倒春寒对开花期小麦根际土壤真菌群落的影响*
2023-10-25王捧娜代雯慈于敏翁颖黄谨卫吴宇蔡洪梅郭健鹏高赛郑宝强李金才陈翔
王捧娜,代雯慈,于敏,翁颖,黄谨卫,吴宇,蔡洪梅,郭健鹏,高赛,郑宝强,李金才,2**,陈翔**
(1.安徽农业大学农学院/农业部华东地区作物栽培科学观测站 合肥 230036;2.江苏省现代作物生产协同创新中心 南京 210095)
联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)报告指出,21世纪末全球地表均温增长可能超过1.5 ℃[1]。随着全球气候变暖的加剧,突发性、难预测性和强破坏性极端气象灾害事件的发生频率增加,导致农业生产的不稳定性增加。小麦(Triticum aestivumL.)是全球20 亿以上人口的主要口粮,能为人类提供约20%的蛋白质和热量[2]。近年来极端低温气候灾害事件的频发重发已严重影响中国、美国和澳大利亚等小麦主产国小麦的稳产丰产与优质,对全球粮食安全造成了极大的威胁[3]。1980-2020 年中国小麦主产区发生近20 次大规模低温灾害事件,其中尤以黄淮麦区的倒春寒(late spring coldness in wheat,LSCW)灾害危害最为严重[4]。小麦倒春寒主要指拔节至孕穗期间遭遇突然降温天气,造成幼穗受伤或死亡,部分小穗不结实甚至全穗不结实,从而导致小麦减产的一种农业气象灾害[5]。黄淮麦区在2009 年、2013年、2015 年、2018 年和2020 年均有倒春寒灾害发生,年际发生频率达到了40%,灾害严重年份小麦减产高达50%以上[4,6]且导致籽粒品质劣化[7]。因此,倒春寒灾害已成为限制我国小麦生产高产高质高效发展的重要因素。
前人关于小麦倒春寒灾害的研究集中于对叶片(源)[8-11]、茎秆(流)[12]、穗部(库)[13-15]等地上部器官的影响。刘绿洲等[10]研究表明倒春寒会破坏小麦功能叶的细胞结构,使叶肉细胞内叶绿体面积和数目减小、基粒片层厚度和数量下降,进而导致其光合性能下降。柯媛媛等[16]研究表明倒春寒会降低小麦干物质的积累量、花前干物质的转运量、花前贮藏干物质对籽粒的贡献率,减少光合同化产物向穗部的转运与分配,从而导致小麦减产。Zhang 等[15]研究进一步指出倒春寒胁迫后小麦下部小穗的灾损要大于上部和中部小穗。Jiang 等[17]通过转录组分析研究表明小麦穗部在4 ℃和-4 ℃倒春寒下差异表达基因大于4000 个,其中蔗糖合成酶基因表达的上调可能对小麦的抗倒春寒能力起着重要作用。但目前鲜有报道研究倒春寒对小麦地下部根系及根际微生物群落结构和多样性的影响。作物根际是土壤微生物活跃的热点区域[18],参与作物的发育、营养吸收和胁迫响应等多个过程,对其生长及抗逆性十分重要[19]。土壤微生物对环境变化十分敏感,细微的外界环境变化都会改变微生物数量和群落结构[20]。因此阐明作物根际土壤微生物群落对环境变化的响应规律有助于增强其在提高作物抗逆能力方面的潜在功能,然而倒春寒灾害下小麦根际土壤真菌群落结构和功能变化尚不清楚。本研究选用抗倒春寒性强弱不同的两个小麦品种于幼穗分化的药隔期人工模拟倒春寒,利用Illumina 测序技术对小麦根际真菌群落特征进行分析,旨在探究倒春寒灾害对小麦根际土壤真菌群落结构和多样性的影响,为深入了解小麦根际真菌群落对倒春寒的响应提供理论支撑,从而丰富和完善小麦倒春寒的致灾机理研究。
1 材料与方法
1.1 试验地概况
试验于2021 年11 月4 日-2022 年5 月18 日在安徽农业大学农萃园(31°52′9.86″N,117°14′50.91″E,海拔21.3 m)进行,该地区属于北亚热带湿润季风气候,年均日照时数2100 h,年均气温15 ℃,年均降水量875 mm。该区域倒春寒多发生在每年3 月中下旬至4 月上旬,此时小麦正处于拔节——孕穗期,其幼穗正处于分化的关键时期,组织幼嫩、含水量较高,对低温十分敏感,且以幼穗分化的药隔形成期为低温敏感期[10-11,16]。试验土壤为黄棕壤土,取自地表0~20 cm,土壤pH为6.5,有机质16.3 g·kg-1、全氮1.3 g·kg-1、速效氮112.2 mg·kg-1、速效磷23.0 mg·kg-1、速效钾161.6 mg·kg-1。
