广西金钟山细叶云南松径向生长及其对气候因子的响应
2023-11-01余碧云唐金利韦海勇胡宝清苏宏新
余碧云 唐金利 韦海勇 胡宝清 苏宏新




摘 要:为探究广西金钟山细叶云南松径向生长及其对气候的响应,该文建立了细叶云南松树木年轮标准年表,采用响应分析探讨了径向生长与气候因子的关系,并使用逐步回归和方差分解量化了不同气候因子对径向生长的影响。结果表明:(1)响应分析结果显示,上一年1月、8月、9月、11月的日照时数以及上一年10月的平均最低气温和平均气温与标准年表年轮宽度均呈显著正相关,而上一年6月降雨量>10 mm的天数与年轮宽度呈显著负相关;当年2月的降雨量、3月的空气相对湿度和平均最低气温、6月的日照时数以及9月的平均气温和平均最高气温均与年轮宽度呈显著正相关。(2)逐步回归最终模型的方差分解结果显示,上一年10月的平均最低气温对细叶云南松径向生长的影响最大(方差解释量达23.35%),其次是当年9月的平均最高气温(方差解释量为10.39%);上一年1月和11月的日照时数分别解释了径向生长变异的3.94%和6.58%。综上表明,秋季的温度和冬季的光照条件是限制细叶云南松径向生长最主要的气候因子,早春干旱和雨季大降雨量会降低细叶云南松的径向生长。
关键词: 径向生长, 气候响应, 相对贡献率, 细叶云南松, 广西金钟山
中圖分类号:Q948
文献标识码:A
文章编号:1000-3142(2023)09-1568-10
收稿日期:2022-11-14
基金项目:广西自然科学基金(2018GXNSFAA281277); 南宁师范大学科研启动经费项目(20220513); 广西地表过程与智能模拟重点实验室系统基金(GTEU-KLOP-X1802)。
第一作者: 余碧云(1989-),博士,副研究员,研究方向为森林生态学,(E-mail)biyunyu1989@126.com。
*通信作者:苏宏新,博士,副研究员,研究方向为生态学,(E-mail)hxsu@nnnu.edu.cn。
Radial growth of Pinus yunnanensis var. tenuifolia and its
response to climatic factors in Jinzhongshan, Guangxi
YU Biyun1, TANG Jinli1, WEI Haiyong2, HU Baoqing1, SU Hongxin1*
( 1. Key Laboratory of Environment Change and Resources Use in Beibu Gulf (Ministry of Education)/Guangxi Key Laboratory of
Earth Surface Processes and Intelligent Simulation, Nanning Normal University, Nanning 530001, China; 2. Guangxi
Jinzhongshan Syrmaticus Humiae National Nature Reserve Management Center, Baise 533000, Guangxi, China )
Abstract:In order to explore the radial growth of Pinus yunnanensis var. tenuifolia in Jinzhongshan of Guangxi and its response to climate, a standard tree-ring chronology of P. yunnanensis var. tenuifolia was established, response analysis was used to explore the relationship between radial growth and climatic factors, and stepwise regression and variance decomposition were used to quantify the effects of climatic factors on radial growth of P. yunnanensis var. tenuifolia. The results were as follows: (1) Sunshine duration in January, August, September and November, and the mean minimum temperature and the mean temperature in October of the previous year were significantly positively correlated with tree-ring width of standard chronology. By contrast, there was a significant negative correlation between the number of days with the amount of precipitation >10 mm in June of the previous year and tree-ring width. Precipitation in February, relative air humidity and the mean minimum temperature in March, sunshine duration in June, and the mean maximum temperature and mean temperature in September of the current year were positively correlated with tree-ring width. (2) The variance decomposition results of the final stepwise regression model revealed thatthe mean minimum temperature in October of the previous year had the greatest effect on the radial growth of P. yunnanensis var. tenuifolia (explaining 23.35% of the variance), followed by the mean maximum temperature in September of the current year (explaining 10.39% of the variance). Sunshine duration in January and November of the previous year explained 3.94% and 6.58% of the variation in radial growth of P. yunnanensis var. tenuifolia, respectively. In conclusion, autumn temperature and winter light condition are the main climatic factors limiting the radial growth of P. yunnanensis var. tenuifolia in Jinzhongshan. In addition, both drought in early spring and heavy rainfall in rainy season can reduce the radial growth of P. yunnanensis var. tenuifolia.
