元宝枫籽象的重要天敌昆虫
——元宝枫刺胫广肩小蜂线粒体全基因组的测定与分析
2023-10-30刘慧慧李恩杰曹亮明包青春王小林辛学兵杨忠岐
刘慧慧, 李恩杰, 曹亮明, 包青春, 王小林, 辛学兵, 杨忠岐,*
(1. 中国林业科学研究院华北林业实验中心, 北京九龙山暖温带森林国家长期科研基地, 北京102300;2. 中国林业科学研究院森林生态环境与保护研究所, 国家林业和草原局森林保护学重点实验室, 北京100091;3. 内蒙古科尔沁右翼中旗五角枫自然保护区管理局, 兴安盟 137400)
广肩小蜂科(Eurytomidae)在全世界广泛分布,目前分为3个亚科:海广肩小蜂亚科(Hemibrinae)、肿腿广肩小蜂亚科(Rileyinae)和广肩小蜂亚科(Eurytominae)(Gauld and Bolton, 1988; 杨忠岐, 1992)。该科全世界记载了87属1 500多种(Gatesetal., 2006; Gates, 2007; Lotfalizadehetal., 2007; Noyes, 2023),其中绝大部分属于广肩小蜂亚科。广肩小蜂科大部分种类为寄生性,少数为植食性。广肩小蜂属Eurytoma是该科中最大的一个属,其大多数种类寄生于其他昆虫(绝大多数为害虫)体上,寄主类群包括鞘翅目(Coleoptera)、双翅目(Diptera)、鳞翅目(Lepidoptera)、膜翅目(Hymenoptera)等昆虫。瘿广肩小蜂属Sycophila是该科中的另外一个属,主要寄生于膜翅目瘿蜂科(Cynipidae)昆虫。截至目前,全世界已知广肩小蜂属702种和瘿广肩小蜂属119种(Gauld and Bolton, 1988; Gates, 2007; Noyes, 2023)。由于广肩小蜂属具有重要的生物防治价值,应对其进行深入研究,以发掘该天敌类群的生物防治价值和利用于生物防治。
元宝枫Acertruncatum属于槭树科(Aceraceae)槭树属Acer多年生落叶亚乔木,作为我国特有的珍贵木本油料树种,不仅是我国园林绿化重要树种,也被广泛用于荒山造林和治理荒漠化。元宝枫材质优良,在我国分布广,适应性强,其种子和叶片等产生的多种产品已经广泛用于化工、医药、食品、化妆品等领域。更有价值的是,元宝枫籽油中含有丰富的神经酸(顺-15-二十四碳单烯酸),在治疗和预防人类神经系统疾病上具有良好的药用和保健作用,我国目前正在开展利用元宝枫籽油研制治疗帕金森症、老年痴呆症等神经性疾病的药物(王性炎, 2013)。然而,我国最大的元宝枫采种基地——内蒙古科尔沁沙漠4 000 hm2的天然元宝枫林,近年来遭受一种象甲的严重危害,经我们调查,明确其为象甲科的元宝枫籽象Bradybatussp.。这种害虫在元宝枫翅果中蛀食元宝枫种仁。2021年经我们调查显示,内蒙古兴安盟代钦塔拉保护区85%以上元宝枫种仁被其蛀食一空,导致元宝枫种子几乎绝收。为了利用天敌对这种隐秘性生活的重大害虫进行无公害防治,我们同时也对元宝枫籽象天敌区系进行了调查,发现1种寄生其幼虫、蛹甚至刚羽化的成虫的小蜂。经过我们分类研究,确定该物种为广肩小蜂科(Eurytomidae)广肩小蜂亚属Eurytoma的一个新种——元宝枫刺胫广肩小蜂EurytomaacutibialisYang, LiuetCao。该种对元宝枫籽象寄生率高达45%,在自然控制该害虫上发挥着重要作用。鉴于元宝枫刺胫广肩小蜂是元宝枫重大种实害虫元宝枫籽象的优势寄生蜂,为了进一步明确该小蜂的分类地位和其与相近种的亲缘关系,我们测定和分析了该小蜂的线粒体全基因组序列,为膜翅目小蜂总科广肩小蜂科昆虫系统分类和发育关系等研究提供分子生物学证据。