1.2 试验设计
试验采用盆栽方式种植,土壤过筛后装入PVC桶中(直径25 cm,高度29 cm),每盆装土10 kg,播种前施纯氮0.12 g·kg-1、P2O50.10 g·kg-1和K2O 0.10 g·kg-1,拔节期追施纯氮0.04 g·kg-1(土)。在前期研究的基础上[21],选取抗倒春寒性弱的‘新麦26’(XM)和抗倒春寒性强的‘烟农19’(YN)两小麦品种为供试材料,每个处理6 盆,每盆播种籽粒饱满均匀且无病虫害的小麦种子18 粒,播种深度为3 cm,三叶期(2021年12 月9 日)间苗,每盆留9 株长势基本一致的幼苗。
于小麦幼穗分化的药隔形成期(第3 节间伸长、倒二叶露出)利用智能超低温光照培养箱(DWGZ-1008B3 型号,合肥右科仪器设备有限公司)模拟大田倒春寒灾害的温变规律进行低温胁迫处理,以10 ℃作为对照处理(CK);在前期研究基础上[15]设置2 ℃(T1)、-2 ℃ (T2)两个胁迫温度,于2022 年3 月17日1:00-5:00 低温处理4 h[16](培养箱内光强设置为0 Lux,相对湿度为75%)。处理结束后将盆栽移出培养箱,搬至大田生长直至成熟收获。其他田间管理措施同一般高产田。
1.3 取样方法
小麦开花期(2022 年4 月8 日)分别采集各处理小麦植株和根际土壤样本。将小麦植株分为地上部和根系两部分,于105 ℃烘箱中杀青30 min 后75 ℃烘干至恒重,测定其干物质重并计算根冠比。根冠比=根系干物质重/地上部干物质重。根际土壤样本采用多点混合样本采集方法,以盆为单位分别小心地从盆中取出植株,去除根际附着不紧密的土壤[19,22],然后用无菌刷收集附着紧密的土壤,混合每盆中植株根际土壤,每处理均获取6 个生物学重复,封入无菌袋,置于冰盒带回实验室,保存于-80 ℃冰箱备用。各处理根际土壤样本分别表示为XM_CK、XM_T1、XM_T2、YN_CK、YN_T1、YN_T2,由上海欧易生物医学科技有限公司进行测序。
1.4 DNA 提取与高通量测序
采用DNA 抽提试剂盒(MagPure Soil DNA LQ Kit)对土壤样本的基因组DNA 进行提取,之后利用琼脂糖凝胶电泳检测DNA 的纯度和浓度。真菌ITS 区使用前端ITS1F 引物5′-CTTGGTCATTTAGAGGAAGTAA-3′和后端ITS2R 引物5′-GCTGCGTTCTTCATCGATGC-3′[23]进行PCR 扩增。扩增程序为94 ℃预变性 5 min,33 个循环(94 ℃变性30 s,56 ℃退火 30 s,72 ℃ 延伸20 s),最后72 ℃5 min。扩增体系为30 μL,15 μL 2×Gflex PCR 缓冲液,正、反向引物(5 pmol∙μL-1)各1 μL,0.6 μL (1.25 U∙μL-1) Tks-Gflex DNA 聚合酶;50 ng DNA 模板,其他用双蒸水补齐。扩增产物使用Illumina NovaSeq 6000测序平台进行测序。
1.5 生物信息学分析
原始数据首先使用cutadapt 软件去掉引物序列,然后在QIIME2 (2020.11)[24]使用DADA2[25]进行拼接、质控、拼接合并生成ASV (Amplicon Sequence Variant,扩增子序列变异)。使用QIIME 2 软件包挑选出各个ASV 的代表序列后,并使用q2-feature-classifier 软件默认参数进行分析将所有代表序列与Unite(ver 8)数据库进行比对注释。
通过计算Chao1、Shannon、ACE、Simpson 指数表征小麦根际土壤微生物的多样性和丰富度,进行样本内物种多样性即Alpha 多样性和样本间Beta多样性分析表征真菌群落多样性变化;通过PCA 分析(principal components analysis,主成分分析)表征不同处理真菌群落的差异。计算公式如下[26-28]:
式中:SChao1为估计到的ASV 数量,Sobs为实际观测到的ASV 总数,n1为只含一条序列的ASV 的数量,n2为含两条序列的ASV 的数量,HShannon为香农指数,N为样品序列总数,DSimpson为辛普森指数,ni为第i个ASV 的序列数量。
20世纪六七十年代,文化大革命时期,全国大部分印刷企业处于停产歇业状态,中科印刷却依然在坚持生产。这一时期,中科院的主要科研成果——陈景润的哥德巴赫猜想、人工合成胰岛素等内容的排版印刷都在中科印刷完成。中科印刷在这一时期,发挥了举足轻重的作用。
1.6 数据分析
采用IBM SPSS Statistics 23 对数据进行方差分析,采用LSD 法比较各处理间的统计学差异(P<0.05)。用Microsoft Excel 2021 进行物种丰度柱状图绘制和图表制作(图表中数据均表示为平均值±标准误差),用Origin 2021 软件进行PCA 分析,采用欧易生物云分析平台进行Venn 图、LEfse 分析和FUNGuild 真菌功能预测。
2 结果与分析
2.1 倒春寒对开花期小麦生物量积累的影响
由表1 可见,倒春寒胁迫后两品种小麦开花期根干物质重和地上部干物质重较对照处理均下降,且降幅随倒春寒胁迫强度增加逐渐增大。与对照相比,T1 和T2 处理下,XM 开花期根干物质重和地上部干物质重分别显著下降7.03%、12.22%和21.66%、35.49%(P<0.05),YN 在T1 和T2 处理下根干物质重和地上部干物质重分别下降1.65%、4.13%和15.05%、18.31%。表明药隔期倒春寒会降低开花期小麦地上地下部生物量的积累且对抗倒春寒性弱的XM 影响更大。同时根冠比随倒春寒胁迫强度增加逐渐上升且与对照之间差异显著,XM 根冠比显著上升18.68%和36.08%,YN 根冠比显著上升15.79%和17.36%。
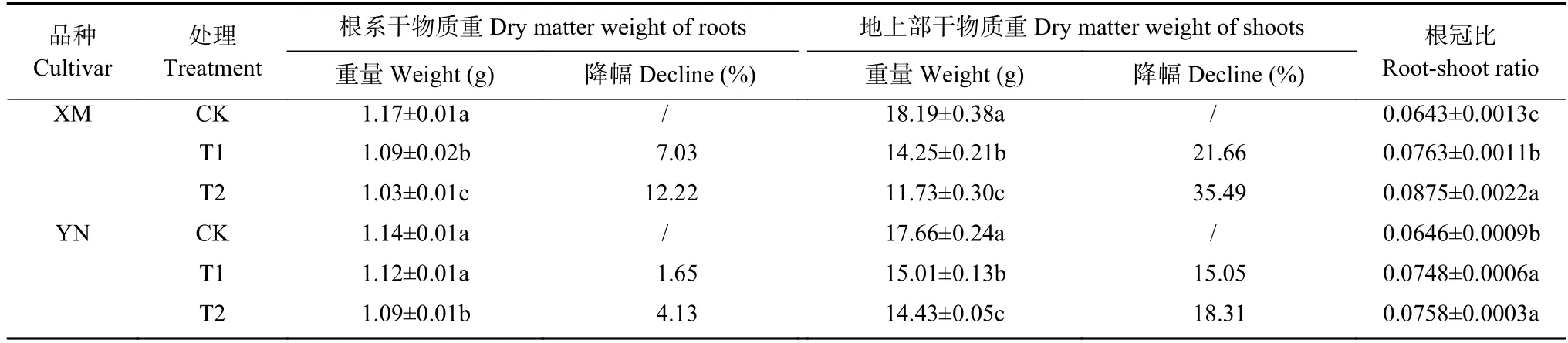
表1 不同倒春寒处理对不同品种小麦开花期干物质重及根冠比的影响Table 1 Effects of late spring coldness on root dry weight,aboveground dry weight and root-shoot ratio of different wheat cultivars at flowering stage
2.2 倒春寒对开花期小麦根际土壤真菌群落结构组成的影响
ASVs 的代表序列的分类学分析共得到8 门29纲76 目145 科256 属的土壤真菌。由图1A 可见,在门水平上,所有土样中共有8 个可注释到的门被检出,包含子囊菌门(Ascomycota)、接合菌门(Zygomycota)、担子菌门(Basidiomycota)、壶菌门(Chytridiomycota)、罗兹菌门(Rozellomycota)、丝足虫门(Cercozoa)、球囊菌门(Glomeromycota)和黄藻门(Anthophyta)。其中子囊菌门为第1 优势菌门,所有样品中的相对丰度为24.37%~42.32%;接合菌门为第2 优势菌门,在所有样品中的相对丰度为15.79%~40.06%。两品种中子囊菌门相对丰度均随胁迫程度增强而升高。而接合菌门的相对丰度在XM_T2 根际土壤中占比为15.79%,显著低于XM_CK、XM_T1、YN_CK和YN_T2 根际土壤接合菌门的相对丰度(P<0.05)。抗倒春寒性强的YN 根际土壤中担子菌门相对丰度在各不同胁迫程度下均高于抗倒春寒性弱的XM,但对于品种XM,2 ℃的倒春寒胁迫使其担子菌门相对丰度较CK 显著升高398.26% (P<0.05),而-2 ℃倒春寒胁迫较CK 显著升高61.57% (P<0.05)。