中亚热带赤皮青冈天然种群表型多样性分析
2023-11-01秦之旷刘娜周霞李志辉曹福亮李何
秦之旷 刘娜 周霞 李志辉 曹福亮 李何
摘 要:赤皮青岡(Quercus gilva)是我国中亚热带地区极具经济价值的珍贵用材树种。为揭示其表型多样性与变异规律以及影响表型变异的关键地理气候因子,该研究以14个赤皮青冈天然种群115个单株为材料,对其15个树体和叶片性状进行测量,并采用巢式方差分析、表型分化系数分析、多样性指数分析、相关性分析、主成分分析以及聚类分析等方法,探究赤皮青冈的表型变异规律及其与地理气候因子的相关性。结果表明:(1)赤皮青冈表型多样性水平较高,15个表型性状的变异系数和Shannon-Wiener指数平均值分别为35.070%和1.998,14个种群的变异系数在14.94%(洞口)~35.56%(龙山)之间、Shannon-Wiener指数在1.127(松阳)~1.980(常宁)之间。(2)15个表型性状在种群间和种群内均存在极显著差异(P<0.01),种群平均表型分化系数为41.491%,表型变异主要来源于种群内。(3)相关性分析发现,赤皮青冈部分性状间存在显著或极显著相关性,表型性状与地理因子间相关性不明显,降水是影响赤皮青冈表型性状的主要气候因子。(4)主成分分析发现,基于11个表型可提炼出4个主成分,累计贡献率为82.961%。(5)根据聚类分析可将14个种群分为三类,分别为小叶片小叶柄类、大叶片大叶柄类和中叶片小叶柄类,种群的聚类结果与其地理位置间无明显相关性。该研究结果为赤皮青冈种质资源的保护和利用提供了科学依据,为其良种选育奠定了重要基础。
关键词: 赤皮青冈, 珍贵树种, 种质资源, 表型性状, 变异规律
中图分类号:Q948.12
文献标识码:A
文章编号:1000-3142(2023)09-1622-14
收稿日期:2022-09-15
基金项目:国家自然科学基金 (32201589); 湖南省自然科学基金 (2021JJ41069); 湖南省林业科技创新资金项目 (XLKY202218); 福建省公益类科研院所专项(2022R1010007)。
第一作者: 秦之旷(1997-),硕士,研究方向为林木育种,(E-mail)1667830341@qq.com。
*通信作者:李何,博士,讲师,硕士研究生导师,研究方向为林木育种,(E-mail)lihecsuft@126.com。
Phenotypic diversity of Quercus gilva natural
populations in middle subtropical China
QIN Zhikuang1, LIU Na1, ZHOU Xia1, LI Zhihui1, CAO Fuliang2, LI He1*
( 1. College of Forestry, Central South University of Forestry and Technology, Changsha 410004, China;
2. College of Forestry, Nanjing Forestry University, Nanjing 210037, China )
Abstract:Quercus gilva is a precious timber tree species in middle subtropical China. In order to investigate the phenotypic diversity, phenotypic variation pattern and the major geographic and climatic factors influencing phenotypic variation of Q. gilva populations, 15 growth and leaf traits of 115 individuals from 14 natural populations were measured, and then the phenotypic diversity, variation pattern and the correlation between phenotypic variation and geographic and climatic factors were studied using nested analysis of variance, phenotypic differentiation coefficient, diversity index, correlation analysis, principal component analysis and cluster analysis. The results were as follows: (1) The average variation coefficient and the average Shannon-Wiener index of 15 phenotypic