昆虫线粒体基因组具有母系遗传、长度短、排列较为保守和进化速率快等特点,被广泛应用于膜翅目昆虫的物种界定、进化和系统发育等研究中(Dowtonetal., 2009; Cameron, 2014; Maoetal., 2015; Tangetal., 2019)。随着测序技术的成熟和分子生物学的快速发展,越来越多的膜翅目昆虫物种的线粒体全基因组被测序。然而,Chen等(2004)利用线粒体16S rRNA基因和cox1对广肩小蜂科进行系统进化分析时发现,无法很好地支持各亚属的单系性。目前NCBI基因数据库仅收录了广肩小蜂科3种昆虫的线粒体基因组信息,分别是广肩小蜂(Eurytomasp. ZJUH 2013和Eurytomasp. TJS-2016)和瘿广肩小蜂(Sycophilasp. 2 JXW 2020),广肩小蜂科昆虫线粒体基因组的数据还非常有限。本研究通过测定元宝枫刺胫广肩小蜂线粒体基因组,对其线粒体基因组结构、基因排列顺序、核苷酸组成、氨基酸组成、密码子使用、tRNA基因二级结构、基因组重排现象等进行分析;此外,利用40种膜翅目寄生蜂线粒体基因组的13个蛋白质编码基因(protein-coding genes, PCGs)进行系统发育分析,以探究元宝枫刺胫广肩小蜂所在属的高级阶元的分类地位。这些研究在建立广肩小蜂科的自然分类系统方面具有重要意义。
1 材料与方法
1.1 昆虫收集
2021年10月,在内蒙古兴安盟代钦塔拉五角枫自然保护区(45°18′N, 121°02′E)采集元宝枫翅果约1 500粒,将这些翅果置于养虫笼中,放置于室外越冬(模拟自然界越冬情况)。2022年3月将翅果转移到室内,在温度(25±1) ℃、相对湿度40%~50%、光周期 14L∶10D的人工气候箱中培养,每天检查、收集羽化出的寄生蜂,在实体显微镜下鉴定这些寄生蜂种类和区分性别,并统计数量等,将收集元宝枫刺胫广肩小蜂成虫利用液氮速冻,随后储存于-80 ℃冰箱中保存备用。
1.2 线粒体基因组Illumina HiSeqTM4000高通量测序和组装
挑选30头元宝枫刺胫广肩小蜂,使用DNeasy组织试剂盒(Qiagen,Hilden,德国)提取全基因组DNA。使用1 μg DNA,利用TruSeq DNA文库制备试剂盒构建DNA文库。在上海派森诺生物科技股份有限公司利用Illumina HiSeqTM4000(Illumina, 美国)平台对元宝枫刺胫广肩小蜂样品进行双末端(paired-end, PE)测序,共获得3.35 Gb的原始数据,Q30为92.89%。利用SPAdes v3.10.1(http:∥bioinf.spbau.ru/spades)(Bankevichetal., 2012)和A5-miseq v2015052229(Coiletal., 2015)对原始数据的读段进行拼装,得到contig和scaffold序列。根据拼接序列的测序深度提取序列,将高测序深度的序列同NCBI上的nt数据库进行blastn (https:∥ftp.ncbi.nlm.nih.gov/blast/executables/blast+/LATEST/)比对,筛选出各拼接结果的线粒体序列。利用mummer v3.1(Kurtzetal., 2004)软件进行共线性分析,确定contig之间的位置关系,并填补contigs之间的gap。最后,使用pilon v1.18(Walkeretal., 2014)软件进行校正,得到最终的线粒体序列。
1.3 线粒体基因组序列注释与特征分析
