CYP19A1对兔卵巢颗粒细胞增殖和凋亡的影响
2023-10-29杨安琪李嘉诚靳荣帅赵博昊吴信生
杨安琪,李嘉诚,宋 颖,陈 欣,靳荣帅,赵博昊,吴信生,陈 阳
(扬州大学动物科学与技术学院,扬州 225009)
颗粒细胞(granulosa cells,GCs)对卵母细胞的生长与成熟起着重要的调节作用,GCs参与维持成熟分裂的阻断和诱导排卵,调节卵母细胞成熟分裂[1-3]。在所有哺乳动物中,GCs的增殖率会随着卵泡大小的变化而改变[3]。在原始卵泡中,GCs增殖率低,但随着卵泡腔的生长和卵泡的增大,细胞由分裂逐步向分化转变,限制了正在分化的GCs增殖[3]。GCs的增殖和凋亡受到许多基因的控制。SMAD7基因抑制山羊卵巢颗粒细胞的增殖,促进其凋亡[4]。IGF1R基因表达下调,使AKT /mTOR信号通路下调,进而抑制卵巢GCs增殖及促进其凋亡[5]。抑制WNT5a基因的表达,使BAX/BCL-2的比值上调,从而调控Wnt/β-Catenin通路来抑制卵巢GCs的增殖及促进凋亡[6]。外源BMP6对鸡卵泡GCs体外增殖有显著影响[7]。在母兔卵巢卵泡发育过程中,哪些因子发挥了重要作用,还有待进一步挖掘。
芳香化酶(aromatase, CYP19)是细胞色素P450酶系中的一种,P450芳香化酶(cytochrome P450, family 19, subfamily A, polypeptide 1,CYP19A1)是芳香化酶的编码基因。研究发现,CYP19A1是猪卵巢GCs中的抗凋亡因子,也是猪卵巢GCs中雌二醇(E2)合成必需的[8]。牦牛卵丘卵母细胞复合体(COCs)成熟过程中CYP19A1上调内源性E2水平,使早期卵母细胞的发育水平得到提升[9]。GCs中CYP19A1 mRNA水平与E2水平呈正比例关系,在一定范围内,CYP19A1 mRNA表达量越高,分泌的E2越多,说明CYP19A1可以通过调控激素水平间接影响卵泡的发育,CYP19A1 mRNA表达量的变化与卵泡发育的数量密切相关[10]。而CYP19A1基因对兔卵巢GCs增殖和凋亡的调控功能尚不明确。
课题组前期通过转录组测序筛选了多个参与母兔繁殖周期调控的基因,其中关键差异表达基因CYP19A1在甾类激素生物合成、卵巢类固醇生成等信号通路中都有所富集,由此推测CYP19A1可能对母兔繁殖周期的调控也起着重要的作用[11-13]。本研究计划克隆CYP19A1基因,利用生物信息学预测分析其蛋白质结构和理化性能。进一步分离并鉴定兔卵巢GCs,构建pcDNA3.1-CYP19A1过表达载体和siRNA,分别转染卵巢GCs,以探究CYP19A1对GCs增殖和凋亡的影响,以及对类固醇激素合成相关基因表达的影响,为进一步了解母兔卵泡发育和繁殖周期调控的分子机制提供理论依据。
1 材料与方法
1.1 卵巢GCs的分离和培养
采集8月龄健康新西兰白兔母兔的卵巢,置于含有DMEM培养基的无菌培养皿中,刺破卵泡,使GCs释放到培养基中。将GCs混合液过滤、低速离心后弃上清液。加入2 mL红细胞裂解液,轻吹混匀后裂解5 min,再次放入离心机低速离心后弃去上清液。加入含有10%胎牛血清和1%双抗的DMEM-F12,制成细胞悬液后接种到6孔板中,在5% CO2,37 ℃的条件下培养。
1.2 卵巢GCs的鉴定
原代GCs培养24 h后,用4%多聚甲醛固定液在室温下固定30 min,再用0.3% TritonX-100处理60 min,每步完成后都需要用PBS洗3次。1% BSA室温封闭60 min,弃去封闭液后向其中加入一抗(anti-FSHR,1∶250),在4 ℃环境下培养过夜。过夜后用PBST漂洗3次,加入1∶2 000稀释过后的山羊抗兔IgG(FITC)二抗,在37 ℃孵育60 min。PBST漂洗3次,滴加DAPI,在室温下染色10 min后,用荧光显微镜观察显色,拍照保存。
1.3 pcDNA3.1-CYP19A1质粒构建
以pcDNA3.1(+)质粒作为载体,使用CE Design软件设计一步克隆引物,构建CYP19A1的真核表达载体。在上游引物中加入XbaⅠ酶切位点和上游载体末端同源序列,在下游引物中加入EcoR Ⅴ酶切位点和下游载体末端同源序列(表1)。使用XbaⅠ酶和EcoR Ⅴ酶对pcDNA3.1(+)载体进行双酶切,琼脂糖凝胶回收。使用重组酶Exane®Ⅱ把CYP19A1 CDS扩增产物和双酶切后的pcDNA3.1(+)载体连接。转化后,测序,进行无内毒素质粒小提。

表1 兔CYP11A1克隆的引物序列