Key words: radial growth, climate response, relative contribution rate, Pinus yunnanensis var. tenuifolia, Jinzhongshan, Guangxi
全球变化下,森林碳汇已成为碳中和的一项有效途径(Lewis et al., 2019)。树木径向生长(即树木年轮形成)是树木乃至森林最重要的碳吸收和贮存的过程之一,其动态变化很可能会影响树木和森林的碳汇(Sass-Klaassen, 2015;Rossi et al., 2016)。树木年轮是树木在径向生长过程中,因受到气候交替变化影响而形成的轮状结构(Fritts, 1976),其宽度既代表树木每年的生长状况也反映了与温度、降水和太阳辐射等气候因子的关系,可作为研究气候變化的一个重要参数(范玮熠和王孝安,2004;Fonti et al., 2010)。因此,研究树木年轮与气候因子之间的关系是探讨全球变化背景下树木和森林碳汇的重要指标(Fonti et al., 2010)。
目前,树木年轮与气候因子关系的研究区域主要集中于中高纬度地区(Z·ywiec et al., 2016;Babst et al., 2019;Camarero et al., 2021),而对亚热带等地区的研究相对较少(Luo et al., 2017;Huang et al., 2018;Liang et al., 2019)。亚热带森林是全球森林生态系统的重要组成部分,具有较高的固碳潜力(Locosselli et al., 2020),对全球气候调控发挥着不可或缺的作用(Yu et al., 2014)。近年来,因气候变化而导致亚热带地区旱季降水频率减少和雨季强降雨事件频发(Zhou et al., 2011, 2013)。为探讨气候变化对亚热带森林的影响,亟需开展该地区树木径向生长与气候关系的研究。由于亚热带地区气候较适宜树木生长,树木年轮的界限相对不明显,因此限制了该地区树木径向生长-气候关系的研究。尽管已有学者对亚热带树木径向生长与气候的关系进行了调查,但研究树种主要集中于马尾松(Pinus massoniana)和木荷(Schima superba)等少数树种(董志鹏等,2014;Luo et al., 2017;Liang et al., 2019)。因此,为更准确地预测未来亚热带森林树木的生长和碳汇,还需扩大对不同树种的相关研究。
广西金钟山黑颈长尾雉国家级自然保护区(以下简称金钟山保护区)位于广西壮族自治区最西端,属云贵高原边缘,对云贵高原与广西丘陵之间的物种扩散交流具有重要意义(姜灿荣,2008)。金钟山保护区地处南亚热带西部,具有亚热带常绿阔叶林东部湿润区向西部半湿润区过渡的特点,其独特地理位置和植物区系在科学研究上具有重要价值。细叶云南松(Pinus yunanensis var. tenuifolia)属松科松属常绿树种,是金钟山保护区唯一的天然针叶林-暖性针叶林的单一优势树种(黄元河等,2006)。目前,影响和制约金钟山细叶云南松径向生长的气候因子以及未来气候变化下其碳汇能力的变化情况尚不清楚。厘清这些问题对了解未来金钟山保护区天然针叶林的碳汇和发展具有重要意义。因此,本研究将从树木年轮学角度探讨金钟山保护区细叶云南松径向生长对气候的响应,以期为将来金钟山保护区天然针叶林的保护和管理提供理论依据。
1 材料与方法
1.1 样地概况
金钟山保护区(104°46′13″—105°00′46″ E、24°32′44″—24°43′07″ N)位于广西西北部、云贵高原东南缘。该保护区地处南亚热带西部,受来自云南高原焚风影响,具有干湿季明显的气候特点,属南亚热带季风气候区。境内平均气温17.1 ℃,霜期短。最冷月和最暖月分别为1月和7月。年降雨量1 200 mm,属广西降水量较少区域之一,雨量集中分布于5—9月,约占全年降水量的80%。土壤质地干硬,呈垂直分布,其中中山针阔混交林的土壤为山地黄壤。受云南高原焚风的影响,金钟山保护区具有亚热带常绿阔叶林东部湿润区向西部半湿润区过渡的特点,属亚热带常绿阔叶林西部半湿润区,其森林类型及组成和东部大不相同(苏宗明,1998)。暖性针叶林是金钟山保护区重要的森林类型之一,以细叶云南松为单优势种(苏宗明,1998;黄元河等,2006)。