利用MITOS(http:∥mitos.bioinf.uni-leipzig.de/index.py)对元宝枫刺胫广肩小蜂线粒体基因组进行注释和tRNA二级结构预测。注释结果与NCBI数据库中的同源序列进行比较得到确认,然后将结果提交至NCBI。采用CGview软件绘制线粒体基因组圈图。使用MEGA-X(Kumaretal., 2018)计算基因的核苷酸组成,包括A+T含量、AT偏斜、GC偏斜,AT偏斜=(A-T)/(A+T),GC偏斜=(G-C)/(G+C)。并使用Phylosuite v1.2.2(Zhang Detal., 2020)分析PCGs相对同义密码子使用率(relative synonymous codon usage, RSCU)以及PCGs密码子第1, 2和3位的G+C含量。以祖先昆虫亚库巴果蝇Drosophilayakuba线粒体基因组的排列方式为模式(Clary and Wolstenholme, 1985)对40种膜翅目昆虫线粒体基因组基因结构进行对比分析。
1.4 系统发育分析
以细蜂总科(Proctotrupoidae)的长腹细蜂Pelecinuspolyturator和离颚细蜂Vanhorniaeucnemidarum及姬蜂总科(Ichneumonoidea)的前裂长管茧蜂Diachasmimorphalongicaudata线粒体基因组为外群,基于40种膜翅目昆虫的13个PCGs核苷酸序列,使用贝叶斯法(Bayesian inference, BI)和最大似然法(maximum likelihood, ML)两种方法构建系统发育树。所有线粒体基因组序列从NCBI获得。使用 Phylosuite软件提取小蜂全序列中的13个PCGs的核苷酸序列,利用MAFFT 7.149(Katoh and Standley, 2013)和MACSE v. 2.03(Ranwezetal., 2018)对13个PCGs进行密码子比对和优化,最后利用Gblocks v0.91b(Castresana, 2000)对序列进行串联处理。利用ModelFinder(Kalyaanamoorthyetal., 2017)进行模型分析,然后利用XSEDE (https:∥www.phylo.org/)贝叶斯法和最大似然法构建系统发育树,采用Bootstrap(1 000次重复)检验系统树各分支节点的置信值。利用ITOL(https:∥itol.embl.de/personal_page.cgi)和adobe illustrator对系统发育树进行美化。
2 结果
2.1 元宝枫刺胫广肩小蜂线粒体基因组全序列
元宝枫刺胫广肩小蜂线粒体全基因组长15 849 bp (GenBank登录号: OQ473654),A+T含量82.00%,包括43.60% T,38.40% A,11.00% G和6.90% C,AT偏斜为-0.063,GC偏斜为0.230;线粒体基因组由37个典型基因组成,包括2个rRNA基因[rrnS(12S rRNA)和rrnL(16S rRNA)]、13个PCGs、22个tRNA基因和1个非编码控制区(control region, CR)(图1)。其中重链(J)的基因27个,分别是2个rRNA基因、11个PCGs(nad1,nad2,nad3,nad4,nad4L,nad5,cox1,cox2,cox3,atp6和atp8)、14个tRNA基因(trnI,trnV,trnA,trnL1,trnP,trnM,trnH,trnF,trnL2,trnD,trnG,trnC,trnR和trnW)和1个控制区。由轻链(N)编码的基因共10个,包括2个PCGs(cytb和nad6)和8个tRNA基因(表1)。
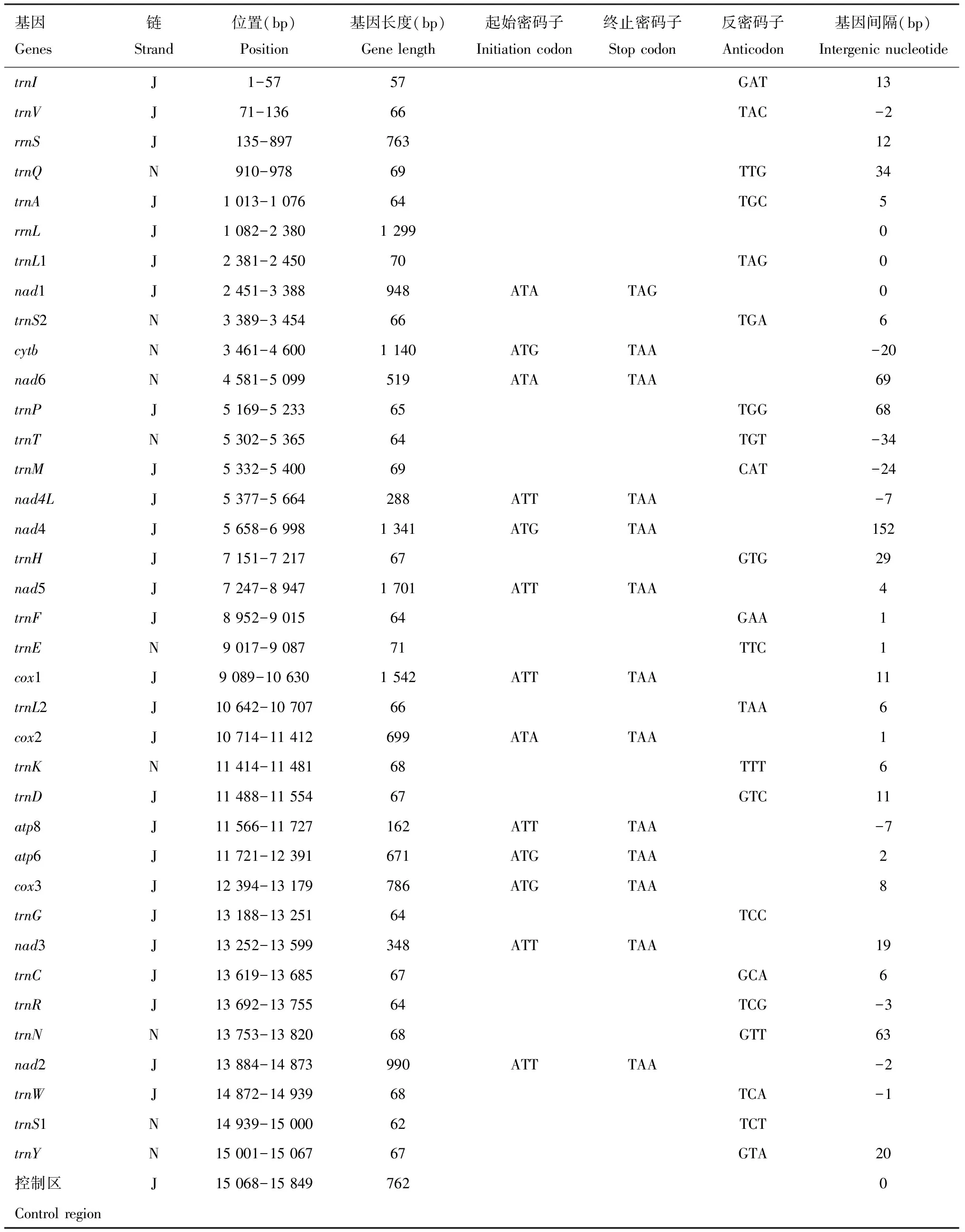