壶菌门在XM_T2 根际土壤的相对丰度显著高于其他处理,为23.83% (P<0.05),在 XM_CK、XM_T1、YN_CK、YN_T1、YN_T2 根际土壤中的相对丰度分别为4.15%、5.32%、15.40%、3.24%、8.11%。综上,倒春寒胁迫下两小麦品种根际土壤子囊菌门相对丰度随胁迫程度增强而升高,XM_T2 中接合菌门相对丰度显著低于其他处理,而壶菌门相对丰度显著高于其他处理(P<0.05);YN 根际土壤中担子菌门在不同胁迫程度下相对丰度均高于抗倒春寒性弱的XM,两品种间真菌群落门水平上的相对丰度差异显著(P<0.05)且倒春寒胁迫对抗倒春寒性弱的XM 影响更大。
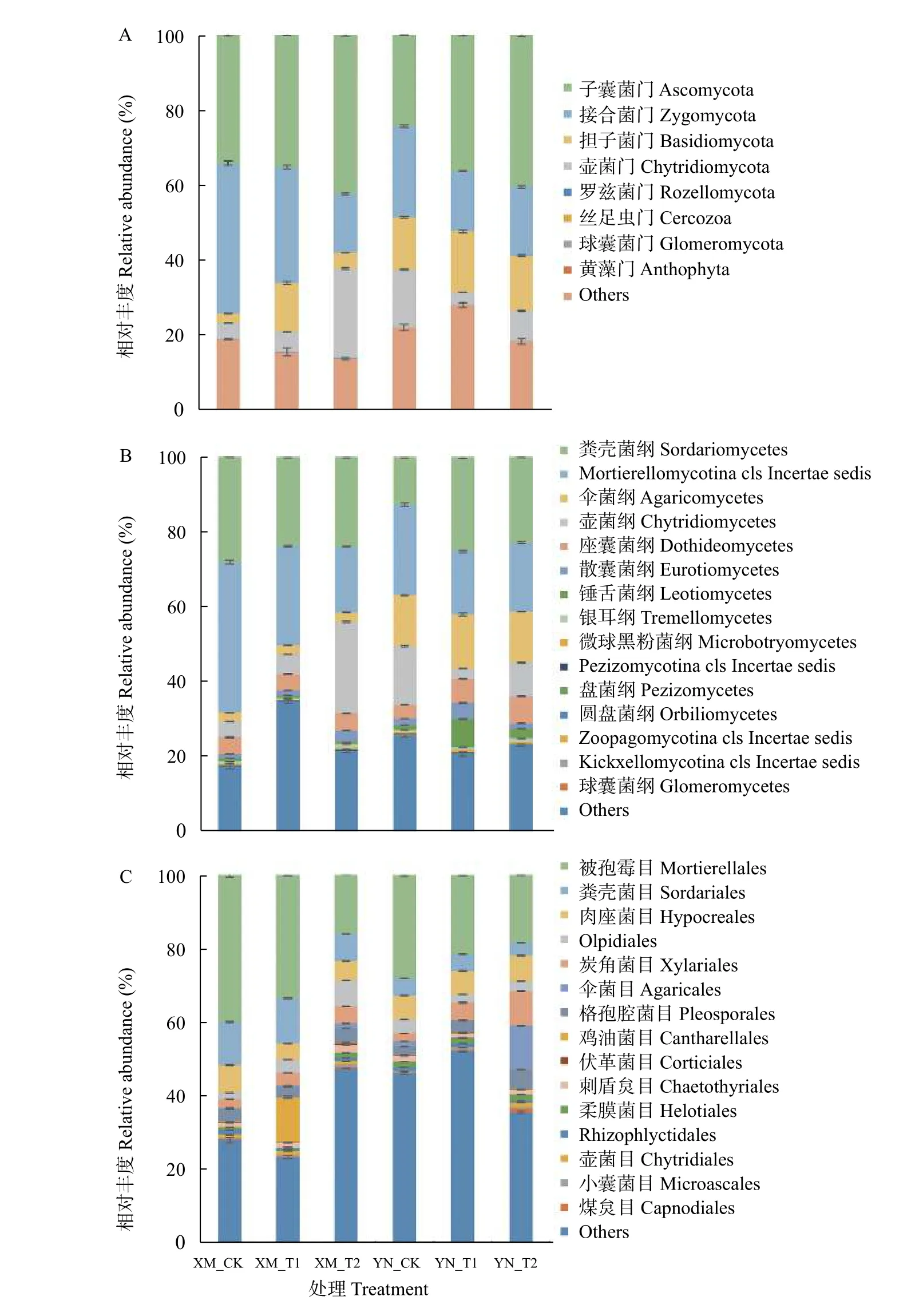
图1 不同倒春寒处理对不同品种小麦的根际土壤真菌各分类水平的组成和相对丰度的影响Fig.1 Effects of late spring coldness on the composition and relative abundance of fungi in rhizosphere soil of different wheat varieties
由图1B 可见,在纲分类水平上,排名前15 的分别为粪壳菌纲(Sordariomycetes)、Mortierellomycotina cls Incertae sedis、伞菌纲(Agaricomycetes)、壶菌纲(Chytridiomycetes)、座囊菌纲(Dothideomycetes)、散囊菌纲(Eurotiomycetes)、锤舌菌纲(Leotiomycetes)、银耳纲(Tremellomycetes)、微球黑粉菌纲(Microbotryomycetes)、Pezizomycotina cls Incertae sedis、盘菌纲(Pezizomycetes)、圆盘菌纲(Orbiliomycetes)、Zoopagomycotina cls Incertae sedis、Kickxellomycotina cls Incertae sedis、球囊菌纲(Glomeromycetes)。壶菌纲在XM_T2 根际土壤中的相对丰度显著高于其他处理,为24.27% (P<0.05)。此外,伞菌纲在YN 根际土壤中的相对丰度显著高于XM(P<0.05)。综上,T2 胁迫使得XM 中壶菌纲相对丰度显著增高(P<0.05),两小麦品种间根际土壤伞菌纲相对丰度表现为YN 显著高于XM。
由图1C 可见,在目水平上,排名前15 的分别为被孢霉目(Mortierellales)、粪壳菌目(Sordariales)、肉座菌目(Hypocreales)、Olpidiales、炭角菌目(Xylariales)、伞菌目(Agaricales)、格孢腔菌目(Pleosporales)、鸡油菌目(Cantharellales)、伏革菌目(Corticiales)、刺盾炱目(Chaetothyriales)、柔膜菌目(Helotiales)、Rhizophlyctidales、壶菌目(Chytridiales)、小囊菌目(Microascales)、煤炱目(Capnodiales)。不同程度倒春寒胁迫下,两品种小麦根际土壤中被孢霉目的相对丰度均随胁迫强度的增加呈下降趋势。对于抗倒春寒性较弱的XM,T1 根际土壤中被孢霉目的相对丰度相对于CK 下降16.06%,T2 相对于CK下降60.03%;而对于抗倒春寒性较强的YN,T1 和T2 较CK 分别降低23.03%和34.36%。粪壳菌目的相对丰度在两品种间差异显著(P<0.05),且表现为XM 根际土壤大于YN。综上,倒春寒胁迫下两品种小麦根际土壤中被孢霉目的相对丰度均随胁迫强度的增加而下降,且XM 下降幅度更大;不同胁迫程度下XM 的粪壳菌目相对丰度显著高于YN (P<0.05)。