1.2 树木年轮样品处理及数据获取
1.2.1 树木年轮样品采集与处理 本研究在细叶云南松分布较为集中的中山地段(1 100~1300 m)采样点随机选取样树,使用内径为5.12 mm的树木生长锥在树木胸高处(1.3 m)钻取树芯样本。每棵立木采集1个树木年轮样芯。样本的预处理过程依照传统方法(Stokes & Smiley, 1968)进行,待树芯样品自然风干后,将树芯白乳胶固定在带有槽沟的木槽上。将固定好的样芯依次采用100、280、320、600和800目的磨砂纸进行打磨,直至样芯的年轮界限在显微镜下清晰可辨。测量前根据样本最后一年的解剖学特征确定靠近树皮最外的年轮形成年,用铅笔从树皮开始按照倒推式的时间顺序进行年份初步标记。
1.2.2 树木年轮宽度测定及交叉定年 将所有样条通过LINTAB 6树木年轮宽度测量平台,使用TSAP-WinTM程序测量宽度,精度为0.001 mm。使用COFECHA软件,对树木年轮可视化交叉定年后的数据进行校正,利用50年窗口25年重叠的滑动相关系数作为检验指标。根据COFECHA程序检验结果和样芯实际生长情况,将所有可能存在定年问题,如缺轮、伪轮和断裂丢失片段等的样芯在显微镜下逐一检查和校对,并将未能定年的样芯剔除,最终获得36根成功定年的样芯用于后续分析。
1.3 气象数据获取
用于年轮对气候响应分析的气象数据从国家气象信息中心(http://data.cma.cn/)下载,选取离样点最近气象站(105°07′ E、24°07′ N)的月值数据集进行分析。气象数据包括月平均最高气温(Tmax)、月平均最低气温(Tmin)、月平均气温(Tmean)、月降雨量(precipitation)、月降雨量>10 mm的天数(Pre10)、月平均空气相对湿度(relative air humidity, RH)和月日照时数(sunshine duration, SD)。
1.4 统计分析
1.4.1 干旱指数计算和气候因子的年际变化趋势分析 标准化降水蒸散指数(standardized precipitation evapotranspiration index, SPEI)是应用最为广泛的干旱指数之一,综合考虑了降水和温度对干旱发生的共同效应以及不同时间尺度的降水和温度因素的累加效应,具有多时间尺度特性(Vicente-Serrano et al., 2010)。气象站逐月的SPEI采用R语言程序包SPEI1.7(https://cran.rproject.org/web/packages/SPEI/)计算。其中,1个月尺度SPEI值(SPEI.1)对短时间内的干旱变化情况比较敏感,能较为清晰地反映旱涝的细微性变化,3个月尺度SPEI值(SPEI.3)可以反映季节的干旱发生情况,2个月尺度SPEI值(SPEI.2)介于SPEI.1和SPEI.3之间。采用线性回归分析各气候因子的月值数据随年份的变化趋势。
1.4.2 树木年轮标准年表建立 由于年际间的树木年轮宽度同时受到气候和树木本身遗传因子的影响,因此在分析气候因子对树木径向生长的影响前需要除去树木本身的遗传信号,即年轮去趋势(Cook, 1985)。采用R软件“dplR”包的“detrend”函数对COFECHA交叉定年后的树木年轮宽度序列进行去趋势化(Bunn, 2008)。去趋势的方法采用平滑样条函数(67%样芯长度步长)进行,采用R软件“dplR”包的“chron”函数建立年表。此外,由于年表的可信度随着复本量而变化,即复本量越低可信度越低,因此为增加差值年表的可信度,选取样本量大于5根样芯且群体表达信号(express population signal, EPS)大于0.85的树木年轮宽度数据来建立年表,并记录年表的主要特征参数,如平均敏感度(mean sensitivity, MS)、信噪比(signal to noise ratio, SNR)和样芯间相关系数(mean inter-series correlation, rbar)等参数。