表1 元宝枫刺胫广肩小蜂线粒体基因组特征Table 1 Characteristics of the mitochondrial genome of Eurytoma acutibialis

图1 元宝枫刺胫广肩小蜂线粒体基因组结构Fig. 1 Mitochondrial genomic structure of Eurytoma acutibialis
元宝枫刺胫广肩小蜂线粒体所有PCGs均以ATN为起始密码子,其中6个PCGs (nad2,nad3,nad4L,nad5,cox1和atp8)的起始密码子为ATT,4个PCGs(cox3,nad4,atp6和cytb)的起始密码子为ATG,3个PCGs (nad1,nad6和cox2)的起始密码子为ATA。所有PCGs均以传统的TAA/TAG为终止密码子(表1)。
线粒体基因组整个编码区相邻基因之间存在基因间隔或基因重叠现象。其中基因间隔区有23处,总长度为 547 bp (基因间隔范围:1~152 bp),其中nad4和trnH之间间隔最长为152 bp,其次是trnP和nad6之间,基因间隔为69 bp;基因重叠有9处,总长度为100 bp (基因重叠范围:1~34 bp),最长一处位于trnT和trnM之间,重叠序列为34 bp; 此外,还有5个区域既无重叠又无间隔(表1)。
2.2 4种广肩小蜂PCGs
元宝枫刺胫广肩小蜂的13个PCGs全长为11 133 bp,占整个线粒体基因组序列全长的70.20%,13个PCGs的长度范围为162 (atp8)~1 701 bp (nad5)。A+T含量为80.90%,包括44.60% T, 36.30% A, 9.65% G和9.40% C,AT偏斜为-0.103,GC偏斜为0.010。在密码子第1位,G+C含量为22.90%,AT偏斜为0.044,GC偏斜为0.188;在密码子第2位,G+C含量为26.15%,AT偏斜为-0.351,GC偏斜为-0.118;在密码子第3位,G+C含量为8.05%,AT偏斜为-0.029,GC偏斜为-0.001(表2)。其他3种广肩小蜂也表现出明显的AT偏好性,例如,广肩小蜂Eurytomasp. ZJUH 2016013的13个PCGs全长为11 169 bp,A+T含量为81.40%,AT偏斜为-0.104,GC偏斜为-0.013(其线粒体基因组全长17 267 bp,A+T含量为84.40%,AT偏斜为0.074,GC偏斜为-0.239)。瘿广肩小蜂Sycophilasp. 2 JXW 2020的13个PCGs全长为11 115 bp,A+T含量为80.00%,AT偏斜为-0.136,GC偏斜为0.014(其线粒体基因组全长14 665 bp,A+T含量为81.40%,AT偏斜为-0.160,GC偏斜为0.235)。广肩小蜂Eurytomasp. TJS-2016的13个PCGs全长11 232 bp,A+T含量为80.80%,AT偏斜为-0.118,GC偏斜为-0.017(其线粒体基因组全长14 946 bp,A+T含量为82.60%,AT偏斜为0.083,GC偏斜为-0.154)。

表2 元宝枫刺胫广肩小蜂线粒体基因组核苷酸组成Table 2 Nucleotide composition of the mitochondrial genome of Eurytoma acutibialis