由图1D 可见,在科水平上,排名前15 的分别为被孢霉科(Mortierellaceae)、赤壳科(Nectriaceae)、Xylariales fam Incertae sedis、毛球壳科(Lasiosphaeriaceae)、毛壳菌科 (Chaetomiaceae)、球盖菇科(Strophariaceae)、角担菌科(Ceratobasidiaceae)、伏革菌科(Corticiaceae)、晶杯菌科(Hyaloscyphaceae)、Rhizophlyctidaceae、黑盘孢科(Amphisphaeriaceae)、格孢腔菌科(Pleosporaceae)、鬼伞科(Psathyrellaceae)、Cantharellales fam Incertae sedis、壶菌科(Chytridiaceae)。6 个处理根际土壤中共有且所占比例较高的菌群为被孢霉科,属于接合菌门,其在 XM_CK、XM_T1、XM_T2、YN_CK、YN_T1 和YN_T2 中的占比分别为39.95%、31.22%、18.74%、30.35%、23.94%、18.27%。XM 和YN 的T1、T2分别较其CK 下降21.86%、53.09%和21.14%、39.82%。综上,倒春寒胁迫下两品种根际土壤中被孢霉科真菌均随胁迫程度增强而显著下降(P<0.05),且XM 下降幅度更大。
由图1E 可见,属分类水平上,相对丰度排名前15 的菌属分别为被孢霉属(Mortierella)、微结节菌属(Microdochium)、田头菇属(Agrocybe)、腐质霉属(Humicola)、赤霉属(Gibberella)、尾孢属(Cercospora)、Laetisaria、根囊壶菌 属(Rhizophlyctis)、Echria、毛壳菌属(Chaetomium)、短毛盘菌 属(Psilachnum)、刀菌属(Fusarium)、Limonomyces、顶多毛孢属(Bartalinia)、亡革菌属(Thanatephorus)。各处理根际土壤共有且所占比例较高的菌属为被孢霉属,其在两品种中均随胁迫程度增强而降低。XM_T1、XM_T2 和YN_T1、YN_T2 分别较各自的CK 下降20.85%、54.70%和12.58%、26.89%。综上,倒春寒胁迫下两品种根际土壤中被孢霉属真菌均随胁迫程度增强显著下降(P<0.05),且XM 下降幅度更大。
为观察不同样本之间真菌群落的相似性,通过主成分分析(PCA)发现,第1 主成分(PC1)和第2 主成分(PC2)分别可以解释所有变量的6.5%和5.5%。进一步分析表明,XM_T2 与其他处理距离较远,群落组成结构差异较大(图2)。
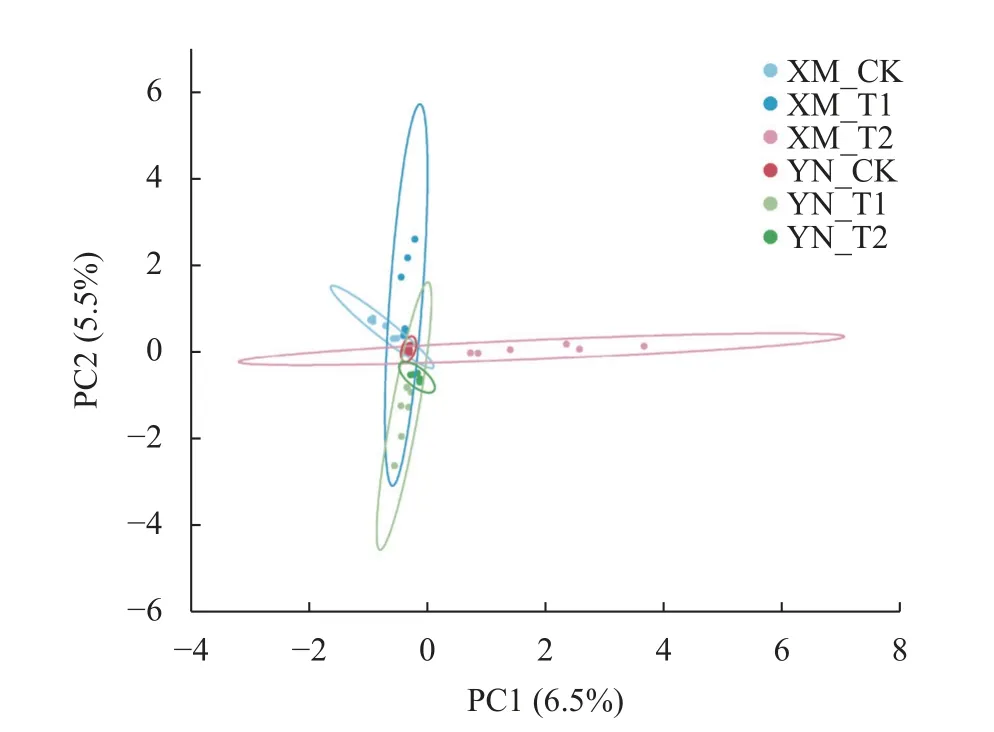
图2 不同倒春寒处理下不同品种小麦的根际土壤真菌PCA 分析Fig.2 Principal Component Analysis of rhizosphere soil fungi of different wheat cultivars under late spring coldness
通过Illumina 测序技术对两品种小麦不同倒春寒胁迫下的根际土壤样品进行测序,得到各样本ASV 分析不同处理之间共有、特有的 ASVs 个数如图3 所示。通过维恩图分析(图3),XM 各处理根际土壤样本共有82 个ASVs,单独存在于CK、T1 中的ASVs 数分别为83、90 个,而T2 处理样本中为114 个,高于CK 和T1,表明对于抗倒春寒性较弱的XM,-2 ℃倒春寒胁迫对根际土壤真菌群落结构有较大影响。YN 各处理根际土壤样本共有71 个ASVs,单独存在于CK、T2 中的 ASVs 数分别为 80、75 个,而T1 处理样本中为102 个,高于CK 和T2。表明倒春寒胁迫增加了XM 和YN 的根际土壤真菌群落组成。
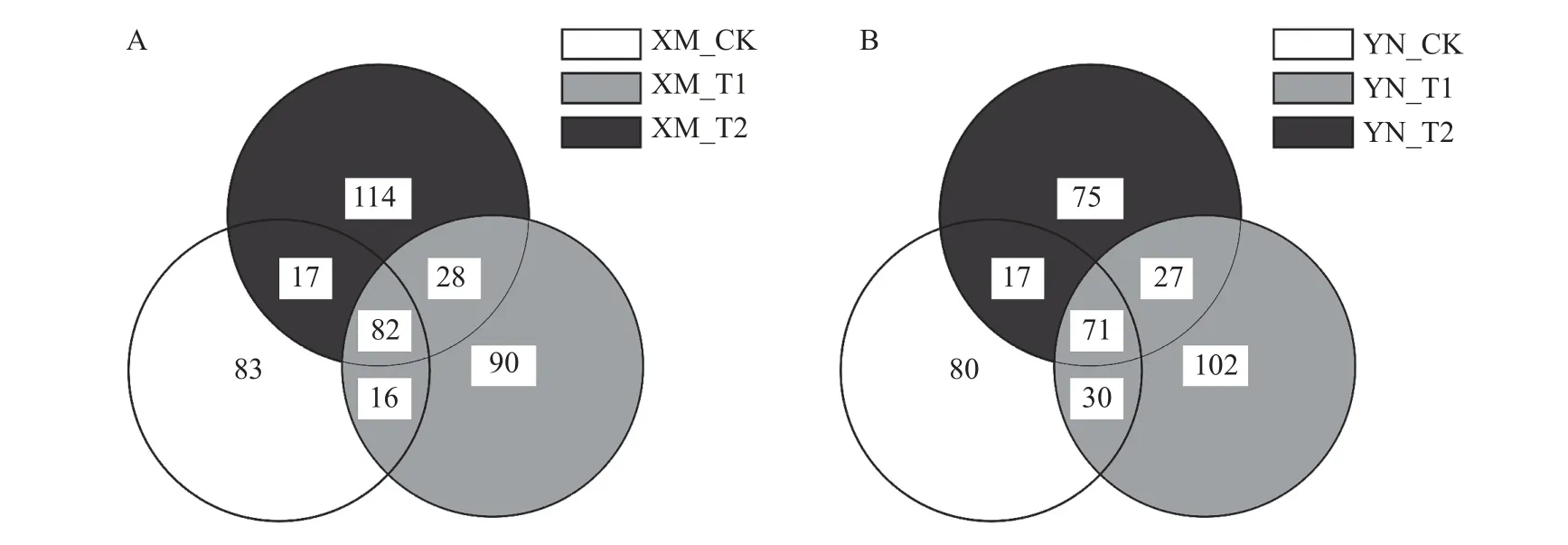
图3 不同倒春寒处理下不同品种小麦的根际土壤真菌 Venn 图Fig.3 Venn diagram of rhizosphere soil fungi of different wheat cultivars under late spring coldness