1.4 siRNA的设计与合成
根据兔CYP19A1序列设计出siRNA序列(表2),用于兔卵巢GCs的转染。

表2 兔CYP19A1 siRNA序列信息
1.5 细胞转染
在24孔板中用DMEM-F12培养基对卵巢GCs进行培养,当细胞长到70%左右时利用LipofectamineTM2000进行转染,每组3个重复。8 h后弃去培养上清,在每个孔中加入500 μL DMEM-F12培养基,继续在培养箱中培养。48 h后,提取并保存细胞总RNA,用于后续试验。
1.6 细胞增殖检测
将转染后的卵巢GCs接种到96孔板中,随后每24 h检测一次细胞的增殖水平。每次测定先加入10 μL CCK-8 Solution,混合均匀后放入培养箱共同培育,2 h后使用酶标仪检测450 nm处的吸光值,并做好相应记录。
1.7 细胞凋亡检测
用Annexin V-FITC-PI试剂盒检测细胞的凋亡水平。转染后的GCs培养48 h后用胰酶消化并收集,1 000 r·min-14 ℃离心5 min,弃去上清液;使用预冷过的PBS洗涤细胞2次,1 000 r·min-1、4 ℃离心5 min,弃上清液;加入100 μL Binding Buffer、5 μL Annexin V-FITC和5 μL PI Staining Solution,每加入一种试剂都要吹打均匀后再加入下一种试剂;全部加入并混匀后在避光、室温的环境下孵育10 min;加入400 μL Binding Buffer,轻轻混匀;染色完成后在1 h内用流式细胞仪对样品进行检测,并使用软件进行分析,计算出细胞凋亡率。
1.8 细胞总RNA提取
收获1×107~5×107GCs,加入1 mL Trizol,反复抽取混匀后室温静置5 min。加入200 μL 氯仿,剧烈振荡25 s左右,静置2 min。12 000 g 4 ℃离心15 min,小心吸取上清,加入500 μL异丙醇,摇匀后在室温条件下静置10 min。12 000 g 4 ℃离心10 min,弃去上清液,加入75%乙醇(冰冷)1 mL,振摇,充分洗涤沉淀。12 000 g 4 ℃离心5 min,弃上清,短暂离心,小心吸取弃上清。加入适量(20 μL)无酶水溶解,测定RNA浓度及纯度,置-80 ℃保存。
1.9 荧光定量PCR
用NCBI Blast(https:∥www.ncbi.nlm.nih.gov/tools/primer-blast/)网页设计荧光定量引物,检测与CYP19A1以及类固醇激素合成相关基因CYP1B1、CYP1A1、IGF1R、BMP6、CYP17A1的表达量,内参基因选用GAPDH,引物详细信息见表3。反应体系20 μL:2×ChamQ SYBR qPCR Master Mix 10 μL,上游引物0.4 μL,下游引物0.4 μL,50×ROX Reference Dye2 0.4 μL,cDNA 1 μL,ddH2O 7.8 μL。反应程序为预变性:95 ℃ 30 s;循环反应:95 ℃ 10 s、60 ℃ 30 s(循环40次);熔解曲线采集:95 ℃ 15 s、60 ℃ 60 s、95 ℃ 15 s。

表3 荧光定量引物信息
1.10 数据统计与处理
采用Excel对数据进行统计,用2-ΔΔct法处理荧光定量结果,用SPSS 25.0成对T检验对数据进行显著性分析,最后使用GraphPad prism 8绘图。*为差异显著(P<0.05),**为差异极显著(P<0.01)。
2 结 果
2.1 CYP19A1基因克隆和过表达载体的构建
提取GCs总RNA,以cDNA为模板,扩增出CYP19A1基因1 500 bp左右的片段(图1A)。经比对,CYP19A1基因序列全长1 512 bp,序列同源性高达99%。进一步构建了pcDNA3.1-CYP19A1质粒,载体经XbaⅠ酶和EcoR Ⅴ双酶切后的载体片段与插入片段大小符合预期(图1B)。

A.CYP19A1扩增电泳图;B.双酶切电泳图A. CYP19A1 amplification electrophoresis figure; B. Double enzyme digestion electrophoresis figure图1 CYP19A1扩增及双酶切电泳结果图Fig.1 Plots of CYP19A1 amplification and double enzyme digestion electrophoresis
2.2 CYP19A1氨基酸序列的预测分析