4种广肩小蜂13个PCGs的氨基酸使用率(图2)的结果显示,元宝枫刺胫广肩小蜂E.acutibialis使用占比由高到低氨基酸排序依次为Leu(15.57%),Ile(11.17%),Met(9.84%),Phe(9.35%)和Asn(6.92%)(总数>200),对应的密码子使用次数由高到低依次为UUA(492), AUU(402), UUU(336), AUA(325)和AAU(242);而Arg(1.30%), Cys(0.92%)和Gln(1.32%)的使用相对较少(总数<50)。除Phe使用率高于Met外,其他3种广肩小蜂结果与元宝枫刺胫广肩小蜂一致(图2)。结果显示,UUA的使用频率最高,RSCU值为5.12, ACG, AGC, GUC, CUG和CUC的相对密码子使用频率最低,其中RSCU值均为0.02。
其他3种光肩小蜂均缺少苏氨酸Thr的ACG密码子,E.sp. ZJUH 2016013缺少亮氨酸Leu的CUC和丝氨酸Ser的AGC密码子,以及Sycophilasp.2 JXW 2020缺少精氨酸Arg的CGC密码子,其他氨基酸使用的密码子均相同,密码子使用率排序基本一致,只有Sycophilasp. 2 JXW 2020的UUU使用率高于AUU(图2),这反映了核苷酸组成的AT偏好性。
2.3 tRNA基因、rRNA基因和控制区
元宝枫刺胫广肩小蜂线粒体基因组包含22个tRNA基因,总长为1 453 bp,占整个线粒体基因组全长的 9.17%(长度范围: 57~71 bp)。其中有14个tRNA基因由重链编码,其余8个tRNA基因由轻链编码;tRNA基因的A+T含量为87.25%,AT偏斜为0.009,GC偏斜为0.190。除了trnI缺受体臂、trnS1缺DHU臂、trnF缺TΨC臂和trnS2的 DHU臂仅形成一个简单的环外,其余18个tRNA基因可以形成常规的三叶草结构(图3)。在trnA,trnL1和trnS2的氨基酸臂、trnG和trnQ的DHU臂中共出现5处错配,错配碱基均为G-U。

图3 元宝枫刺胫广肩小蜂线粒体基因组中22个tRNA基因的预测二级结构Fig. 3 Predicted secondary structure of the 22 tRNA genes in the mitochondrial genome of Eurytoma acutibialis
元宝枫刺胫广肩小蜂线粒体基因组包含两个rRNA基因rrnS和rrnL,长度分别为763和1 299 bp,位于重链J,被trnQ和trnA分隔开。rRNA的基因全长2 062 bp,A+T含量为86.20%,T, C, A和G含量分别为43.10%, 4.90%, 43.10%和9.00%,比整个线粒体基因组A+T碱基含量要高,AT偏斜为0.007,GC偏斜为0.294。控制区长度为762 bp,T, C, A和G 的碱基含量分别为29.67%, 7.73%, 40.53%和19.57%,A+T碱基含量为70.20%,AT偏斜为0.155,GC偏斜为0.313。
2.4 系统进化
贝叶斯法分析的结果显示,支持小蜂总科(Chalcidoidea)各个类群之间的分类关系为(((((柄翅小蜂科(Mymaridae)+((((榕小蜂科(Agaonidae)+小蜂科(Chalcididae))+(((金小蜂科(Pteromalidae)+((蚜小蜂科(Aphelinidae)+长尾小蜂科(Torymidae)+赤眼蜂科(Trichogrammtidae))+(姬小蜂科(Eulophidae)+旋小蜂科(Eupelmidae)+广肩小蜂科(Eurytomidae)+跳小蜂科(Encyrtidae))))))(图4: A)。最大似然法支持的小蜂总科各个类群之间的分类关系为((((柄翅小蜂科(Mymaridae)+小蜂科(Chalcididae))+榕小蜂科(Agaonidae)))+(姬小蜂科(Eulophidae)+金小蜂科(Pteromalidae))))+(跳小蜂科(Encyrtidae)+广肩小蜂科(Eurytomidae)+旋小蜂科(Eupelmidae)+蚜小蜂科(Aphelinidae)+长尾小蜂科(Torymidae)+赤眼蜂科(Trichogrammtidae)))))(图4: B)。