2.3 倒春寒对开花期小麦根际土壤真菌群落多样性的影响
由表2 可知温度显著影响了真菌多样性(P<0.05),品种、品种×温度的互作效应对Chao1 指数和ACE指数影响极显著(P<0.01),品种×温度的互作效应对Shannon 指数影响显著(P<0.05)。且对于XM 各项指数均随胁迫强度增加先降低后升高。
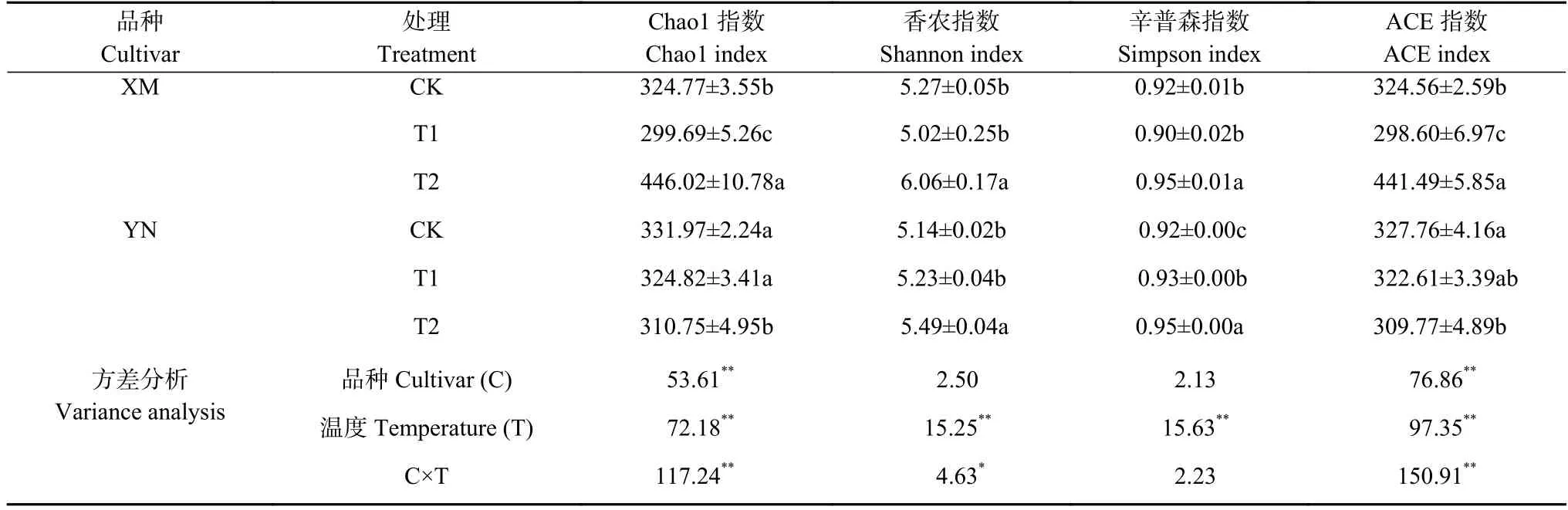
表2 不同倒春寒处理下不同品种小麦的根际土壤真菌α 多样性指数Table 2 Rhizosphere soil fungal Alpha diversity indexes of different wheat cultivars under late spring coldness
XM 各处理间反映群落丰富度的Chao 1 和ACE指数都具有显著性差异(P<0.05),反映群落多样性的Shannon、Simpson 指数在T1 与CK 间差异不显著,而T2 与CK 和T1 差异均显著(P<0.05)。表明XM_T2具有较高的物种丰富度和均匀度。YN 的T2 处理土壤真菌 ACE 指数、Chao1 指数低于其余两处理且与CK 具有显著性差异(P<0.05),Shannon 指数和Simpson 指数显著高于CK 和T1 处理。综上-2 ℃倒春寒胁迫对小麦根际真菌群落丰富度和均匀度影响更明显。
XM为抗倒春寒性弱小麦品种‘新麦26’,YN为抗倒春寒性强小麦品种‘烟农19’。CK、T1和T2分别为小麦幼穗分化的药隔形成期进行10 ℃、2 ℃和-2 ℃处理。同一品种同列不同小写字母表示处理间差异显著(P<0.05)。*和**分别表示在P<0.05和P<0.01水平差异显著。XM and YN are late spring coldness-sensitive and -tolerable wheat culviars,respectively.CK,T1 and T2 are treatments of 10 ℃,2 ℃ and -2 ℃ at anther differentiation period。Different lowercase letters in the same column of the same variety indicate significant differences atP<0.05.*and **indicate significant atP<0.05 andP<0.01 levels,respectively.
2.4 倒春寒对开花期小麦根际土壤真菌的LEfSe 物种差异分析
LEfSe 采用线性判别分析(LDA)来估算每个组分(物种)丰度对差异效果影响的大小。由图4A 可见,XM_CK 富集更多的 Hypocreales fam Incertae sedis、赤壳科、毛壳菌科、被孢霉科、肉座菌目、被孢霉目、Mortierellomycotina cls Incertae sedis,在XM_T2 中富集更多的散囊菌纲、壶菌纲、小囊菌目、伏革菌目、伞菌目、鬼伞科、伏革菌科和Olpidiales。
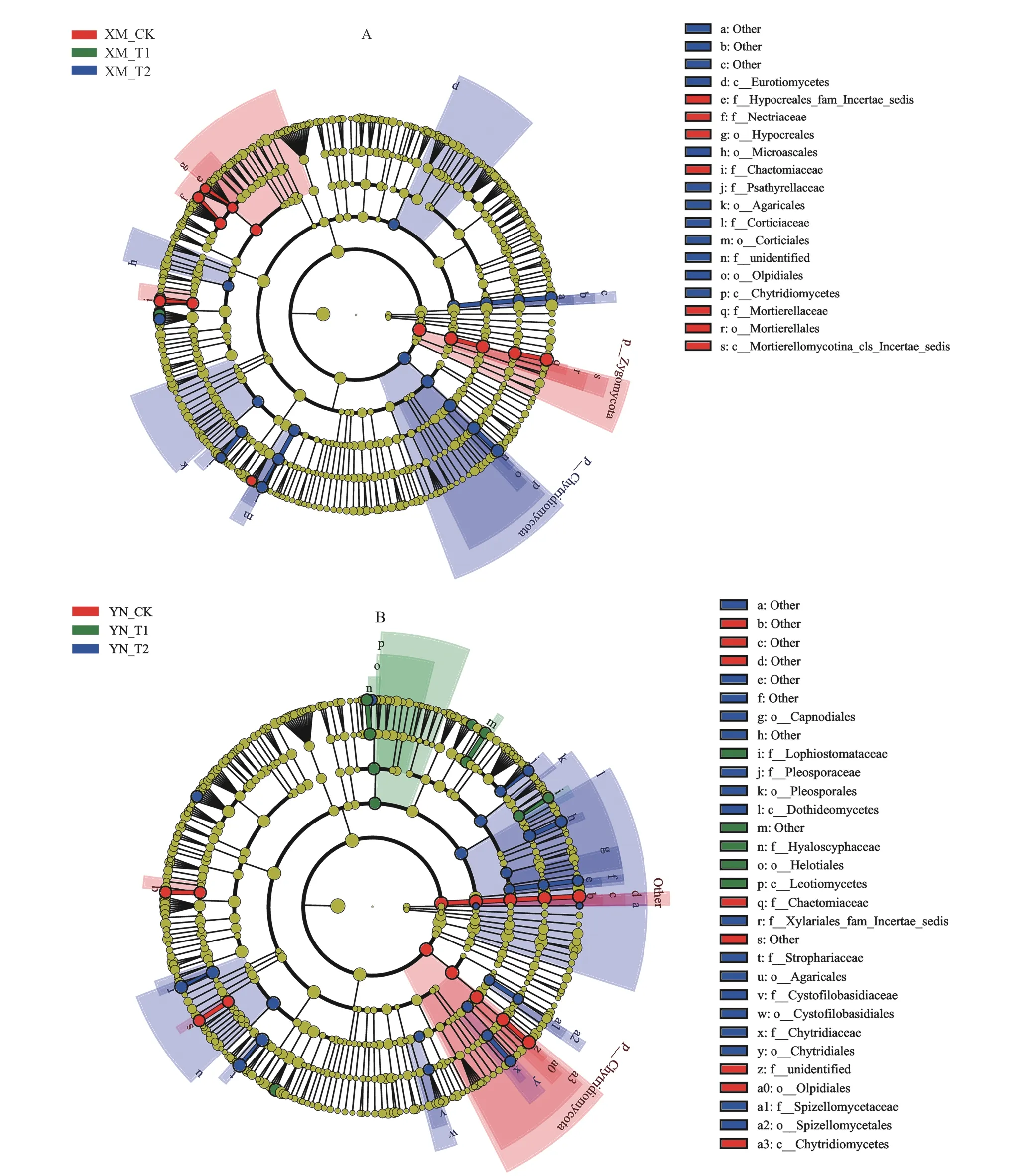
图4 不同倒春寒处理下不同品种小麦的根际土壤真菌LEfSe 物种差异分析Fig.4 Species diversity analysis of LEfSe in rhizosphere soil fungi of different wheat cultivars under late spring coldness
由图4B 可见,YN_CK 富集更多的壶菌纲、毛壳菌科、Olpidiales,YN_T1 富集更多的鸭嘴菌科(Lophiostomataceae)、晶杯菌科、柔膜菌目、锤舌菌纲,YN_T2 富集更多的座囊菌纲、煤炱目、伞菌目、壶菌目、小壶菌目(Spizellomycetales)、Cystofilobasidiales、格孢菌科、囊藻科(Cystofilobasidiaceae)、Xylariales fam Incertae sedis、球盖菇科、壶菌科、Spizellomycetaceae。