经ExPASy ProtParam预测,CYP19A1为稳定的亲水性蛋白(图2A),其蛋白分子式为C2599H4114N672O711S39,不稳定系数为37.24,亲水性平均值为0.088。使用SOPMA和SWISS-MODEL预测发现,CYP19A1二级结构主要由α螺旋和无规卷曲构成(图2B),三级结构预测表现为弯曲螺旋状(图2C)。STRING预测其互作蛋白质主要有HSD17B1、HSD17B2、HSD17B3、CYP1A1、SULT1E1、SRD5A2和SRD5A1等蛋白(图2D)。
利用NetPhos预测,CYP11A1蛋白有31个潜在磷酸化位点(图2E)。经过PSORT Ⅱ Prediction亚细胞定位预测分析,发现CYP19A1蛋白主要存在于内质网和高尔基体上(图2F)。共编码503个氨基酸,其中亮氨酸(Leu)含量最多,占比10.1%,色氨酸(Trp)含量最少,所占比例为1.2%(表4)。CDD database预测发现,CYP19A1蛋白含有一个结构域,属于P450家族(图2H)。

表4 兔CYP19A1蛋白氨基酸组成
通过MEGA11软件将人(Homosapiens)、北美鼠兔(Ochotonaprinceps)、牛(Bostaurus)等9个物种的CYP19A1蛋白序列与兔进行同源性对比,发现兔与北美鼠兔聚类为一支(图2G)。
2.3 兔卵巢GCs分离培养和鉴定
卵巢GCs为贴壁细胞,细胞培养8~12 h后即可贴壁生长,形态为不规则的多边形或梭形。利用FSHR在GCs细胞膜特异性表达的特点,对分离的细胞进行免疫荧光染色。本试验中细胞膜呈绿色荧光(图略),证实获得的细胞为GCs。
2.4 CYP19A1基因过表达和干扰对类固醇激素合成相关基因的影响
在GCs中,对CYP19A1进行过表达和干扰,通过实时荧光定量检测IGF1R、CYP1B1、CYP1A1、BMP6、CYP17A1基因的表达量。发现CYP19A1过表达极显著提高了IGF1R、CYP1B1、CYP1A1的表达量,BMP6、CYP17A1表达量极显著降低(P<0.01)(图3A和3B)。与siRNA-NC相比,3个siRNA均抑制了CYP19A1的表达,以siRNA-2效果最显著(图3C)。进一步进行干扰试验,发现IGF1R、CYP1B1、CYP1A1基因的表达量均显著下降(P<0.05或P<0.01),BMP6和CYP17A1的表达量均极显著增加(P<0.01)(图3D)。结果说明CYP19A1可调控类固醇激素合成相关基因的表达。
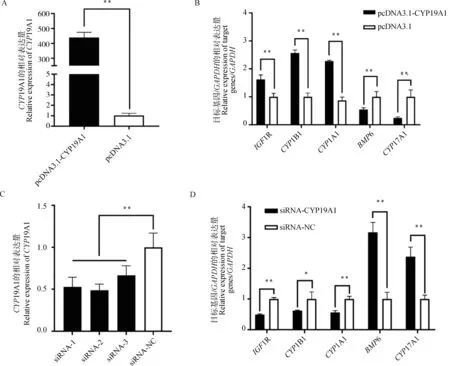
A.CYP19A1过表达效果分析;B.CYP19A1过表达对类固醇激素合成相关基因表达的影响;C.CYP19A1干扰效果分析;D.CYP19A1干扰对类固醇激素合成相关基因表达的影响。*.P<0.05;**.P<0.01,下同A.Analysis of CYP19A1 overexpression effect;B.Effects of CYP19A1 overexpression on the expression of genes related to steroid hormone synthesis;C.Analysis of interference effect of CYP19A1;D.Effects of CYP19A1 interference on the expression of genes related to steroid hormone synthesis. *. P<0.05; ** .P<0.01, the figure below is the same图3 过表达和干扰CYP19A1对类固醇激素合成相关基因表达的影响Fig.3 Effects of overexpression and interference with CYP19A1 on the expression of genes related to steroid hormone synthesis
2.5 CYP19A1基因过表达和干扰对卵巢GCs增殖和凋亡的影响
过表达和干扰CYP19A1基因,利用CCK8法和FITC/PI法检测GCs的增殖和凋亡情况。结果显示,过表达CYP19A1基因24 h后显著提高了GCs的增殖水平,相反,干扰CYP19A1基因24 h后显著降低了GCs的增殖水平(P<0.01)(图4A)。进一步过表达CYP19A1基因显著抑制了GCs的凋亡,反之干扰CYP19A1基因则显著加快了GCs的凋亡(P<0.01)(图4B)。由此可得CYP19A1基因可以促进GCs增殖,抑制GCs凋亡。