1.4.3 树木年轮对气候的响应分析 树木径向生长在很大程度上受到气候因子的影响,尤其是温度和降雨。本研究采用R软件中“treeclim”包的“dcc”函数来分析树木年轮生长与气候因子的关系。由于考虑到气候因子对树木年轮生长的影响具有滞后效应(Fritts, 1976),并且亚热带地区细叶云南松为常绿树种,因此研究选取先前一年生长季(1—12月)以及当年生长季(1—12月)每月的气象数据(时间跨度为24个月)进行树木年轮与气候关系的分析。为进一步量化气候因子对树木径向生长的影响,将筛选出与树木年轮标准年表有显著相关的气候因子,采用R“MASS”包中的“stepAIC”函数进行逐步回归,建立气候因子与树木年轮年表的线性模型。逐步回归从全模型开始,采用膨胀系数(VIF<4)对具有共线性的气候因子变量进行筛除,使用“stepAIC”函数对剔除共线性变量的模型进行逐步递减气候因子变量,依据AIC最小原则选择最终模型。采用R中的“relaimpo”包对最终模型进行方差分解,使用“boot.relimp”函数进行1 000次重复抽样计算置信区间,得出每个气候因子变量的方差解释量即对树木径向生长的贡献率。
本研究所有数据分析和图的绘制均使用R 4.2.0进行。
2 结果与分析
2.1 气象因子特征
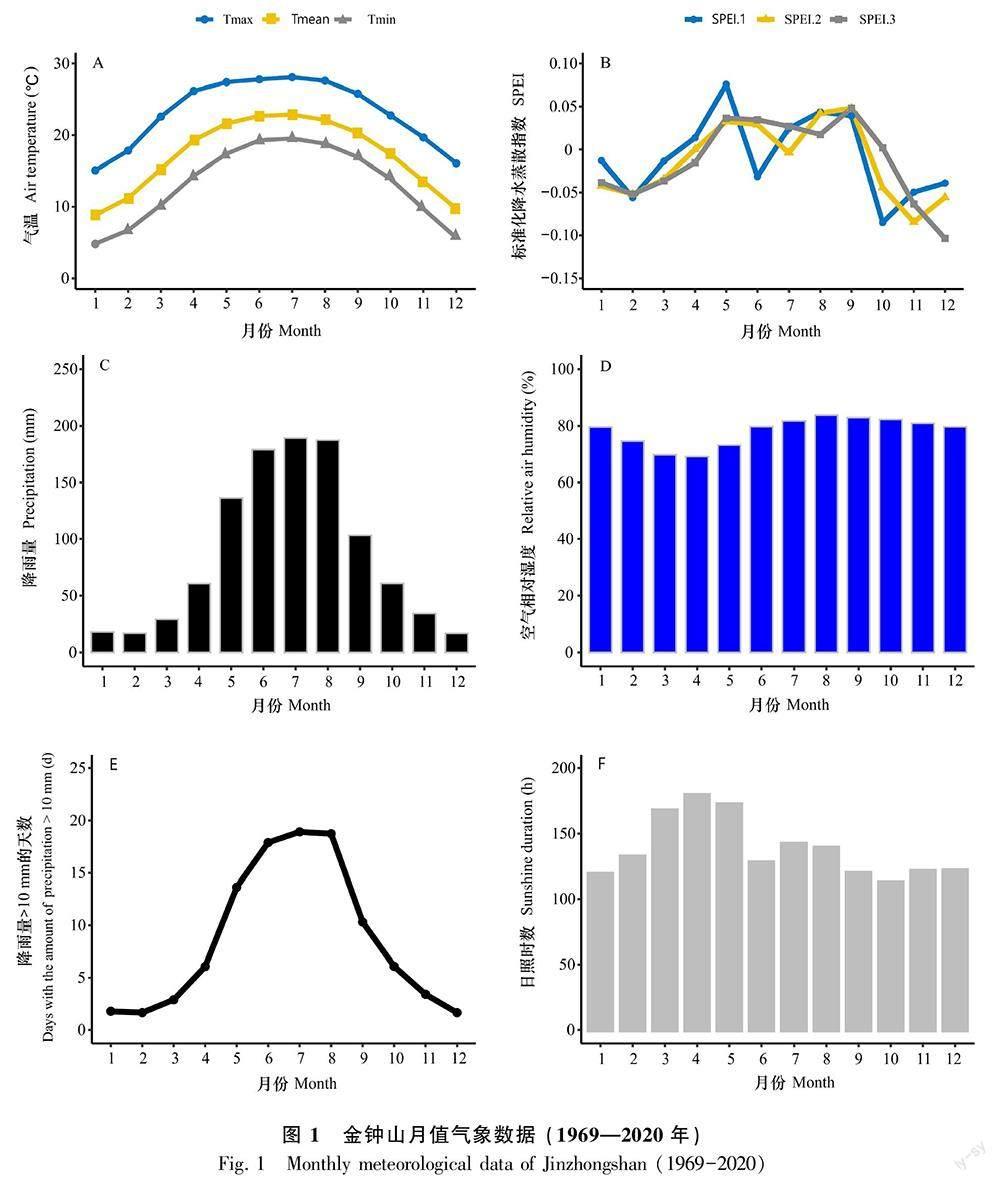
金钟山1969—2020年的月值气象数据如图1所示。6—8月的月平均最低气温和月平均最高气温和月降雨量均高于一年中的其他月份,12月至次年2月的月平均最低气温和月平均最高气温相比其他月份值要低(图1:A)。SPEI.1在2月和10月较其他月份值要低,而SPEI.2和SPEI.3分别在11月和12月最低(图1:B)。11月到次年3月的月降雨量(图1:C)和降雨量>10 mm的天數(图1:E)以及3—4月的平均空气相对湿度(图1:D)在全年中相对其他月份值要低。1月和10月的月平均日照时数在一年中最短(图1:F)。线性回归拟合1969—2020年月值气象数据随年际的变化趋势见表1。表1结果显示,5—9月和11月的月平均最高气温和全年(1—12月)月平均最低气温随年显著升高;与气温相反,5—9月和11月的空气相对湿度则随年呈显著下降趋势;11月的日照时数随年有显著增加趋势,而同期的SPEI.1、SPEI.2、SPEI.3和降雨量>10 mm的天数则有显著减少趋势。
2.2 树木年轮年表统计特征
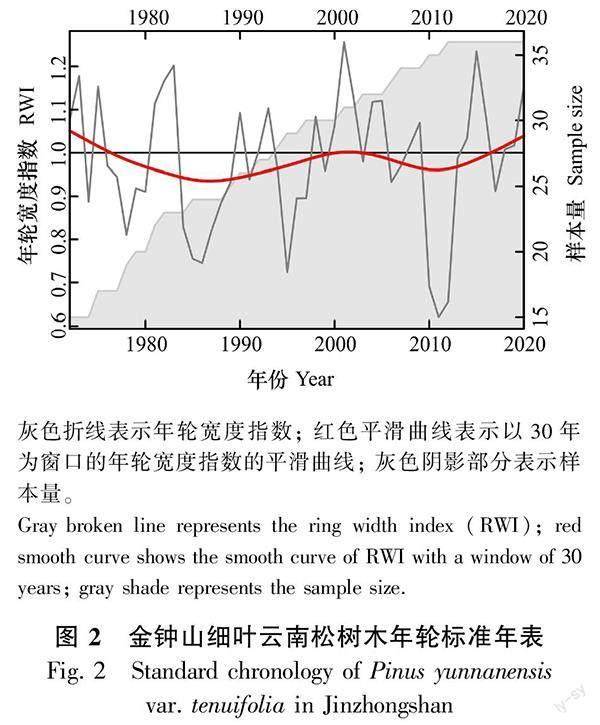
样树的年轮序列时间跨度为1910—2021年,平均年轮序列长度为49.25年。为使树木年轮标准年表具有区域代表性,选取样本数大于5且群体表达信号大于0.85的区间段进行分析(图2,表2)。年表公共区间时间跨度为1972—2020。在公共区间段内,样芯平均敏感度为0.34,表明年轮时间序列对气候信息的敏感程度相对较高,符合树木年轮研究标准;一阶自相关系数(AR1)为0.45,表明前一年气候对当年年轮宽生长有一定影响;群体间表达信号为0.86,达到区域代表性的标准。
2.3 树木年轮年表对气候的响应特征
树木年轮年表和气候因子的相关分析结果(图3)显示,前一年的气候因子对树木年轮生长有显著影响,其中上一年1月、8月、9月和11月的日照时数(SD)、上一年10月的平均最低气温(Tmin)和平均气温(Tmean)与树木年轮年表宽度呈显著正相关,而上一年6月降雨量>10 mm的天数(Pre10)与树木年轮年表宽度呈显著负相关。当年2月的降雨量(Precipitation)、3月的空气相对湿度(RH)和平均最低气温(Tmin)、6月的日照时数(SD)及9月的平均最高气温(Tmax)和平均气温(Tmean)均与树木年轮年表宽度呈显著正相关。