综合比较两种方法,虽然贝叶斯法中个别分支节点上的支持率较低,并且与最大似然法展示的拓扑结构存在差异,但是两种系统发育树均支持小蜂总科各亚科的单系性。根据两种方法构建的系统发育树中,跳小蜂科、广肩小蜂科和旋小蜂科在两种方法中显示为关系最近的姐妹群。无论是贝叶斯法还是最大似然法构建的系统发育树,元宝枫刺胫广肩小蜂均被归类于广肩小蜂属Eurytoma,与形态学分类结果一致,并显示与Eurytomasp. ZJUH 2016013之间存在较为密切的亲缘关系。
2.5 基因重排
研究结果表明,膜翅目小蜂总科普遍存在基因重排现象,主要发生在tRNA基因和rRNA基因,而13个PCGs排序在各亚科内相对稳定。与祖先昆虫相比,外群和柄翅小蜂科的13个PCGs排序与元宝枫刺胫广肩小蜂的一致,而其他小蜂总科昆虫的基因排序则发生了改变,尤其在长尾小蜂科、赤眼蜂科、跳小蜂科和广肩小蜂科中存在多种基因重排方式。本研究鉴定到13个PCGs的排列方式存在8种基因排列类型,其中cox1-cox2-atp8-atp6-cox3-nad3-nad2-nad1-cytb-nad6-nad4L-nad4-nad5,cox1-cox2-atp8-atp6-cox3-nad2-nad3-nad1-cytb-nad6-nad4L-nad4-nad5和cox1-nad5-nad4-nad4L-nad6-cytb-nad1-nad2-nad3-cox3-atp6-atp8-cox2这3种排列方式在多个物种中共享,而广肩小蜂利用其中两种排列方式。
与祖先昆虫相比,元宝枫刺胫广肩小蜂的27个基因发生了重排,其中cox1-L2-cox2-K-D-atp8-xx-cox3-G-nad3排列顺序和方向均一致,而nad2-xx-rrnS-xx-rrnL-L1-nad1-xxx-cytb-nad6-P-T-nad4L-nad4-H-nad5发生了原位倒置。此外,16个tRNA基因也发生了基因重排,其中trnC和trnM发生基因移位,trnH,trnT,trnP和trnL1发生倒置,trnS2发生异位倒置,trnF-trnE,trnW-trnS1-trnY-trnI-trnV和trnQ-trnA发生基因洗牌。rrnS和rrnL由A和Q隔开,这一现象也存在于柄翅卵小蜂科、榕小蜂科、蚜小蜂科、长尾小蜂科、旋小蜂科和跳小蜂科中。
3 讨论
根据魏书军(2009)的研究,大多数膜翅目昆虫的线粒体基因组表现出明显的AT偏斜现象。在膜翅目小蜂总科昆虫中,A+T含量一般在75%~90%之间,其中跳小蜂科基因组具有最低的A+T含量(78.40%),而柄翅小蜂科具有最高的A+T含量(87.50%)。作为广肩小蜂的成员,已知的3个广肩小蜂线粒体基因组A+T含量均超过了80%。例如,元宝枫刺胫广肩小蜂的A+T含量为82.00%(表1),由37个典型基因(13个PCGs、2个rRNA基因、22个tRNA基因)和一段控制区组成,与大部分膜翅目昆虫线粒体基因组的结果(Boore, 1999; Huangetal., 2016; Tangetal., 2021; Zhaoetal., 2021; Zhouetal., 2021)一致。这些基因,有27个位于J链,10个位于N链,这与许多物种的编码基因大多数位于J链,只有少数位于N链的情况(Boore, 1999)一致。关于小蜂总科的文献报道中并未详细说明基因的分布情况,目前仅在长尾小蜂科中的螳小蜂Podagrionsp.的研究中观察到类似的现象(Yangetal., 2019),造成这种现象的原因可能是物种进化过程中基因重排所导致的,具体原因有待进一步研究研讨。