traits were 35.070% and 1.998, respectively. Variation coefficient and Shannon-Wiener index of 14 populations ranged from 14.94% (Dongkou) to 35.56% (Longshan) and from 1.127 (Songyang) to 1.980 (Changning), respectively. These results indicated that Q. gilva showed a relatively high level of phenotypic diversity. (2) Significant differences in 15 phenotypic traits were found among and within populations (P<0.01), with an average phenotypic differentiation coefficient of 41.491%. This result elucidated that the phenotypic variation within populations was greater than that among populations. (3) Correlation analysis showed that there were significant or entremely significant correlations among parts of the traits, whereas no correlation was found between phenotypic traits and geographic factors. Precipitation was the major factor that influenced the phenotypic traits of Q. gilva. (4) Principle component analysis indicated that the first four principal components explained 82.961% of total phenotypic contribution rate of Q. gilva. (5) Cluster analysis revealed that 14 Q. gilva populations were divided into three groups according to their different leaf and petiole size, such as small-leaved micropetiole, large-leaved macropetiole and medium-leaved micropetiole, which was not related to the geographic distribution of populations. This study provides scientific basis for the protection and utilization of Q. gilva germplasm resources, and lays an important foundation for its improved variety breeding.
Key words: Quercus gilva, precious tree species, germplasm resources, phenotypic traits, variation pattern
赤皮青冈(Quercus gilva)是壳斗科青冈属常绿乔木,其树干通直,边材呈黄褐色,心材呈暗红褐色,纹理优美具有光泽,强韧有弹性,有独特香味,抗虫蛀,不易腐蚀,是我国亚热带地区极具经济价值和发展前景的珍贵用材树种之一(端木,1995;Zaynab et al., 2018)。其树体高大,为深根性树种,在水土保持和涵养水源方面有良好的表现,从而具有很高的生态价值(夏蝉,2021)。赤皮青冈在我国中亚热带地区分布较广,主要分布于湖南、福建、浙江、江西、贵州、台湾等省(Zeng et al., 2019;欧阳泽怡等,2021)。然而,从20世纪开始,赤皮青冈天然群落受到明显的人为干扰,绝大部分赤皮青冈天然林被过度采伐,采伐迹地用于城乡发展、筑路修桥或人工林营造,导致大多赤皮青冈处于散生和小群落生存状态,其种质资源大量流失(Deng et al., 2018)。