气候因子对细叶云南松树木径向生长的贡献率(即模型方差总解释量)为44.27%(图4)。气温是影响细叶云南松树木年轮宽度最显著的气候因子,其中上一年10月的平均最低气温(Tmin.prev.oct.)影响最大(方差解释量达23.35%),其次是当年9月的平均最高气温(Tmax.curr.sep.),其方差解释量为10.39%。光照是影响细叶云南松树木径向生长的另一个重要气象因子, 其总体贡献率达10.52%,其中上一年1月的日照时数(SD.prev.jan.)和11月的日照时数(SD.prev.nov.)分别解释了细叶云南松径向生长变异的3.94%和6.58%。可见,气温是影响金钟山细叶云南松径向生长最主要的气候因子,其次是光照。
3 讨论
树木的径向生长不仅受到树木本身遗传因素的影响, 还可能受到各种气候因子的调控 (Fritts, 1976;Plomion et al., 2001;Rathgeber et al., 2016)。本研究发现,前一年的温度和光照对细叶云南松树木年轮生长有显著滞后效应;气候因子中对细叶云南松径向生长变异贡献最大的为上一年10月的平均最低气温,其次是当年9月的平均最高气温以及上一年1月和11月的日照时数。
3.1 温度是影响细叶云南松径向生长的主要气候因子
温度是影响树木径向生长最主要的气候因子之一 (Delpierre et al., 2019)。树木径向生长常常受益于生长季早期温度的升高。一方面,树木径向生长所需的非结构性碳水化合物(non-structural carbohydrates, NSC)的产生依赖于叶片光合作用(Michelot et al., 2012),而光合作用依赖于叶绿素和光合酶的作用,生长季温度的升高可加快光合酶的功能(Shi et al., 2015),增加木質部生长所需的碳水化合物累积。另一方面,生长季早期温度的升高可以通过增强树木根系的活力,如增强对营养物质和水分的吸收来促进树木对营养物质的吸收以及加速蒸腾、增大气孔导度等,最终促进光合同化作用,有利于碳水化合物累积(Poorter et al., 2012;Urban et al., 2017)。本研究发现,金钟山细叶云南松与当年3月(生长季早期)的月平均最低气温呈显著正相关,这与Yang等(2022)研究发现气候干冷条件下云南松(Pinus yunnanensis)径向生长主要受低温限制的结果相似。
本研究还发现,上一年10月的平均最低气温和当年9月的平均最高气温均与细叶云南松的树木年轮宽度呈正相关,并且两者对树木年轮宽度生长的贡献率在气候因子中最大。Huang等(2018)对与细叶云南松同属不同种的南亚热带森林马尾松的研究发现,进入干季后,广东省鼎湖山和石门台国家自然保护区的马尾松木质部的生长速率最快,在干季(10月至次年3月)分别形成58%和39%的年轮宽度,显示了较高的碳积累,表明南亚热带干季适宜的温湿条件相对于湿季更有利于马尾松径向生长。这与中国亚热带森林的净生态系统生产力估算结果相似,即干旱年(2004年)中国亚热带森林系统的碳汇积累量比湿润年(2008年)高出81.4%,这可能与湿季雨水较多导致树木呼吸增加,从而降低碳水化合物累积有关(Yan et al., 2013)。因此,南亚热带干季的温湿条件可能较湿季更适合树木进行碳累积。本研究区9—10月温度升高可能有助于产生干季相对适宜的环境来促进树木的碳累积,该阶段气温与细叶云南松径向生长呈正相关。此外,树木年轮宽度的一部分可由贮藏的NSC支撑(Skomarkova et al., 2006)。前人研究发现,树木径向生长对前一年的环境条件做出响应(Fonti & Garcia-Gonzalez, 2004),在很大程度上归因于NSC对木质部生长的限制作用(Michelot et al., 2012)。本研究上一年10月温度的升高可潜在地增加NSC的累积和贮存,提高下一年早材形成所需的物质和能量储备。因此,上一年10月温度对当年的树木年轮宽度具有促进作用。