2.5 倒春寒胁迫下小麦开花期根际土壤真菌群落FUNGuild 功能预测
真菌FUNGuild 数据库注释结果(图5)显示: 在小麦根际土壤真菌群落中除未分类腐生真菌外,内生真菌(endophyte)的相对丰度最高,在XM_CK、XM_T1、XM_T2、YN_CK、YN_T1、YN_T2 中相对丰度分别为21.16%、20.54%、15.26%、16.70%、16.24%和12.79%。两品种间差异显著且-2 ℃胁迫下XM 和YN 内生真菌相对丰度分别较CK 显著降低27.87%和23.43%。其次为土壤腐生菌(soil saprotroph)、凋落腐生物(litter saprotroph),两功能群在各处理中相对丰度分别为18.76 %、19.07%、13.12%、14.81%、13.15%、19.17%和17.63%、18.52%、11.93%、14.00%、14.03%、9.19%。此外,木腐菌(wood saprotroph)、植物病原体(plant pathogen)在XM_T2 中相对丰度显著高于其他处理,占比分别为9.86%和9.23%。综上,随倒春寒胁迫程度的增强小麦根际土壤中内生真菌的相对丰度显著降低,且XM 降低幅度更大;XM_T2 中木腐菌、植物病原体相对丰度更高。
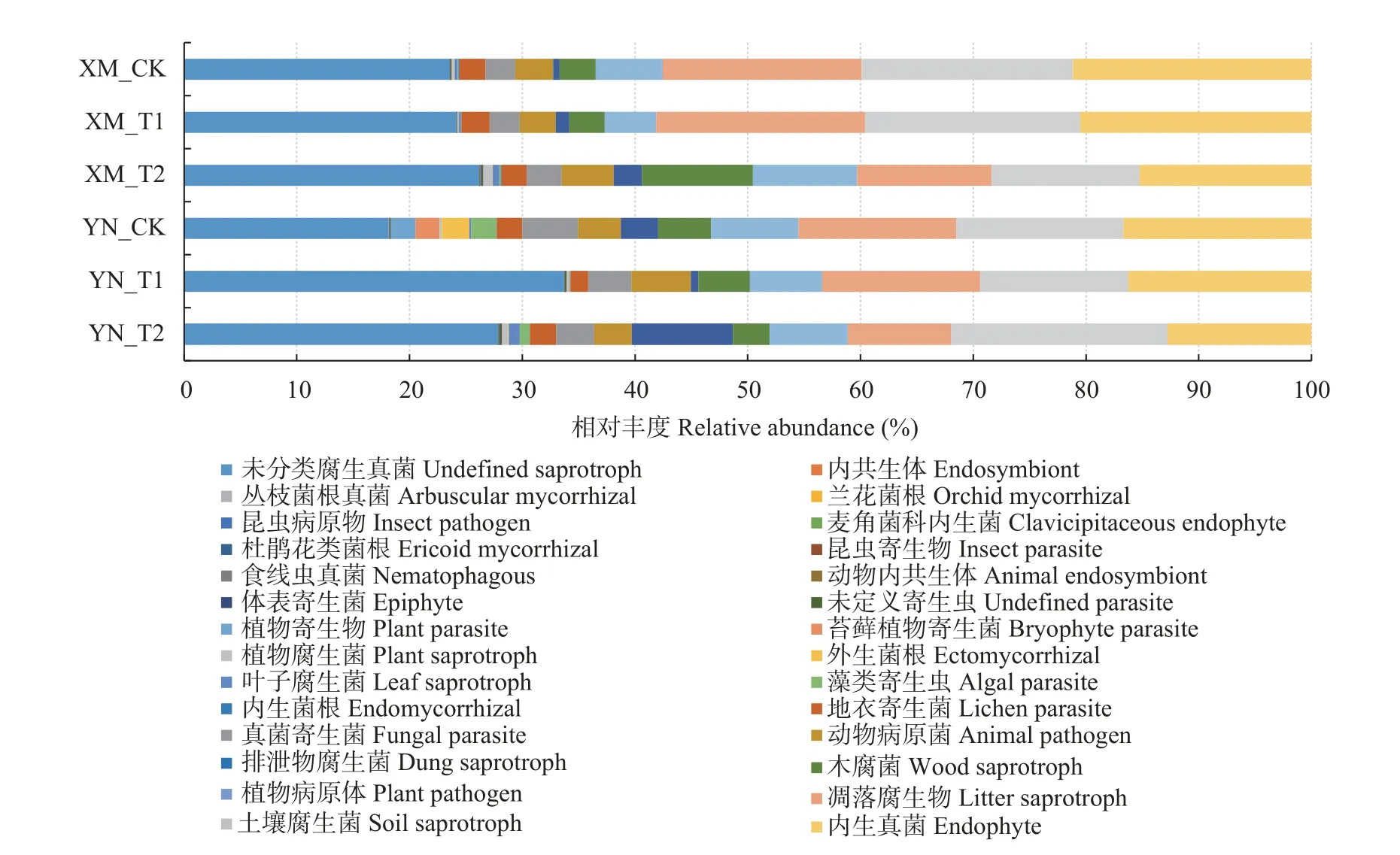
图5 不同倒春寒处理下不同品种小麦的根际土壤真菌 FUNGuild 功能预测Fig.5 FUNGuild function prediction in rhizosphere soil fungi of different wheat varieties under late spring coldness
3 讨论
3.1 倒春寒对开花期小麦植株生长的影响
全球气候变暖背景下黄淮麦区倒春寒频发重发已成为限制该地区小麦高产高质高效发展的重要因素[4],详细了解倒春寒对小麦植株生长发育尤其是对根系的影响对小麦生产的防灾减灾具有重要意义。本研究发现倒春寒会降低开花期小麦根系生物量的积累,这与Muhammad[29]研究结果一致。这可能是因为倒春寒危害降低了小麦根系活力,使根系生长缓慢,生物量积累显著降低[16]。根系是小麦吸收土壤中水分和矿质营养的主要器官,其生长状态会影响小麦地上部植株的生长发育[30]。本研究发现倒春寒显著降低了开花期两品种小麦地上部植株干物质重,这与柯媛媛等[31]研究结果一致。其原因可能是倒春寒不仅会降低小麦根系活力、减缓根系生长[32],影响根系对养分的吸收、同化、积累与转运,同时还会破坏小麦地上部功能叶细胞结构的完整性,导致其光合性能下降[10],影响源器官叶片光合同化有机物的合成积累、转运与分配,最终导致小麦植株地上部生物量积累降低。根系和地上部植株共同构成小麦的植株整体,维持适宜的根冠比不仅能使根系高效利用土壤水分和养分,同时又能控制根系的生长冗余,减少根系生长对物质的消耗[33]。王艳哲等[34]和王宁等[33]研究认为较多光合同化物分配给地上部植株,降低根冠比有利于作物产量的提升。本研究发现倒春寒危害后小麦植株的根冠比显著增加,意味着倒春寒使小麦根系与冠层生长发育失衡,不利于产量的形成。
3.2 倒春寒对开花期小麦根际真菌群落结构的影响