种质资源既是遗传多样性的载体,也是种质创新和林木遗传改良的物质基础,阐明种质资源的多样性是对其合理保护和利用的前提(Bai et al., 2019)。植物表型是遗传因子和环境在长期过程中共同作用的结果,表型多样性是遗传多样性的直接体现(李洪果等,2019;郭燕等,2022)。利用植物的表型性状检测遗传多样性具有简便、快捷的特点(尚帅斌等,2015),是长期以来最基本和最直接的方法。近年来,种质资源的表型性状分析已在闽楠(Phoebe bournei)(Li et al., 2018;欧汉彪等,2021)、黑杨(Populus nigra)(陈存等,2021)、板栗(Castanea mollissima)(郭燕等,2022)、茶条槭(Acer ginnala)(武艳虹等,2018)、唐古特白刺(Nitraria tanguticum)(吉德娟等,2021)等林木上开展,通过分析各表型性状,揭示了表型多样性和变异,挖掘了影响表型变异的关键因子,为相关树种种质资源的保护和利用奠定了重要基础。
目前,赤皮青冈天然种群仍处于持续减少的状态,天然种质资源不断流失,调查与阐明天然种质的多样性以合理制定其保护和利用策略是当前亟待开展的重要研究。但是,关于赤皮青冈种质资源的相关研究非常有限,仅谢健(2011)和林文俊等(2017)对赤皮青冈天然群落进行了调查,并且调查群落仅分布在闽清和擎天岩村,很难具有代表性。在分子层面,虽然有基于ISSR分子标记技术(朱品红,2014)和叶绿体基因组测序技术(Sugiura et al., 2015)的赤皮青冈遗传多样性和遗传结构的相关报道,但所调查的种群分布较集中,不足以揭示我国赤皮青冈种质的遗传多样性;在表型层面,仅谢健(2021)对4个种源赤皮青冈幼林的生长性状进行了分析。树高、胸径、材积和干型等是用材树种的重要经济性状,而叶片是植物进行光合和蒸腾作用的重要媒介,其形态直接影响生长发育和繁殖特性,并最终影响树种的适应和进化(郭燕等,2022)。
本研究以具有代表性的、基本覆盖赤皮青冈在大陆地区分布范围的14个赤皮青冈天然种群115个单株为材料,对其15个树体和叶片性状进行测量,采用巢式方差分析、表型分化系数分析、多样性指数分析、相关性分析、主成分分析以及聚类分析等方法,通过分析表型多样性和变异及其地理变化,拟探讨:(1)赤皮青冈天然种群表型多样性水平;(2)赤皮青冈表型多样性呈现的变异规律;(3)赤皮青冈表型变异与地理气候因子之间的相关性。以期为赤皮青冈种质资源的收集、保护和利用提供科学依据,为其良种选育奠定重要基础。
1 材料与方法
1.1 材料
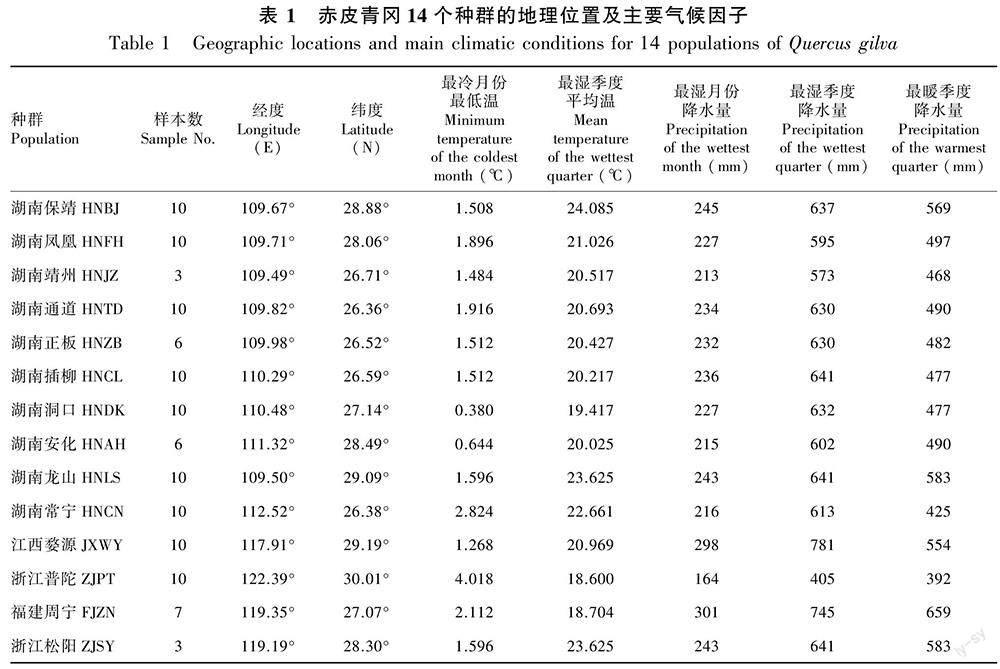
基于前期对赤皮青冈天然分布区内种质资源的调查,2021年5月至2022年3月对14个赤皮青冈天然种群共115个单株进行了调查和样本采集,采样种群基本覆盖了我国中亚热带赤皮青冈天然分布区,各种群的地理气候信息和样本数量如表1所示。对样本数大于10个的种群,随机选取10株样本,并确保样本间较大的距离;由于部分种群样本较少或采样困难,因此对样本数小于10株的种群,则采集种群内的所有样本。用GPS记录各样本的经度和纬度,利用测高仪、胸径尺等工具测量且记录树体性状。用高枝剪等工具在當地12:00—14:00时树冠中下层的阳面,获取健康枝条,随机采集成熟叶片(李珊等,2016),并选取10片无病虫害的完整叶片进行集中拍照、记录。
1.2 气候数据的获取
从全球气候数据网WorldClim(https://www.worldclim.org/)下载当前时期气候数据,共下载与温度和降水相关的19个气候因子,数据精度为2.5’,版本为2.1(Fick & Hijmans, 2017),使用R包Raster中extract函数提取种群所在位置的气候数据。在19个气候因子中,由于一些因子间可能存在高度相关性,因此用prcomp函数对种群所在地的气候因子进行主成分分析,提取前三个主成分中载荷大于0.8的气候因子,得到5个气候因子(表1),用于相关性分析。
1.3 表型性状的测定
对各种质的树高、胸径、冠径等树体性状进行测量,其中冠径为东西冠径和南北冠径的平均值,每个性状重复测量3次,取平均值作为实测值。
计算各种质的单株材积(v),公式为v=0.4π×(d1.3/2)×h+3×10(式中,d1.3为胸径,h为树高)(林昌庚,1964)。
记录树干干型情况,对这一非数值性状由3人分别进行1~10赋值统计。根据刘雨等(2011)的研究且结合赤皮青冈实际情况进行以下赋值:有明显主干无分枝,树干通直记为10;有明显主干无分枝,但树干轻微弯曲记为9;有明显主干无分枝,但树干弯曲明显记为8;有明显主干且出现1个分岔,但分岔和主干较为通直记为7;有明显主干和1个分岔,树干较为弯曲记为6;有明显主干和多个分岔,主干分枝较直记为5;有主干但第1个分岔较早,枝下高小于1.3 m,主干分枝较直记为4;有主干但第1个分岔较早,枝下高小于1.3 m,主干分枝较弯曲记为3;无明显主干,但主要枝条较直记为2;无明显主干,枝条较弯曲记为1。
使用ImageJ 图像分析软件分析各种质的叶片图像,测量获取叶片的叶长、叶宽、叶面积、叶柄长、叶柄宽、叶柄面积、锯齿数等数据,重复3次,取平均值作为实测值。通过测量值计算叶长宽比、叶柄长宽比和叶脉间距。其中,叶脉间距计算公式为叶脉间距=叶长/(单边叶脉数+1)(郭燕等,2020)。