3.2 光照对细叶云南松径向生长的影响
光合作用的能量驱动来自光照,日照时数是影响树木径向生长的另一个重要气候因素(Huang et al., 2020)。日照时数增加会导致植物吸收的热量增加(陆佩玲,2006)。在温度较低的环境中,径向生长过程中形成层的活动与日照时数同步,即光照时间越长生长季早期形成层分裂速度越快(Duchesne et al., 2012)。本研究区1月的日照时数短、温度低,阴冷环境会限制树木的光合作用;寒冷条件下针叶树发芽之前木质部形成的主要能量来源于前一年储藏的非结构碳(Rossi et al., 2009;Huang et al., 2014),使得生长季早期光合作用固定的碳水化合物对树木生长起重要作用(Hansen & Beck, 1994)。因此,上一年1月份的日照时数与细叶云南松径向生长呈正相关。同样,上一年11月份的日照时数与细叶云南松径向生长呈正相关,这可能与11月较低的光照和温度会限制树木的光合作用有关。此外,在降雨高峰期,由于光照相对缺乏,影响了树木蒸腾作用和碳吸收,会降低树木径向生长所需的碳水化合物产量,因此6月日照时数与细叶云南松径向生长呈正相关。
3.3 降雨对细叶云南松径向生长的影响
由于可供树木生长利用的水分主要来自降水,地下水补给的作用较小(Ehleringer, 1985),因此降雨成为限制植物生长的另一个关键气候要素(Z·ywiec et al., 2016)。一方面,通过降雨影响光合速率和碳分配间接影响细胞分裂和扩大(Zweifel et al., 2006),导致土壤水分增加,促进树干对水分的运输,有利于光合作用(Myburg & Sederoff, 2001)。另一方面,降雨过程进入树木体内的水分直接参与形成层活动和细胞扩大(Barbaroux & Breda, 2002;Steppe et al., 2015),通过增加形成层分裂时细胞壁弯曲所需的膨压,促进木质部细胞的分化和延伸(Turcotte et al., 2009;Steppe et al., 2015)。Yang等(2022)研究表明,干燥环境下云南松径向生长受生长季早期的水分有效性限制。由于本研究区2月份的降雨量较少,加上不断升高的温度使研究区相对较干(SPEI.1较低),因此该阶段的降雨量增加有利于树木径向生长,这可能是本研究中当年2月份降雨量与树木径向生长呈正相关的一个原因。而本研究发现,上一年6月降雨量>10 mm的天数与细叶云南松径向生长量呈显著负相关。过量的降雨会增加叶片湿润状态,使气孔关闭(Aparecido et al., 2017),从而导致进入叶片气孔的二氧化碳浓度降低;同时,降雨过多会导致光照缺乏,影响树木蒸腾作用和光合作用,降低树木生长所需的碳水化合物产量。此外,降雨量过多还会导致土壤因过于潮湿而不利于根系对水分和无机盐吸收(董志鹏等,2014),最终对树木径向生长产生负面影响。这可能是本研究中雨季6月降雨量>10 mm的天数与细叶云南松径向生长呈负相关以及6月日照时数与细叶云南松径向生长呈正相关的原因。这与Yang等(2022)的研究结果一致,即在温湿区域云南松径向生长受制于过量的水分。
综上所述,金钟山细叶云南松径向生长既得益于秋季的升温又受限于秋季和春季的低温。秋季温度是影响细叶云南松径向生长最主要的气候因子,其次为冬季光照。在降雨量相对缺乏的早春,降雨和空气湿度增加有利于金钟山细叶云南松的径向生长,而雨季过多的降雨则会限制细叶云南松的径向生长。此外,由于金钟山地区秋季气温具有增加趋势,预计未来在气温继续增加的背景下,细叶云南松的径向生长将得到促进,金钟山天然针叶林的碳汇也将有所增加。
致谢 本研究野外树木年轮采样工作得到广西金钟山黑颈长尾雉国家级自然保护区的大力支持,室内年轮样品处理得到南宁师范大学本科生卢湫水、唐莉莎、邓江梅和顾悦的支持,同时也得到广西第八批特聘专家专项(2019B16)資助,在此一并表示感谢!
