]。杂食/捕食类群线虫体型较大、生活史长、卵量小,一般以其他营养类群线虫为食物来源[21],作为典型k-对策者c-p 4~5 的类群,其对环境变化和胁迫极为敏感。因此,当食真菌类线虫数量上升时,杂食/捕食类线虫占比上升。植物寄生类线虫比例在地表常年干燥条件土壤中显著高于季节性积水条件,这与前人研究一致[8],说明土壤线虫数量只有在水分适宜的条件下才相对较高,而并非含水量越高土壤线虫数量越大。线虫数量表聚性体现在随土壤深度增加,土壤线虫数量急剧降低[30],不同氮添加水平下,这种趋势没有改变,但氮添加量对土壤线虫营养类群会产生一定影响。
富集指数用于预测食物网对可利用资源的响应,结构指数表征干扰及生态恢复过程中土壤食物网结构的变化,两者结合可指示土壤环境受干扰程度和土壤食物网状况[1]。杂食/捕食类线虫作为对环境干扰最敏感的营养类群[31],随着氮素的增加占比上升,同时富集指数小于50 但结构指数大于50,说明在相同水分条件下,氮添加使土壤养分中等程度富集,食物网呈结构化,土壤生态系统稳定性逐渐增强[32],有利于杂食/捕食类线虫的生长繁殖。香农威尔指数侧重于稀有属,优势度指数侧重于优势属,二者共同反映出土壤线虫的多样性。多样性指数较高时,说明群落中物种较多且分布均匀,反之物种较少且分布不均匀[33]。随着氮含量上升,食细菌类线虫占比迅速下降,氮素添加导致土壤中和浓度增加,随着氮浓度升高对土壤线虫产生毒害作用[34]。氮添加对食真菌类线虫占比影响不明显,这与张爱林的研究一致[35]。因此,地表季节性积水区多样性指数下降,但不同处理间不具有显著差异,氮添加降低了线虫属分布均匀程度。季节性积水条件线虫群落多样性比地表常年干燥条件较好。成熟度指数根据土壤线虫群落受环境干扰后,经过一段时间恢复所处的演替阶段来反映土壤环境受干扰程度[17]。氮添加增大了成熟度指数,这表明氮添加可以减缓土壤环境的干扰程度,促进土壤生态系统向成熟稳定发展,有利于增加土壤食物网的稳定性和复杂性。
3.2 土壤线虫群落与土壤环境因子间的关系
土壤环境因子的改变是影响土壤线虫生存的重要驱动因素之一[36]。季节性积水条件土壤pH 低于常年干燥条件土壤,氮添加使土壤pH值降低,对土壤中微生物种群数量造成影响,继而影响土壤线虫群落[29,37]。氮添加影响了土壤电导率、土壤含水量和碱解氮含量。在地表常年干燥条件下,蒙特卡洛检验结果显示,土壤线虫群落变化主要受土壤含水量影响。当土壤水分增加时,土壤线虫群落受电导率和碱解氮影响显著(P<0.05),说明在高寒湿地生态系统中,水分比施氮更能影响土壤线虫群落数量;缺水环境会一定程度降低施氮的影响[1],当土壤水分增加到一定范围,氮素添加对线虫群落的直接或间接影响会逐步显现,尤其是对土壤电导率和碱解氮的影响。从c-p 类群来看,r-对策者的c-p 2 类群世代生活史短,卵量大,耐环境污染和胁迫,在氮添加下变化不明显。而作为k-对策者的c-p 4~5 类群因对环境胁迫敏感,受pH、电导率和土壤含水量的影响远大于土壤养分的影响,因此在氮添加改变土壤环境的情况下k-对策者的c-p 4~5类群迅速减少[38]。地表季节性积水条件土壤线虫香农威尔指数与电导率之间存在显著负相关(P<0.05),说明电导率增加可能会降低土壤线虫多样性,杂食/捕食类线虫的主要种属与土壤pH、碱解氮有较强相关性,表明这些土壤环境因子可能是杂食/捕食类线虫丰度变化的原因。
4 结论
(1)地表季节性积水条件土壤线虫的优势属主要为前矛线属(Prodorylamus),地表常年干燥条件土壤线虫优势属是前矛线属(Prodorylamus)、Laimydorus和螺旋属(Helicotylenchus)。
(2)土壤线虫在土壤中呈现一定的表聚性分布特征,随土层深度增加丰度下降。土壤水分增加,土壤杂食/捕食类线虫和食真菌类线虫丰度升高,植物寄生类线虫丰度降低,氮添加会增加杂食/捕食类线虫丰度。同时促进湿地土壤有机质分解途径由细菌向真菌分解途径发展,有利于增加土壤食物网的稳定性和复杂性。
(3)水分和氮素共同作用下,土壤含水量、土壤电导率和碱解氮成为影响土壤线虫群落结构的主要因素。