1.4 数据的处理和分析
1.4.1 巢式方差分析和多重比较 在SPSS 26软件中,采用巢式设计模型对赤皮青冈的15个表型性状进行巢式方差分析,其线性模型为Yijk=m+ai+bj(i)+eijk。式中, Yijk为第 i个种群的第 j个单株的第 k个观测值, m为总体平均值, ai为第 i个种群的效应值,bj(i)为第 i个种群的第 j个单株的效应值, eijk为第ijk个观测值的试验误差。
1.4.2 多样性指数分析 在Excel软件中,通过公式CV=δ/x× 100%和VST=δ2t/(δ2t+δ2s)计算各性状的变异系数(CV)及表型分化系数(VST)(葛颂等,1988)。式中,δ2t表示种群间方差,δ2s表示种群内方差。在Past 4.09中计算各种群表型性状的Shannon-Wiener多样性指数,先根据平均值(mean,x)和标准差(SD,δ)将所有性状数据分成 10级,转换为质量性状,第一级为< x-2δ,中间每级增加0.5δ,直到第10级为≥ x+2δ,再计算Shannon-Wiener多样性指数。
1.4.3 相关性分析、主成分分析与聚类分析 使用SPSS 26软件,通过Pearson相关系数分析法,研究赤皮青冈各表型性状间以及性状与地理位置和气候因子间的相关性。在R软件中,先通过prcomp函数表型数据进行标准化,再进行主成分分析。基于主成分得分系数矩阵,提取贡献率最大的第一和第二主成分,采用平方欧式距离和组间连接法进行种群间聚类分析。赤皮青冈天然种群受破坏较严重,大多种群内样本数量较少,并且种群间树龄结构差异较大,靖州和松阳种群仅由3株古树和少量次生幼树组成,保靖、通道、正板、插柳、安化、龙山和周宁等种群内仅存几株古树和中龄赤皮青冈,凤凰、洞口、常宁、婺源和普陀种群为树龄较大的同龄林。因此,在进行主成分分析、聚类分析以及表型性状与地理和气候因子相关性分析时,剔除树高、胸径、冠径和材积4个受树龄影响较大的表型性状,使用其余11个表型性状进行分析。
2 结果与分析
2.1 赤皮青冈表型性状变异
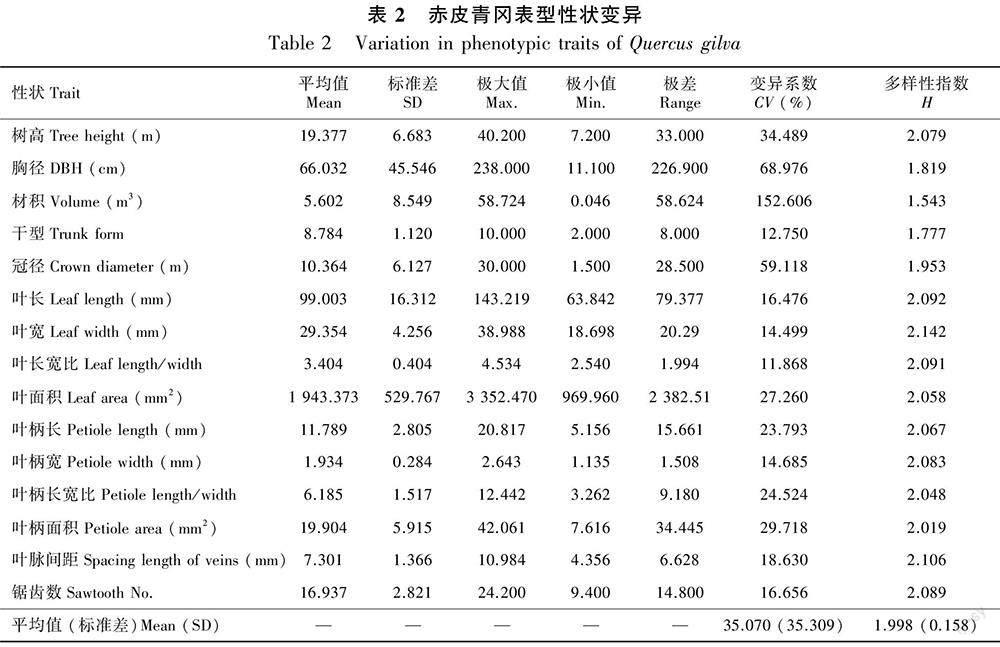
赤皮青冈15个表型性状的变异系数平均值为35.070%(表2)。其中,材积的变异系数最大(为152.606%),变异幅度为0.05~58.72 m3。叶片长宽比变异幅度最小(为11.868%),变异幅度为2.540~4.534。15个表型性状的变异系数由大到小依次为材积>胸径>冠径>树高>叶柄面积>叶面积>叶柄长宽比>叶柄长>叶脉间距>锯齿数>叶长>叶柄宽>叶宽>干型>叶长宽比。这说明赤皮青冈树体性状变异更加丰富。
15个表型性状的多样性指数(H)在1.543~2.142之间,平均值为1.998(表2)。其中,材积的多样性指数最低(为1.543),叶宽的多样性指数最高(为2.142)。15个表型性状的多样性由高到低分别为叶宽>叶脉间距>叶长>叶长宽比>锯齿数>叶柄宽>树高>叶柄长>叶面积>叶柄长宽比>叶柄面积>冠径>胸径>干型>材积。这说明赤皮青冈在个体水平上存在较为丰富的表型变异。
2.2 赤皮青冈表型变异来源及种群变异规律
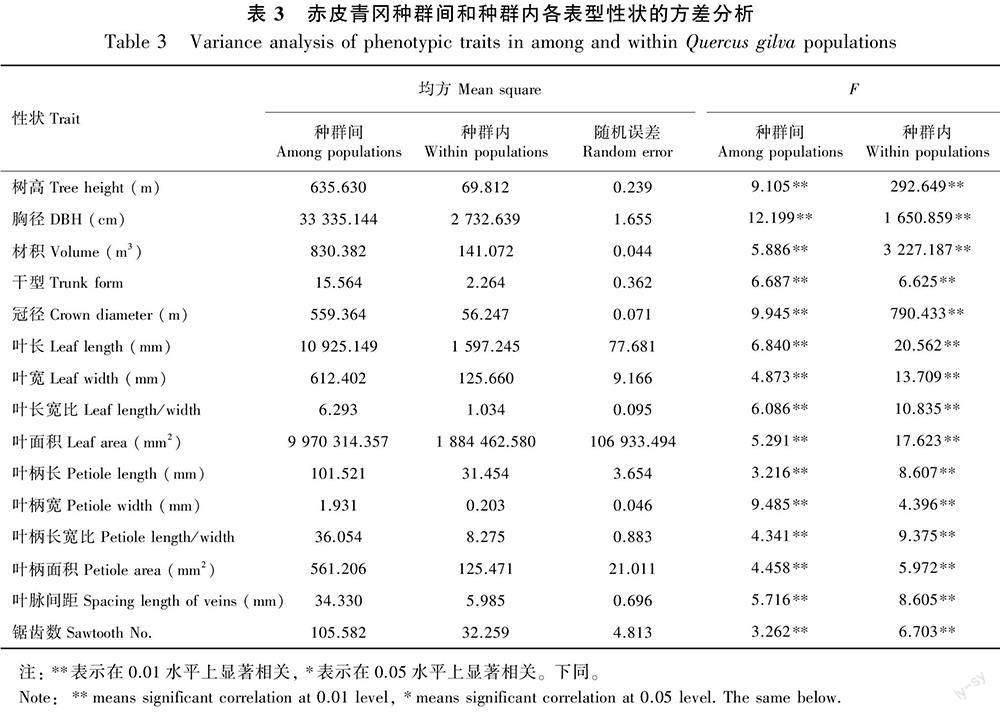
赤皮青冈15个表型性状在种群间和种群内的方差分析结果见表3。由表3的F检验结果可知,无论是在种群间还是种群内15个表型性状都存在极显著差异(P<0.01),说明赤皮青冈的树体和叶片性状在种群间和种群内的变异都较为丰富。
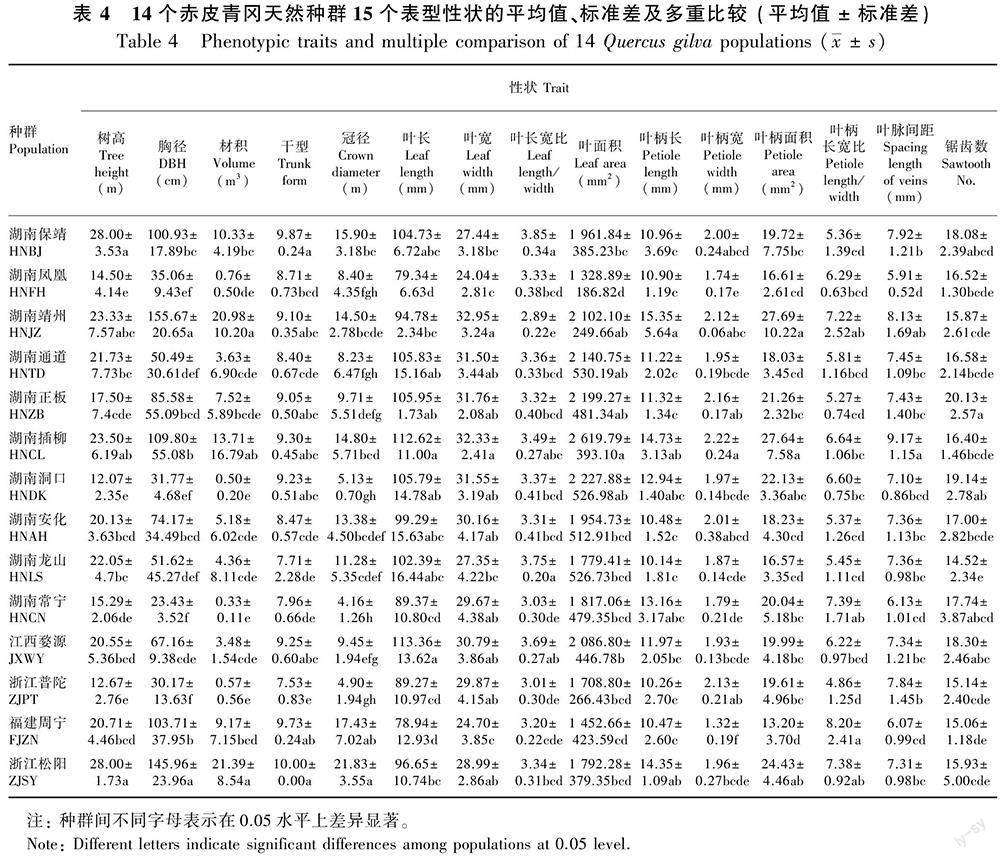
由表4可知,保靖、插柳、安化、龙山、婺源、周宁、松阳种群的树高、胸径、冠径和干型等树体指标值都较大,其中松阳种群的树高、材积、冠径、干型均为最大,表现为树形高大通直的特征;靖州種群的胸径最大;凤凰、洞口、常宁、普陀种群的平均材积相对较小,与各种群实际树龄结构相符。靖州、常宁、普陀种群的叶长宽比较小,叶片表现为短而宽的叶形;保靖、龙山、婺源种群的叶长宽比较大,叶片呈细长型;插柳种群的叶长和叶面积最大;正板赤皮青冈的叶片锯齿数最多,叶面积较大,表现为叶片大且锯齿多的特点。
2.3 赤皮青冈种群内表型变异特征
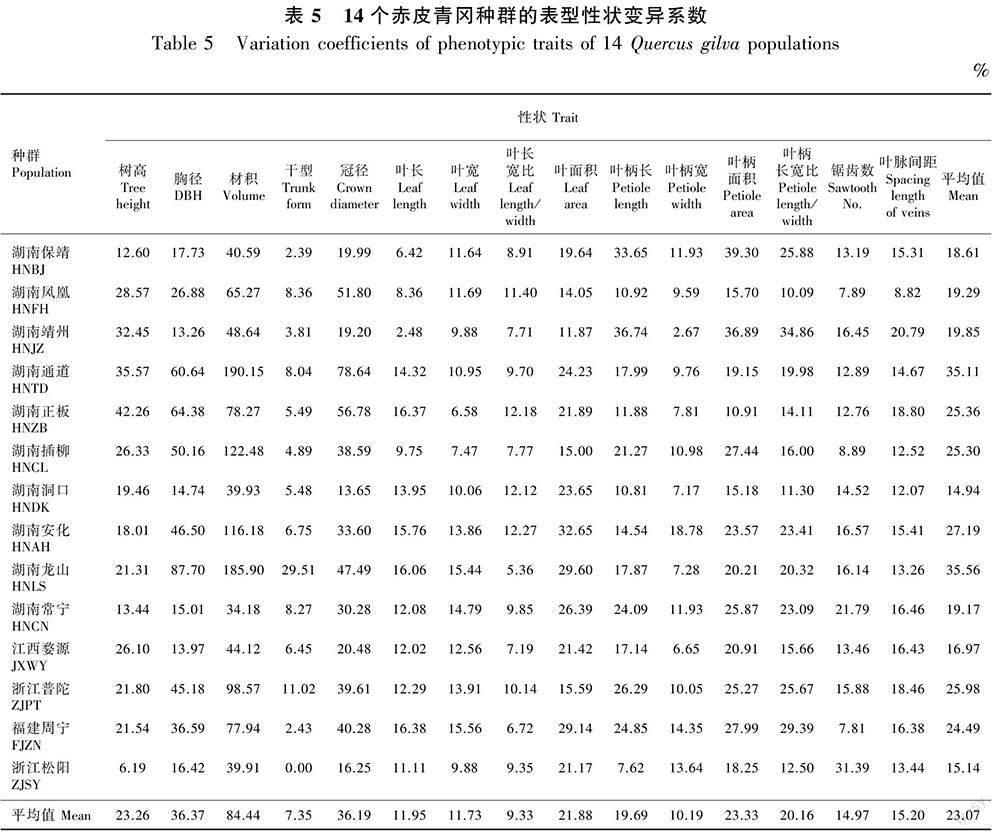
变异系数(CV)表示种群内表型性状的变异情况,变异系数越大表明表型的离散程度越大。由表5可知,14个种群的变异系数在14.94%(洞口)~35.56%(龙山)之间,平均值为23.07%,说明在种群水平上赤皮青冈的表型性状变异水平较高;14个种群中,仅洞口种群的15个表型性状变异系数平均值未超过15%,说明该种群赤皮青冈表型变异水平较低;15个表型性状中干型、叶长、叶宽、叶长宽比、叶柄宽和锯齿数的变异系数平均值都未超过15%,说明这6个性状较为稳定。与表2结果一致,除干型外,树高、胸径、冠径和材积等树体性状的变异系数平均值大于叶片性状的变异系数平均值,说明赤皮青冈天然种质的树体性状受树龄影响过高而表现出较高变异水平。