A.CYP19A1表达量变化对GCs增殖的影响;B.CYP19A1表达量变化对GCs凋亡的影响A. Effects of CYP19A1 expression changes on the proliferation of GCs; B. Effect of CYP19A1 expression changes on apoptosis of GCs图4 过表达和干扰CYP19A1对卵巢GCs增殖和凋亡的影响Fig.4 Effects of overexpression and interference with CYP19A1 on proliferation and apoptosis of ovarian GCs
3 讨 论
CYP19A1属于细胞色素P450超家族,在正常发育的卵泡GCs中表达量较高,是催化睾酮转化为雌二醇的关键酶[14-15]。雌激素能够促进有腔卵泡的快速发育和成熟,在卵泡发育过程中起重要作用,是调控哺乳动物进入发情期的主要激素[16-17]。课题组前期研究结果证实了CYP19A1参与母兔发情周期调控。本研究克隆了兔的CYP19A1基因,经生物信息学预测,发现CYP19A1蛋白为稳定亲水性蛋白,进化树分析确定其是一个十分保守的基因。
进一步为探究CYP19A1基因调控母兔发情周期的分子机制,构建了CYP19A1的过表达载体和干扰序列。经转染后发现,过表达CYP19A1可使IGF1R、CYP1B1和CYP1A1的表达量增加,BMP6和CYP17A1的表达量减少,经干扰后,上述基因表达量的变化趋势与过表达结果均相反,符合预期。已知雌激素是影响动物发情的重要因素[18],CYP1B1、CYP1A1与CYP19A1一样,同为细胞色素P450家族成员,可影响雌激素的氧化代谢[19-20]。CYP1A1有4个位点具有多态性,其基因多态性可影响酶的表达量和活性,从而调控雌激素的体内代谢[21-22]。在发情前期CYP1B1的表达水平明显上升[23]。推测CYP19A1与CYP1A1、CYP1B1可能存在协同作用,通过共同调控体内雌激素水平,间接影响动物的发情周期。雌激素受体(ER)是介导雌激素发挥生物学效应的重要受体,与CYP19A1的表达量呈正相关,ER与IGF1受体(IGF1R)的表达量也呈正相关,由此推测CYP19A1能够通过影响ER的表达进而调控IGF1R的表达量[24-25]。CYP17A1为雄激素合成限速酶,其不仅能够调控雄激素的合成,也有调控GCs合成孕酮的功能,影响着卵泡生长发育直至排卵的全过程[26-27]。CYP19A1的主要功能是促进雌激素合成,推断CYP19A1是通过降低CYP17A1的表达量来抑制雄激素和孕酮的合成,从而促进雌激素的合成。BMP6属于BMPs(骨形态发生蛋白)家族的一员,BMP属于转化生长因子TGF-β(transforming growth factor-β)超家族[28-29],BMPs家族在卵泡发生的各阶段均发挥着重要作用[30-32]。在哺乳动物中,BMP-15、BMP7在原始卵泡的激活及卵泡的发育过程中发挥重要作用,而BMP6的表达缺失对优势卵泡的选择具有较大影响[33-35]。BMP6和BMP7的共同作用会降低GCs的E2合成量[36]。BMP6还可以促进母鸡前卵泡颗粒细胞中的促卵泡生成素(FSH)受体mRNA的表达[37]。此外,BMP6能够通过上调限速酶CYP19A1的表达来促进E2和P4的分泌[38]。本研究中,CYP19A1过表达后,极显著抑制了BMP6表达,推测可能存在负反馈机制,但仍需要进一步探索。综上,提示CYP19A1可能通过调控类固醇激素合成相关基因的表达,参与母兔卵泡的发育。
本研究发现,过表达CYP19A1可促进GCs增殖、抑制GCs凋亡,而干扰其表达则会抑制GCs增殖,促进GCs凋亡。有研究证实,CYP19A1可催化雄激素转化为雌激素,而雌激素中E2最具生物活性,可促进卵巢GCs增殖和卵泡分化[39-40]。为后期深入探究CYP19A1调控GCs增殖和凋亡的途径提供了思路。综上所述,CYP19A1在兔卵泡发育过程中发挥着至关重要的作用,并可能通过调控类固醇激素合成相关基因的表达,影响GCs增殖和凋亡,参与母兔的发情周期。
4 结 论
CYP19A1蛋白是一种稳定的亲水性蛋白,与其它物种同源性较高。CYP19A1基因可以促进GCs增殖,抑制GCs凋亡,调控类固醇激素合成相关基因的表达,进而参与母兔卵巢卵泡发育。