参考文献:
APARECIDO LM, MILLER GR, CAHILL AT, et al., 2017. Leaf surface traits and water storage retention affect photosynthetic responses to leaf surface wetness among wet tropical forest and semiarid savanna plants [J]. Tree Physiol, 37(10): 1285-1300.
BABST F, BOURIAUD O, POULTER BI,et al., 2019. Twentieth century redistribution in climatic drivers of global tree growth [J]. Sci Adv, 5(1): eaat4313.
BARBAROUX C, BREDA N, 2002. Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees [J]. Tree Physiol, 22(17): 1201-1210.
BUNN AG, 2008. A dendrochronology program library in R (dplR) [J]. Dendrochronologia, 26(2): 115-124.
CAMARERO JJ, GAZOL A, SNCHEZ-SALGUERO R, et al., 2021. Global fading of the temperature-growth coupling at alpine and polar treelines [J]. Glob Change Biol, 27(9): 1879-1889.
COOK ER, 1985. A time series analysis approach to tree ring standardization [D]. Tucson: University of Arizona.
DELPIERRE N, LIREUX S, HARTIG F, et al., 2019. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers [J]. Glob Change Biol, 25(3): 1089-1105.
DONG ZP, ZHENG HZ, FANG KY, et al., 2014. Responses of tree-ring width of Pinus massoniana in Sanming, Fujian Province to climate change[J]. J Subtrop Res Environ, 9(1): 1-7.[董志鹏, 郑怀舟, 方克艳, 等, 2014. 福建三明马尾松树轮宽度对气候变化的响应 [J]. 亚热带资源与环境学报, 9(1): 1-7.]
DUCHESNE L, HOULE D, D’ORANGEVILLE L, 2012. Influence of climate on seasonal patterns of stem increment of balsam fir in a boreal forest of Québec, Canada [J]. Agric For Meteorol, 162/163: 108-114.
EHLERINGER J, 1985. Annuals and perennials of warm deserts [M]//CHABOT BF, MOONEY HA. Physiological ecology of North American plant communities [C]. New York: Chapman and Hall: 162-180.
FAN WY,WANG XA, 2004. Advances in studies on the relationships between tree-ring width and climatic factors [J]. Acta Bot Boreal-Occident Sin, 24(2): 345-351.[范玮熠, 王孝安, 2004. 树木年轮宽度与气候因子的关系研究进展 [J]. 西北植物学报, 24(2): 345-351.]
FONTI P, GARCIA-GONZALEZ I, 2004. Suitability of chestnut earlywood vessel chronologies for ecological studies [J]. New Phytol, 163(1): 77-86.
FONTI P, VON ARX G, GARCIA-GONZALEZ I,et al., 2010. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings [J]. New Phytol, 185(1): 42-53.
FRITTS HC, 1976. Tree ring and climate [M]. London: Academic Press.
HANSEN J, BECK E, 1994. Seasonal changes in the utilization and turnover of assimilation products in 8-year-old Scots pine (Pinus sylvestris L.) trees [J]. Trees, 8(4): 172-182.
HUANG JG, DESLAURIERS A, ROSSI S, 2014. Xylem formation can be modeled statistically as a function of primary growth and cambium activity [J]. New Phytol, 203(3): 831-841.
HUANG JG, GUO X, ROSSI S, et al., 2018. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season [J]. Tree Physiol, 38(8): 1225-1236.
HUANG JG, MA Q, ROSSI S,et al., 2020. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers [J]. Proc Natl Acad Sci USA, 117(34): 20645-20652.
HUANG YH, TANG SQ, LU ZY, et al., 2006. Characteristics of main vegetation types in Jinzhongshan Nature Reserve, Guangxi Province [J]. J Trop Subtrop Bot, 14(6): 499-503.[黃元河, 唐绍清, 卢贞燕, 等, 2006. 广西金钟山自然保护区主要植被类型的特征 [J]. 热带亚热带植物学报, 14(6): 499-503.]
JIANG CR, 2008. On protection value of Guangxi Jinzhongshan National Nature Reserve [J]. Cent S For Invent Plan, 27(4): 37-39.[姜灿荣, 2008. 论广西金钟山国家级自然保护区的保护价值 [J]. 中南林业调查规划, 27(4): 37-39.]
LEWIS SL, WHEELER CE, MITCHARD E, et al., 2019. Regenerate natural forests to store carbon [J]. Nature, 568(7750): 25-28.
LIANG HX, HUANG JG, MA QQ, et al., 2019. Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China [J]. Agric For Meteorol, 274: 7-17.