2.4 赤皮青冈种群内表型多样性
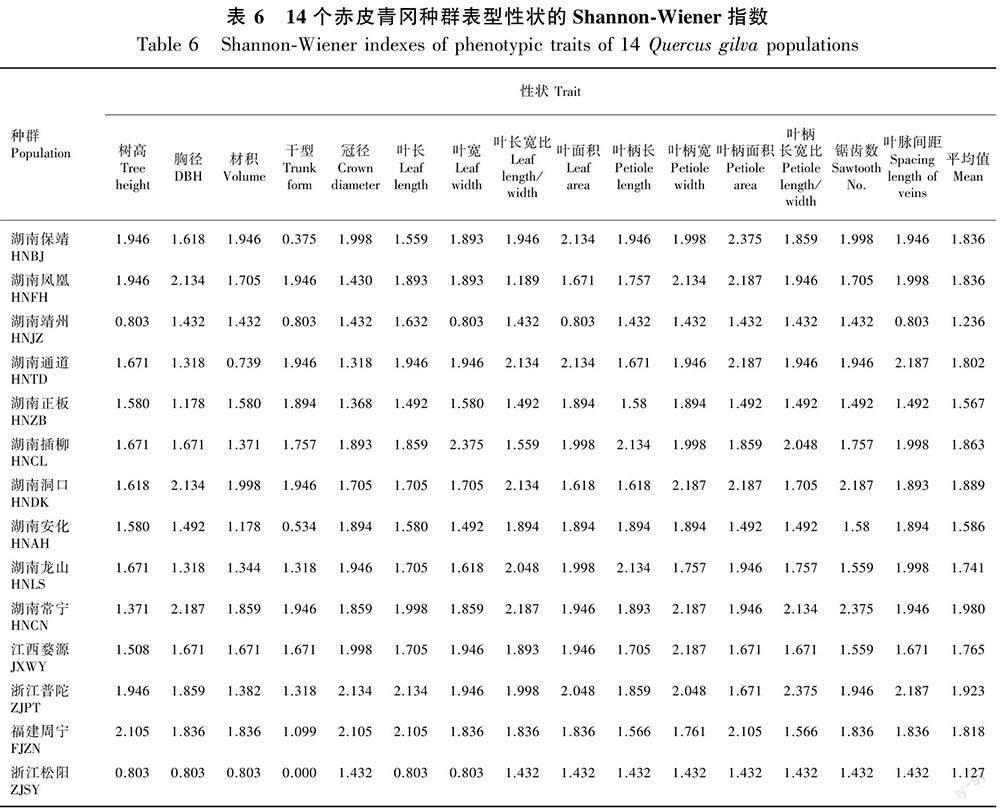
Shannon-Wiener多样性指数反映不同表型性状的丰富和均匀程度,14个种群的15个表型性状的多样性指数见表6。由表6可知,Shannon-Wiener指数在1.127(松阳)~1.980(常宁)之间,14个种群的多样性指数从高到低依次为常宁>普陀>洞口>插柳>保靖>凤凰>周宁>通道>婺源>龙山>安化>正板>靖州>松阳。Shannon-Wiener多样性指数与种群内样本数量大小有一定的关系,靖州、安化、松阳等样本较少的种群表现出较低的Shannon-Wiener指数,在种质资源收集和保护工作中应重视。
2.5 赤皮青冈表型性状变异来源及种群间表型分化
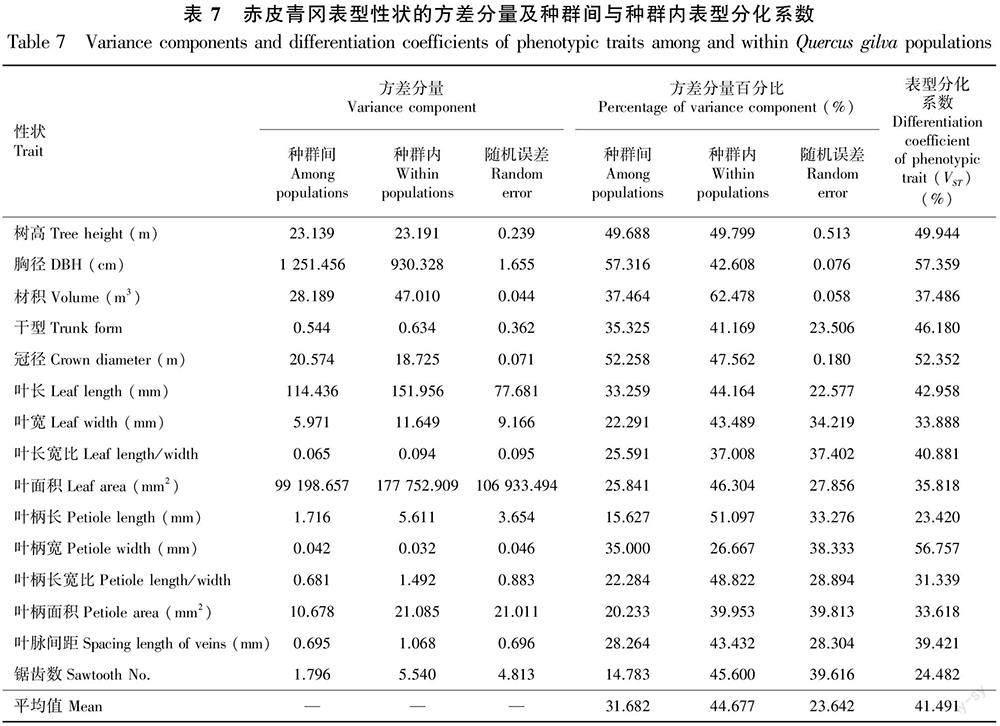
表型分化系数(VST)是种群间表型变异占总表型的百分比,用于评估种群间的表型分化情况。由表7可知,15个赤皮青冈表型性状的分化系数在23.420%(叶柄长)~57.359%(胸径)之间。赤皮青冈种群间表型方差分量占总表型变异的31.682%,种群内占44.677%,表型分化系数为41.491%,说明种群内的变异在赤皮青冈表型变异中起主要作用。
2.6 赤皮青冈表型性状间的相关性及其与地理和气候因子间的相关性
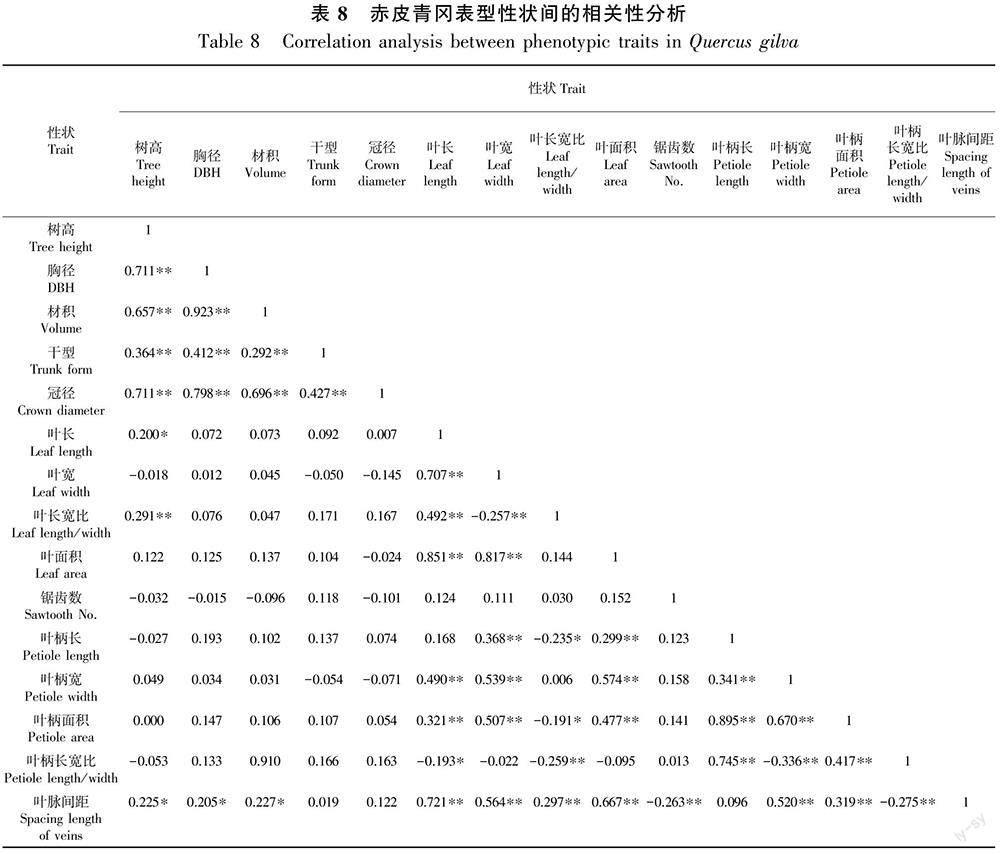
15个赤皮青冈表型性状间的相关性结果如表8所示。其中,部分性状间存在显著或极显著相关性。树高与胸径、材积、干型、冠径、叶长宽比呈极显著正相关性,与叶长和叶脉间距呈显著正相关,说明较高的赤皮青冈往往具有较大的胸径、较优良的干型和冠形,叶片也较为狭长。干型与胸径、材积、冠径均呈极显著正相关,说明优良的干型在一定程度上可以提高赤皮青冈的生长潜力。叶面积与叶长宽比无明显相关性,说明不同种质的叶长宽比表现出一定程度的稳定性。叶宽与叶柄长、叶柄宽、叶柄面积均呈极显著正相关,叶长和叶面积与叶柄宽和叶柄面积呈极显著正相关,说明大的叶片通常伴随叶柄粗长的特征。叶脉间距仅与锯齿数呈极显著负相关,说明锯齿越多叶脉间距越小。
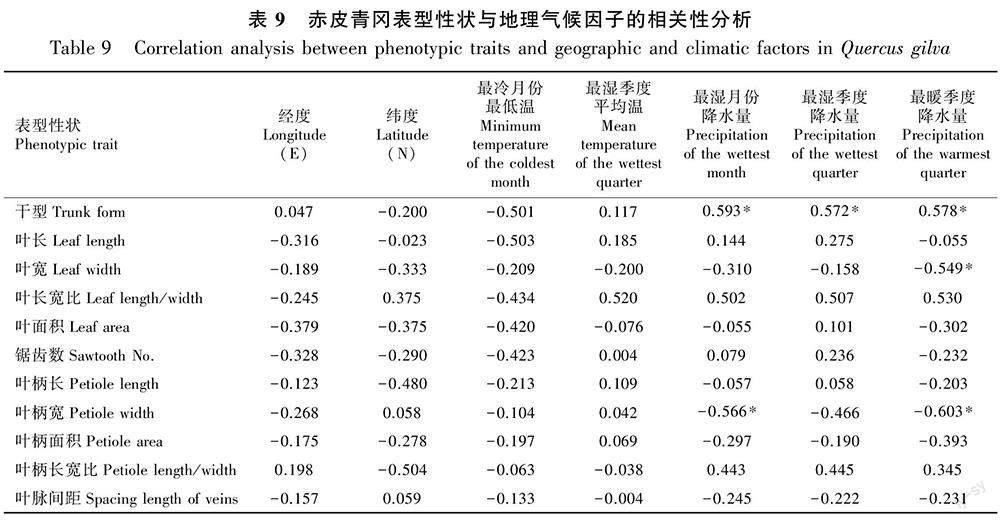
11个表型性状与种群所在地的地理和气候因子的相关性分析结果如表9所示。赤皮青冈表型性状与地理气候因子间的相关性不明显。干型与最湿月份降水量、最湿季度降水量和最暖季度降水量呈显著正相关,叶宽与最暖季度降水量呈显著负相关,叶柄宽与最湿月份降水量和最暖季度降水量呈显著负相关;经度、纬度、最冷月份最低温度和最湿季度平均温度与赤皮青冈表型性状间无相关性。这表明降水对赤皮青冈表型性状的影响大于温度和地理因子。
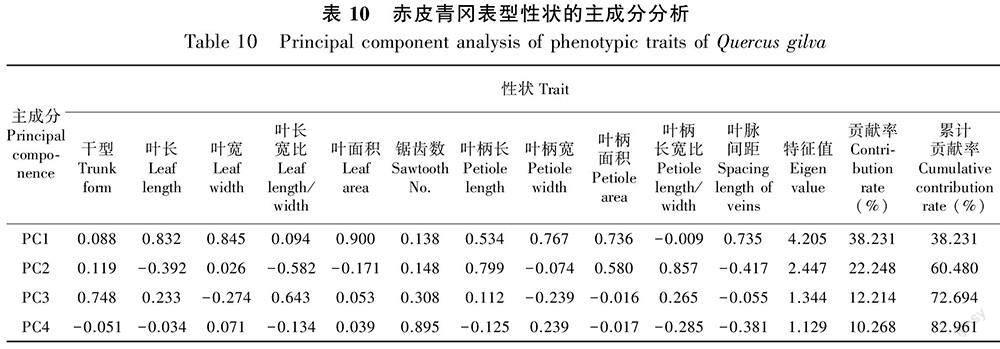
2.7 赤皮青冈表型性状的主成分分析和聚类分析
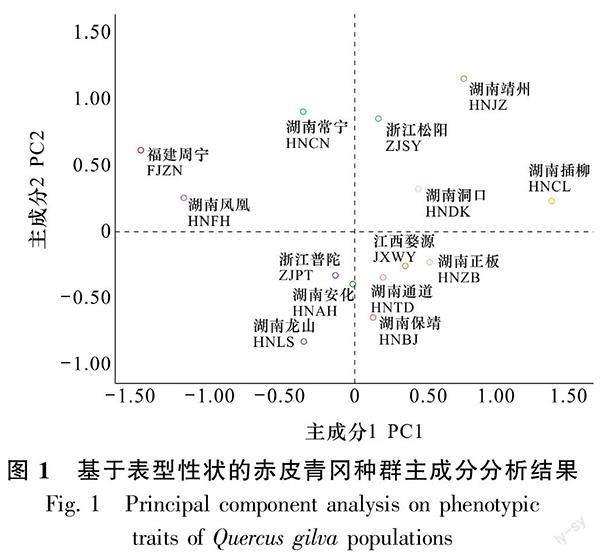
对赤皮青冈11个表型性状进行主成分分析,得到4个特征根大于1的主成分,累计贡献率为82.961% (表10)。由表10可知, PC1特征值为4.205,解释总变异的38.231%, 主要包括叶长、 叶宽、叶面积(载荷的绝對值>0.8),主要表征叶片大小;PC2的特征值为2.447,解释了22.248%的总变异,主要包括叶柄长和叶柄长宽比,与叶柄性状相关;PC3特征值为1.344,主要表征干型,解释了12.214%的总变异;PC4特征值为1.129,主要表征锯齿数,解释了10.268%的总变异。由图1可知,赤皮青冈表型性状未呈现出明显的地理变异规律,其中湖南凤凰和福建周宁种群以其较低的PC1得分与其他种群区分开来。
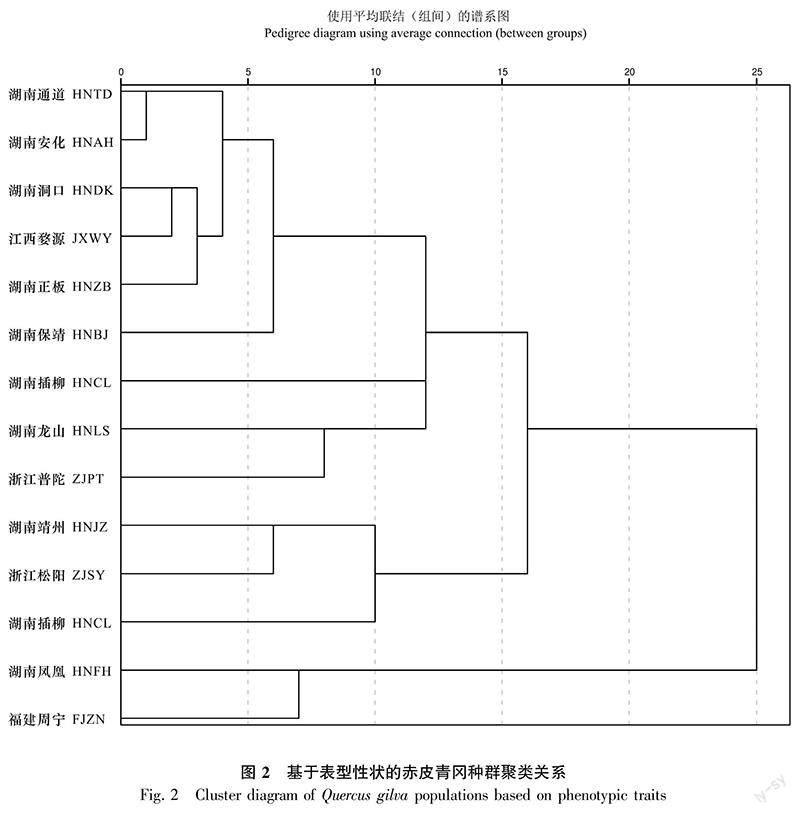
以平方欧式距离为14个赤皮青冈种群间的距离,采用组间连接法进行聚类(图2)。当平方欧式距离为15时,根据11个表型性状可将14个种群分为三类(与主成分分析结果相符):第一类为湖南凤凰和福建周宁,主要特征为叶长、叶宽、叶面积、叶柄长、叶柄宽、叶柄面积和叶脉间距值均最小(表4),表现为小叶片小叶柄类;第二类为湖南靖州、浙江松阳和湖南插柳,其叶宽、叶面积、叶柄长、叶柄宽、叶柄面积和叶脉间距的值最大,叶长宽比值最低,说明第二类呈现出叶片大、叶宽且叶柄粗长的特征,表现为大叶片大叶柄类;第三类包括湖南保靖、湖南通道、湖南正板、湖南洞口、湖南安化、湖南龙山、湖南常宁、江西婺源和浙江普陀9个种群,属于第一类和第二类的衔接部分,叶片形态接近第二类,叶长、叶宽、叶面积的值普遍较大,但叶柄形态更接近第一类,普遍较小,总体表现为中叶片小叶柄类。通过比较种群的聚类结果和地理位置之间的关系,发现二者关联并不显著,说明赤皮青冈表型性状与地理因子间的相关性不明显。
3 讨论与结论
3.1 赤皮青冈表型性状的多样性
表型性状的变异是环境和遗传因子共同影响下的表达(曹德美等,2021),尤其是赤皮青冈这类珍稀植物,种质资源流失严重且面临复杂的生存环境和长期的地理隔离,极易发生树体和叶片等表型性状的变异。