根际是植物根系与土壤间物质与能量转换的直接界面[35],也是植物根系自身生命活动和代谢对土壤影响最直接、最强烈的区域[36]。温度胁迫可以改变植物根系分泌物的组成和含量,诱导植物根系分泌大量的物质进入到根际土壤中,吸引附近土壤环境中的微生物,致使作物根际微生物组成结构发生改变[19,37]。本研究通过Illumina 测序发现药隔期倒春寒胁迫改变了两品种小麦根际土壤真菌群落分类学上的相对丰度,其中子囊菌门和担子菌门是主要的优势菌群,这与前人研究结果一致[38-39]。子囊菌门是农业土壤中最大的真菌类群,XM 和YN 根际土壤中子囊菌门相对丰度均随倒春寒胁迫程度的加重而增加。不同胁迫程度下抗倒春寒性强的品种YN 根际土壤中担子菌门相对丰度均高于抗倒春寒性弱的品种XM,可能是因为担子菌门能与小麦共生形成菌根,增强植株抗性[40]。壶菌纲在XM_T2 根际土壤中的相对丰度与其他处理的根际土壤相比显著增加,这与张江伟等[41]对于逆境胁迫下小麦根际微生物群落变化的研究结果一致。微生物数量与结构的变化影响作物根际土壤环境及根系对土壤养分的吸收和转化,与作物的生长发育密切相关[42]。被孢霉真菌是植物生长促生菌[43],能促进植物产生赤霉素、吲哚乙酸等植物激素及大量多不饱和脂肪酸[44],并为植物提供氮、磷等营养物质[45],促进植物的生长,显著提高植物生产力与抗胁迫能力[46]。前人研究表明脲酶活性与被孢霉菌呈正相关[47],可促进土壤有机质中的氮转化成氨以及尿素氮肥的水解,增强土壤的供氮能力,从而影响微生物群落结构[48]。本研究中两品种小麦根际土壤被孢霉目、被孢霉科、被孢霉属的相对丰度均随胁迫强度的增加呈下降趋势。其原因可能是倒春寒胁迫使根际土壤脲酶活性下降[48],减少对土壤有机质的转化,降低了对微生物养料的供给,因此抑制了小麦根际土壤被孢霉属真菌的生长。
3.3 倒春寒对小麦根际真菌群落多样性的影响
土壤根际微生物的多样性影响着作物生长与抗逆能力[49-50]。根际微生物区系结构合理,多样性和物种丰富度越高时,作物抗逆能力就越强[51]。本研究Alpha 多样性结果分析表明,倒春寒改变了小麦根际土壤真菌物种多样性与丰富度。其原因可能是倒春寒胁迫对小麦植株地上地下部的生长产生了不利影响,从而使得不同处理间小麦根系分泌物发生了改变,进而改变了根际微生物生态环境,并形成与之相适应的真菌群落,复杂的微生物群落可能提高了根际土壤微生物的整体代谢活性,促进了真菌群落结构多样化的形成[18]。大量研究表明,在特定的胁迫环境中作物根系通过分泌一系列物质诱导菌群繁殖[52],并招募能够缓解自身胁迫压力的特定微生物种群聚集于根际[53]。毛梦雪等[54]研究指出植物根系分泌物与植物激素在非生物胁迫下协同作用,提高根际微生物活性,改善植物生长并提高植物胁迫耐受能力。姜丽娜等[55]研究表明一定的低温胁迫下小麦根系分泌更多的可溶性蛋白、可溶性糖及总酚,这些物质能为根际微生物的生长和繁殖提供充足的营养促进其生长[56],影响土壤微生物种类、数量及其在植物根际的分布[57],从而缓解逆境胁迫对小麦造成的损伤。本研究中根际土壤真菌群落物种多样性与丰富度的变化也体现了小麦对环境变化的适应能力。此外,本研究发现随倒春寒胁迫程度增强,抗倒春寒性弱的XM 较抗倒春寒性强的YN 根际土壤中真菌多样性和丰富度变化更大,这可能是因为抗倒春寒性弱的小麦品种应激效应更为剧烈,其根系分泌物中可溶性蛋白、可溶性糖等物质含量增加,为微生物提供更多的底物,促进了微生物的生长繁殖。不同根际分泌物的种类、含量及土壤酶活性会影响根际微生物数量及群落结构,因此后期将进一步研究探讨倒春寒胁迫下根际土壤酶活性的变化及对小麦根系分泌物种类和含量的影响,以及小麦根系活力的动态变化规律,从根土环境出发深入探究“根系-土壤-微生物”交互系统对倒春寒的响应。
4 结论
1)倒春寒降低开花期小麦地上、地下部干物质积累,同时使根冠比升高,其对抗倒春寒性弱的XM影响大于抗倒春寒性强的YN。
2)倒春寒改变了小麦根际土壤真菌的群落结构,使子囊菌门相对丰度显著升高,而被孢霉目、被孢霉科、被孢霉属相对丰度显著降低,且随胁迫程度增强XM 的下降幅度更大。抗倒春寒性强的品种YN 中伞菌纲真菌的相对丰度显著高于抗倒春寒性弱的XM。
3)倒春寒胁迫对小麦根际土壤真菌群落多样性与丰富度影响显著,同时改变不同生态功能群的占比。随倒春寒胁迫程度增强,抗倒春寒性弱的XM根际土壤中真菌多样性和丰富度较抗倒春寒性强的YN 变化更剧烈。