LOCOSSELLI GM, BRIENEN RJW, LEITE MS,et al., 2020. Global tree-ring analysis reveals rapid decrease in tropical tree longevity with temperature [J]. Proc Natl Acad Sci USA, 117(52): 33358-33364.
LU PL, 2006. Study of the response of main woody plant phenology to climate change in China [D]. Beijing: Beijing Forestry University.[陸佩玲, 2006. 中国木本植物物候对气候变化的响应研究 [D]. 北京: 北京林业大学.]
LUO DW, HUANG JG, JIANG XY, et al., 2017. Effect of climate and competition on radial growth of Pinus massoniana and Schima superba in China’s subtropical monsoon mixed forest [J]. Dendrochronologia, 46: 24-34.
MICHELOT A, SIMARD S, RATHGEBER C,et al., 2012. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics [J]. Tree Physiol, 32(8): 1033-1045.
MYBURC AA, SEDEROFF RR, 2001. Xylem structure and function [M/OL]//
Encyclopedia of Life Sciences. Nature Publishing Group. http://dx.doi.org/10.1038/npg.els.0001302.
PLOMION C, LEPROVOST GG, STOKES A, 2001. Wood Formation in Trees [J]. Plant Physiol, 127(4): 1513-1523.
POORTER H, NIKLAS KJ, REICH PB, et al., 2012. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control [J]. New Phytol, 193(1): 30-50.
RATHGEBER CB, CUNY HE, FONTI P, 2016. Biological basis of tree-ring formation: A crash course [J]. Front Plant Sci, 7: 734.
ROSSI S, ANFODILLO T, CUFAR K, et al., 2016. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere [J]. Glob Change Biol, 22(11): 3804-3813.
ROSSI S, RATHGEBER CBK, DESLAURIERS A, 2009. Comparing needle and shoot phenology with xylem development on three conifer species in Italy [J]. Ann For Sci, 66(2): 206-206.
SASS-KLAASSEN U, 2015. Tree physiology: Tracking tree carbon gain [J]. Nat Plants, 1: 15175.
SHI CG, SILVA LCR, ZHANG HX, et al., 2015. Climate warming alters nitrogen dynamics and total non-structural carbohydrate accumulations of perennial herbs of distinctive functional groups during the plant senescence in autumn in an alpine meadow of the Tibetan Plateau, China [J]. Agric For Meteorol, 200: 21-29.
SKOMARKOVA MV, VAGANOV EA, MUND M,et al., 2006. Inter-annual and seasonal variability of radial growth, wood density and carbon isotope ratios in tree rings of beech (Fagus sylvatica) growing in Germany and Italy [J]. Trees, 20(5): 571-586.
STEPPE K, STERCK F, DESLAURIERS A, 2015. Diel growth dynamics in tree stems: linking anatomy and ecophysiology [J]. Trends Plant Sci, 20(6): 335-343.
STOKES MA, SMILEY TL, 1968. An introduction to tree-ring dating [M]. Chicago: The University of Chicago Press: 73.
SU ZM, 1998. The classified system of natual vegetation in Guangxi [J]. Guihaia, 18(3): 237-246.[苏宗明, 1998. 广西天然植被类型分类系统 [J]. 广西植物, 18(3): 237-246.]
TURCOTTE A, MORIN H, KRAUSE C,et al., 2009. The timing of spring rehydration and its relation with the onset of wood formation in black spruce [J]. Agric For Meteorol, 149(9): 1403-1409.
URBAN J, INGWERS M, MCGUIRE MA,et al., 2017. Stomatal conductance increases with rising temperature [J]. Plant Signal Behav, 12(8): e1356534.
VICENTE-SERRANO SM, BEGUERA S, LPEZ-MORENO JI, 2010. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index [J]. J Clim, 23(7): 1696-1718.
YAN JH, LIU XZ, TANG XL, et al., 2013. Substantial amounts of carbon are sequestered during dry periods in an old-growth subtropical forest in South China [J]. J For Res, 18(1): 21-30.
YANG RQ, FU PL, FAN ZX,et al., 2022. Growth-climate sensitivity of two pine species shows species-specific changes along temperature and moisture gradients in southwest China [J]. Agric For Meteorol, 318: 108907.
YU GR, CHEN Z, PIAO SL, et al., 2014. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region [J]. Proc Natl Acad Sci USA, 111(13): 4910-4915.
ZHOU GY, PENG CH, LI YL, et al., 2013. A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China [J]. Glob Change Biol, 19(4): 1197-1210.
ZHOU GY, WEI XH, WU YP, et al., 2011. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China [J]. Glob Change Biol, 17(12): 3736-3746.
ZWEIFEL R, ITEM H, HSLER R, 2006. Link between diurnal stem radius changes and tree water relations [J]. Tree Physiol, 21: 869-877.
Z·YWIEC M, MUTER E, ZIELONKA T, et al., 2016. Long-term effect of temperature and precipitation on radial growth in a threatened thermo-Mediterranean tree population [J]. Trees, 31(2): 491-501.
(責任编辑 蒋巧媛)