本研究结果表明,赤皮青冈15个表型性状的Shannon-Wiener多样性指数在1.543~2.142之间,平均值为1.998。高于濒危植物云南黄连(1.634~1.839)(Coptis chinensis)(杨维泽等,2013),与濒危植物格木(Erythrophleum fordii)(2.027 8)(李洪果等,2019)相近,低于四川珙桐(2.360)(Davidia involucrata)(张腾驹等,2019)。这表明赤皮青冈虽面临种质资源流失的问题,仍表现出较高的多样性水平,特别是叶长、叶宽和叶脉间距等性状。叶片的形态直接影响叶片生长发育特性,间接影响植物的适应能力,赤皮青冈叶片表型性状的多样性说明其具有较强的环境适应性。
本研究中,赤皮青冈15个表型性状的极大值是极小值的1.78(叶长宽比)~1 174.40(材积)倍,其中树体性状的极大值与极小值的比值普遍大于叶片性状,表明树体性状较叶片性状更易受到树龄本身的影响。本研究中,赤皮青冈15个表型性状的变异系数在11.868%~152.606%之间,天然种质间表型性状差异较大,尤其是树高、胸径、冠径和材积等受树龄影响大的树体性状;剔除以上4个性状后,其余11个表型性状的变异系数为11.868%~29.718%,与灰楸(Catalpa fargesii)(9.66%~30.39%) (于晓池等,2021) 和板栗(Castanea mollissima)(8.68%~32.73%)(郭燕等,2022)相近。这说明赤皮青冈干型和叶片性状变异丰富,在不同环境因子和遗传因子的影响下做出相应的改变,这与Shannon-Wiener多样性指数的结论一致,表明赤皮青冈在复杂生长环境中表现出较强的适应能力。
3.2 赤皮青冈表型性状的变异来源和分化
本研究中,赤皮青冈15个表型性状在种群间和种群内都表现出较丰富的变异,而种群内的表型变异更为丰富(平均表型分化系数VST=41.491%)。这与大多数林木种质资源的表型研究结果基本一致,如小果白刺(Nitraria sibirica)(40.71%)(董昕等,2017)、香椿(Toona sinensis)(40.209%)(吴军等,2018)等树种均存在较为丰富的表型变异。这表明赤皮青冈叶片等表型性状在种群间的遗传分化程度相对较低,可能是本研究所调查种群均位于我国亚热带地区,分布区内土壤理化性质和气候环境等条件差别不大,并且赤皮青冈在自然分布范围内呈碎片化分布的时间较短,暂未因生殖隔离而形成较大的差异。
本研究中,赤皮青冈11个叶片及干型性状无论是在种群间还是在种群内都存在极显著差异,而种群内的表型变异为主要变异来源(表型分化系数在23.420%~56.757%之间)。同时,14个种群的11个叶片及干型性状的变异系数平均值在7.35%(干型)~23.33%(叶柄面积)之间,并且同一表型性状的Shannon-Wiener指数在种群间存在较大差异,提示该物种较易发生种群分化。本研究所调查的赤皮青冈天然种群大多零散分布于村落周围,虽然个别天然种群保存较为完整,但大部分种群因受人为干扰较严重而造成天然生境的改变,加上生境不连续、样本较少的零散分布,种群间的基因流受阻,从而引起种群分化和表型性状变异。
3.3 赤皮青冈表型变异与地理气候因子间的相关性
不同物种对环境因子的敏感度和适应力有一定的差异,其表型性状呈现出不同的地理变异规律,如板栗(郭燕等,2022)的叶片由南向北表现出长宽比变小的趋势,唐古特白刺(吉德娟等,2021)的叶片厚度与海拔呈正相关,青杨(Populus cathayana)(曹德美等,2021)的侧脉数与经纬度呈极显著正相关。本研究中,在适宜赤皮青冈生长的环境下,经纬度和温度对赤皮青冈表型性状的影响不显著,而降水与赤皮青冈表型性状间存在一定的相关性。这与姜小龙(2020)的研究结果类似,降水是影响南岭青冈(Quercus championii)和福建青冈(Q. chungii)分布最大的气候因子。本研究发现,气候因子中最湿月份降水量、最湿季度降水量以及最暖季度降水量仅与赤皮青冈干型呈正相关,而最湿月份降水量和最暖季度降水量与叶宽和叶柄宽呈负相关。在适宜的降水范围内,降水量的增加会促进维管形成层的活动,有助于树干的均匀增粗,从而表现出优良的干型。随着降水量的增加,栎类树种的叶片和叶柄倾向于长条状发展(李东胜等,2013),叶宽和叶柄宽表现出减小的趋势,该现象与苦楝(Melia azedarach)(陈丽君等,2016)、板栗(郭燕等,2022)等的研究结果相似。因此,降水是影响赤皮青冈表型和分布最主要的因子,这为赤皮青冈的资源圃和人工林基地建设提供了科学依据。
3.4 赤皮青冈的保护与开发利用策略