元宝枫刺胫广肩小蜂有部分tRNA二级结构上存在缺陷,如trnS1缺失了 DHU臂,这与小蜂科、姬小蜂科、跳小蜂科、金小蜂科等的情况 (Yanetal., 2019; Zhang YCetal., 2020; Tangetal., 2021; Zhouetal., 2021)相似。trnS2的DHU臂仅形成小环,这与广肩小蜂E.sp. TJS 2016以及其他一些膜翅目昆虫的情况(Weietal., 2010; Suetal., 2016; Zhangetal., 2017; Xingetal., 2021)相似。两个rRNA基因由trnQ和trnA分隔,这一特征在Eurytomasp. TJS-2016和Eurytomasp. ZJUH_2016013中观察到,但在Sycophilasp. 2 JXW-2020中却不存在。Su等(2016)指出这一特点在膜翅目广肩小蜂为独特存在,然而,我们发现柄翅卵小蜂科、榕小蜂科、蚜小蜂科、长尾小蜂科、旋小蜂科和跳小蜂科也存在这种现象。
线粒体基因重排在缨翅目、蚧虫、虱目、膜翅目昆虫中普遍存在(Shaoetal., 2001; Dowtonetal., 2002; 魏书军, 2009; Weietal., 2010; Xiaoetal., 2011; 陈志腾和杜予州, 2016; Luetal., 2020; Tyagietal., 2020; 王建霞等, 2021; 邓鋆等, 2022; Yietal., 2022)。本研究通过对膜翅目小蜂总科、细蜂总科和姬蜂总科的线粒体基因组进行比较分析,发现小蜂总科昆虫线粒体基因重排速率高,特别是在tRNA和rRNA基因的排列上存在大量移位、原位和异位倒置。元宝枫刺胫广肩小蜂的线粒体基因组发生了重排,其中tRNA基因的两侧除trnV和trnS1外均有基因间隔,这与魏书军(2009)的报道一致,即发生重排的基因两侧一般存在基因间隔;PCGs在单系群内的排列相对保守,在所有分析的膜翅目中,atp8-atp6-cox3,nad4-nad4L和nad6-cytb保持不变,这些PCGs之间无tRNA基因,而小蜂总科得cox3和nad3之间发生了基因重排;但在膜翅目青蜂总科、蜜蜂总科、胡蜂总科、姬蜂总科、细蜂总科和旗腹蜂总科中atp8-atp6-cox3-trnG-nad3保持不变(魏书军, 2009),进一步说明了小蜂总科的进化速率更快。
通过贝叶斯法和最大似然法两种方法构建的系统发育树,结果均支持广肩小蜂科的单系性,并支持跳小蜂科、广肩小蜂科和旋小蜂科为姐妹群,与其他小蜂总科各亚科形成不同的姊妹群和并系群,这与先前报道的系统发育分析结果(Zhaoetal., 2021)一致。贝叶斯法支持柄翅小蜂科在小蜂总科的基部位置,但不支持Eurytoma亚属和Sycophila亚属的单系性;而最大似然法支持Eurytoma亚属和Sycophila亚属的单系性。无论是贝叶斯法还是最大似然法构建的系统发育树,元宝枫刺胫广肩小蜂均被归类于广肩小蜂属Eurytoma,与形态学分类结果一致,并表明其与Eurytomasp. ZJUH 2016013亲缘关系较近。本研究获得了元宝枫刺胫广肩小蜂线粒体基因组序列和结构组成等基础信息数据,丰富了广肩小蜂科基因组的生物信息学数据库,并分析了小蜂总科中基因重排现象,结果提示线粒体基因组可以作为研究类群分化的参考依据,但不能作为小蜂总科分类学的主要指标。
致谢元宝枫籽象Bradybatussp.由中国科学院动物研究所张润志研究员帮助鉴定,在此谨表谢意。