赤皮青冈天然种群的生境不连续、跨度较大、碎片化较为严重,目前大多天然种群处于被孤立状态且包含的种质数量较少。赤皮青冈种子为顽拗性种子,外形较大,自然扩散距离十分有限,并且花粉的传播受到我国南方复杂地形的阻碍,长此以往,高片段化分布的种群间基因流受阻(姜小龙,2020),加上环境异质性的影响,从而引起种群分化和表型性状变异。因此,仅开展就地保护既不能有效地为赤皮青冈种群间的基因交流提供廊道,也不利于种群的延续和遗传多样性的维持。为了更好地保护、管理和利用赤皮青冈种质资源,应加强对现有天然种群和单株的保护力度,特别是单株数量稀少的种群;加快开展种质资源收集工作,鉴于赤皮青冈种群内表型变异水平较高,要注重對种群内每个单株的收集;应将赤皮青冈作为乡土树种在其天然分布区内推广造林,以适应能力较强的大叶片大叶柄类的靖州、松阳和插柳种群为种源进行采种育苗和造林,既可提高赤皮青冈的适应性,又可改善种群片段化分布的现状,为种群间的交流提供廊道,帮助该物种的延续。
参考文献:
BAI QS, CAI YL, HE BX, et al., 2019. Core set construction and association analysis of Pinus massoniana from Guangdong Province in southern China using SLAF-seq[J]. Sci Rep, 9: 13157.
CAO DM, ZHANG YH, CHENG XQ, et al., 2021. Genetic variation of leaf phenotypic traits in different populations of Populus cathayana [J]. Sci Silv Sin, 57(8): 56-67.[曹德美, 张亚红, 成星奇, 等, 2021. 青杨不同种群叶片表型性状的遗传变异 [J]. 林业科学, 57(8): 56-67.]
CHEN C, DING CJ, HUANG QJ, et al., 2021. Construction of phenotypic core collection of Populus deltoides [J]. For Res, 34(2): 1-11.[陈存, 丁昌俊, 黄秦军, 等, 2021. 美洲黑杨表型核心种质库构建 [J]. 林业科学研究, 34(2): 1-11.]
CHEN LJ, LIU MQ, LIAO BY, et al., 2016. Variation of leaf morphological characters of Melia azedarach from different provenances [J]. J SW For Univ, 36(5): 16-20.[陈丽君, 刘明骞, 廖柏勇, 等, 2016. 苦楝不同种源叶片性状变异研究 [J]. 西南林业大学学报, 36(5): 16-20.]
DENG M, JIANG XL, HIPP AL, et al., 2018. Phylogeny and biogeography of East Asian evergreen oaks (Quercus section Quercus; Fagaceae): Insights into the Cenozoic history of evergreen broad-leaved forests in subtropical Asia [J]. Mol Phylogenet Evol, 119: 170-181.
DONG X, WANG L, LU YZ, et al., 2017. Phenotypic variation of Nitraria sibirica natural populations in Shandong [J]. For Res, 30(2): 293-299.[董昕, 王磊, 鲁仪增, 等, 2017. 山东稀有植物小果白刺天然群体表型变异研究 [J]. 林业科学研究, 30(2): 293-299.]
DUAN M, 1995. Comprehensive utilization of Quercus resources in China [J]. J Beijing For Univ, 17(2): 109-110.[端木, 1995. 我国青冈属资源的综合利用 [J]. 北京林业大学学报, 17(2): 109-110.]
FICK SE, HIJMANS RJ, 2017. WorldClim 2: new 1km spatial resolution climate surfaces for global land areas [J]. Int J Clim, 37(12): 4302-4315.
GE S, WANG MX, CHEN YW, 1988. An analysis of population genetic structure of Masson pine by isozyme tech-nique [J]. Sci Silv Sin, 24(4): 399-409. [葛颂, 王明庥, 陈岳武, 1988. 用同工酶研究马尾松群体的遗传结构 [J]. 林业科学, 24(4): 399-409.]
GUO Y, ZHANG SH, LI Y, et al., 2022. Diversity analysis of 36 leaf phenotypic traits of Chinese chestnut [J]. Sci Agric Sin, 55(5): 991-1009.[郭燕, 张树航, 李颖, 等, 2022. 中国板栗36个叶片表型性状的多样性 [J]. 中国农业科学, 55(5): 991-1009.]
GUO Y, ZHANG SH, LI Y, et al., 2020. Studies on the leaf morphology, anatomical structure and drought resistance evaluation of 238 Chinese chestnut varieties (Strains) [J]. Acta Hortic Sin, 47(6): 1033-1046.[郭燕, 张树航, 李颖, 等, 2020. 中国板栗238份品种(系)叶片形态、解剖结构及其抗旱性评价 [J]. 园艺学报, 47(6): 1033-1046.]
JIANG XL, 2020. Phylogenetic relationship and population genetic structure of Quercus chungii and Q. championii[D]. Changsha: Central South University of Forestry and Technology.[姜小龍, 2020. 福建青冈和岭南青冈系统发育关系及居群遗传结构 [D]. 长沙: 中南林业科技大学.]
JI DJ, ZHANG DF, YU Q, 2021. Phenotypic diversity of Nitraria tangutorum Bobr. in Qaidam basin [J]. J Centr S Univ For Technol, 41(10): 57-66.[吉德娟, 张得芳, 于倩, 2021. 柴达木盆地唐古特白刺的表型多样性 [J]. 中南林业科技大学学报, 41(10): 57-66.]
LIN CG, 1964. Stem control in forest volume measurement technology [J]. Sci Silv Sin, (4): 87-97.[林昌庚, 1964. 林木蓄积量测算技术中的干形控制问题 [J]. 林业科学, (4): 87-97.]
LI YG, LIU XH, MA JW, et al., 2018. Phenotypic variation in Phoebe bournei populations preserved in the primary distribution area [J]. J For Res, 29(1): 35-44.
LI DS, SHI ZM, FENG QH, et al., 2013.Response of leaf morphometric traits of Quercus species to climate in the temperate zone of the North-South Transect of Eastern China [J]. Acta Phytoecol Sin, 37(9): 793-802.[李东胜, 史作民, 冯秋红, 等, 2013. 中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应 [J]. 植物生态学报, 37(9): 793-802.]
LI S, GAN XH, HAN HY, et al., 2016. Leaf phenotypic traits of Tetracentron sinense, an endangered plant species [J]. For Res, 29(5): 687-697.[李珊, 甘小洪, 憨宏艳, 等, 2016. 濒危植物水青树叶的表型性状变异 [J]. 林业科学研究, 29(5): 687-697.]
LIU Y, LI DW, QIN YS, et al., 2011. Phenotypic diversity for natural populations of Juniperus rigida in the Helan Mountains of Ningxia [J]. J Zhejiang A & F Univ, 28(4): 619-627. [刘雨, 李登武, 秦廷松, 等, 2011. 宁夏贺兰山杜松天然群体的表型多样性 [J]. 浙江农林大学学报, 28(4): 619-627.]
LIN WJ, LIU QY, LUO X, et al., 2017. Research on species diversity and niche of natural small population of Quercus glauca [J]. Fujian For, (6): 38-41. [林文俊, 劉沁月, 罗萧, 等, 赤皮青冈天然小居群物种多样性与生态位研究 [J]. 福建林业, (6): 38-41.]
OU HB, DONG LJ, LI J, et al., 2021. Relationship between phenotypic diversity of Phoebe bournei deed traits and environmental geographical factors [J]. J NE For Univ, 49(8): 45-50.[欧汉彪, 董利军, 李娟, 等, 2021. 闽楠种实性状表型多样性与环境地理因子的关系 [J]. 东北林业大学学报, 49(8): 45-50.]
OUYANG ZY, OUYANG SL, WU JY, et al., 2021. Research progress of precious commercial tree species Quercus gilva [J]. Hunan For Sci Technol, 48(6): 74-79.[欧阳泽怡, 欧阳硕龙, 吴际友, 等, 2021. 珍贵用材树种赤皮青冈研究进展 [J]. 湖南林业科技, 48(6): 74-79.]
SHANG SB, 2015. Genetic diversity in natural populations of Vatica mangachapoi on Hainan Island, China [D]. Beijing: Chinese Academy of Forestry.[尚帅斌, 2015. 海南岛青梅天然居群遗传多样性 [D]. 北京: 中国林业科学研究院.]
SUGIURA N, TANG DQ, KUROKOCHI H, et al., 2015. Genetic structure of Quercus gilva Blume in Japan as revealed by chloroplast DNA sequences [J]. Botany, 93(12): 873-880.
WU J, ZHONG ZZ, LOU J, et al., 2018. A study on phenotypic diversity of cone and seed in natural populations of Toona sinensis [J]. Acta Agric Univ Jiangxi, 40(2): 248-256.[吴军, 钟志真, 楼军, 等, 2018. 香椿天然群体种实性状表型多样性研究 [J]. 江西农业大学学报, 40(2):248-256.]
WU YH, FAN ZL, LI J, et al., 2018. Phenotypic diversity of seeds and fruits in natural populations of Acer ginnala in China [J]. Guihaia, 38(6): 795-803.[武艳虹, 樊泽璐, 李佳, 等, 2018. 茶条槭自然种群种子和果实表型多样性研究 [J]. 广西植物, 38(6): 795-803.]
XIA C, 2021. Effects of shading on the growth and physiological characteristics of Quercus gilva seedlings [D]. Changsha: Central South University of Forestry and Technology.[夏婵, 2021. 遮阴对赤皮青冈幼苗生长及生理特性的影响 [D]. 长沙: 中南林业科技大学.]
XIE J, 2021. Variations in growth traits and provenance selection of Quercus gilv [J]. J Fujian Agric For Univ (Nat Sci Ed), 50(5): 636-640.[谢健, 2021. 不同种源赤皮青冈幼林生长性状变异及种源选择 [J]. 福建农林大学学报(自然科学版), 50(5): 636-640.]
XIE J, 2011. Research on population ecology of Quercus gilvs [D]. Fuzhou: Fujian Agriculture and Forestry University.[谢健, 2011. 赤皮青冈种群生态学研究 [D]. 福州: 福建农林大学.]
YANG WZ, JIN H, LI WY, et al., 2013. Phenotypic diversity of different populations of endangered plant Coptis yunnanensis [J]. J Yunnan Univ (Nat Sci Ed), 35(5): 719-726.[杨维泽, 金航, 李晚谊, 等, 2013. 濒危植物云南黄连不同居群表型多样性研究 [J]. 云南大学学报(自然科学版), 35(5): 719-726.]
YU XC, LI F, OU Y, et al., 2021. Construction of core germplasm of Sorbus mandshurica based on phenotype [J]. For Res, 34(6): 38-45.[于晓池, 李凤, 欧阳, 等, 2021. 基于表型的灰楸核心种质构建 [J]. 林业科学研究, 34(6): 38-45.]
ZAYNAB M, PAN D, NOMAN A, et al., 2018. Transcriptome approach to address low seed germination in Quercus gilva to save forest ecology [J]. Biochem Syst Ecol, 81: 62-69.
ZENG QM, LIU B, LIU RQ, et al., 2019. The complete chloroplast genome sequence of Quercus gilva (Fagaceae)[J]. Mitchondrial DNA B, 4(2): 2493-2494.
ZHANG TJ, CHEN XH, KANG XK, et al., 2019. Phenotypic diversity of leaf morphologic traits of Davidia involucrata natural populations in Sichuan Province [J]. Sin J Acta Ecol, 38(1): 35-43.[张腾驹, 陈小红, 康喜坤, 等, 2019. 四川省珙桐天然种群叶表型多样性 [J]. 生态学杂志, 38(1): 35-43.]
ZHU PH, 2014. Analysis on genetic diversity and genetic structure of Quercus gilva population [D]. Changsha: Central South University of Forestry and Technology.[朱品紅, 2014. 赤皮青冈居群遗传多样性与遗传结构分析 [D]. 长沙: 中南林业科技大学.]
(责任编辑 蒋巧媛 王登惠)
