白花草木樨结瘤缺失型突变体的结瘤表型及生物量分析
2023-10-21王升升段珍周培张吉宇
王升升,段珍,周培,张吉宇
(兰州大学草地农业科技学院,兰州大学草种创新与草地农业生态系统全国重点实验室,甘肃 兰州 730020)
氮元素是所有植物维持正常生长发育不可或缺的生命元素,缺氮会抑制植物生长发育,导致农作物产量下降[1-2]。尽管空气中含有 78%的惰性氮素,但由于活化惰性气体极其困难,氮气难以被大多数植物直接利用,且土壤本身的含氮化合物十分匮乏,严重限制了作物产量[3]。所以在农业生产中经常使用工业氮肥来提升作物产量,但目前,农业生产严重依赖工业化肥的投入,我国平均每年的氮肥使用量为 305 kg·hm-2,氮的利用效率只有0.25,然而全球的氮利用效率为 0.42[4-6],且农业土壤中积累的工业氮肥对环境和人类健康产生了诸多危害,包括土壤大面积酸化、地下水污染、氧化亚氮等温室气体的积累[6]。为了实现绿色农业的可持续发展,提高作物生产的氮利用效率,亟须在农业生产中找到可持续的绿色氮素供应方式来代替工业氮肥[7]。
生物固氮是地球生态系统中氮素来源最主要的形式,全球有 70%的结合态氮产自生物固氮,地球上每年通过生物固氮可生产 2 亿t 的氮素量[7],其中最主要的来源是豆科(Leguminosae)植物与根瘤菌的共生固氮[3]。豆科植物种类众多,其成员遍布全球[8],大多数豆科植物能与根瘤菌形成共生固氮关系,在豆科植物根部形成根瘤器官,将空气中的氮气转换为可供植物利用的氨态氮[9]。豆科植物与根瘤菌的共生固氮使得豆科植物能够在氮匮乏的环境中正常生长发育[10],同时能够向土壤中输入大量的氮、磷、铁等营养元素,从而改善土壤质量[11],使得豆科植物成为了全球生态系统中的重要物种,而豆科植物与根瘤菌的共生固氮关系成为最有潜力的可持续绿色氮素供应方式。
豆科植物与根瘤菌之间通过复杂的信号交流形成共生固氮关系,目前在豆科模式植物中通过正反向遗传学发现了 200 多个调控共生固氮的基因[12],一些关键基因的突变和缺失将导致侵染线和根瘤的形成及发育受阻。例如,百脉根(Lotus japonicus)LjNUP85基因的突变体,与根瘤菌共生后无法形成正常的根瘤[13];蒺藜苜蓿(Medicago truncatula)中MtLIN基因的突变导致无法形成侵染线[14]。其中,通过正向遗传学发现的基因对于豆科植物与根瘤菌的共生固氮影响最大,在豆科模式植物中通过甲基磺酸乙酯(ethylmethane sulfonate,EMS)诱变产生了大量共生固氮相关的突变体[12,15],按照表型分为不能形成根瘤或侵染线的结瘤缺失型突变体[16]和能形成白色或绿色等小根瘤但不能进行固氮或固氮能力显著下降的固氮缺失型突变体[17]。这些突变体被用于共生固氮体系的建立和宿主结瘤基因的研究中,发现了参与共生信号通路的关键结瘤基因[18]。但豆科植物与根瘤菌的共生固氮过程非常复杂,根瘤菌侵入宿主根细胞后建立共生关系的许多过程仍然未知,所以亟须挖掘新的共生固氮基因。
草木樨(Melilotus)是一年生或两年生的二倍体豆科草本植物[19-20],白花草木樨(Melilotus albus)是该属最常见的生产利用种之一[21]。草木樨固氮能力强,能够改善土壤质量,被用于农业生产作物轮作系统[22],且已通过连续轮回选育,育成了草木樨低香豆素新材料[23-24]。但目前对草木樨与根瘤菌共生固氮的机制研究较少,本研究以白花草木樨野生型Ma389 经EMS 诱变后的3 种结瘤缺失型突变体(Ma58、Ma61、Ma62)为材料[25],在前期的研究基础上[26],对3 种结瘤缺失突变体植株的结瘤表型和生物量进行分析,并对突变体Ma62 的根瘤进行切片观察,为进一步阐明豆科植物与根瘤菌的共生固氮机制奠定基础。
1 材料与方法
1.1 试验材料
1.1.1 植物材料 白花草木樨野生型Ma389,野生型经EMS 诱变产生的突变体Ma58、Ma61 和Ma62 均由加州大学洛杉矶分校的Ann M. Hirsch 教授友情提供。2022 年4 月在兰州大学草地农业科技学院智能温室(16 h 光照26 °C/8 h 黑暗18 °C)内种植培养。
1.1.2 菌株 本试验使用的根瘤菌是苜蓿中华根瘤菌(Sinorhizobium meliloti,SM1021)。
1.1.3 培养基 TY 培养基(1 L):蛋白胨5.0 g,酵母提取物3.0 g,无水CaCl₂ 0.9 g,pH=7.4(固体培养基加15.0 g 琼脂)。1×PBS 液体(1 L):NaCl 8.0 g,KCl 0.2 g,Na2HPO41.44 g,KH2PO40.24 g,pH=7.4。MS 营养液(1 L):MS 培养基(Solarbio)4.74 g。无氮营养液(1 L):改良霍格兰营养液(无氮、不含硝酸钙)(海博生物技术有限公司) 0.635 g。
1.2 试验方法
1.2.1 材料种植条件 挑选颗粒饱满的野生型Ma389 和突变体Ma58、Ma61 和Ma62 的种子,用砂纸破除硬实后,使用 75%的乙醇处理30 s,5%的次氯酸钠溶液处理5 min 进行表面消毒[22]。将处理好的种子放入铺有两层滤纸的培养皿中,加入少量蒸馏水后用锡箔纸外包放入4 ℃冰箱春化2 d,再放到25 ℃培养箱(16 h 光照/8 h 黑暗)萌发5 d 后,分别移至富氮和低氮基质的花盆中生长,生长一周后在植物根部接种根瘤菌。富氮基质:灭菌蛭石(MS 营养液浇灌),低氮基质:灭菌蛭石(自来水浇灌),富氮基质和低氮基质浇水次数保持一致,每组试验设置5 个生物学重复,每隔10 d 观察记录植株生长性状,直至观察至接种后50 d 为止。
1.2.2 根瘤菌培养和接菌 取保存于-80 ℃冰箱的SM1021 菌株,在超净工作台中用灭菌的接种环在固体TY 培养基上进行划线,30 ℃倒置培养3~4 d,挑取单菌落于10 mL 液体 TY 培养基的离心管中,200 r·min-1下培养 2 d,按照菌液和液体 TY 培养基比例为 1∶200 的体积进行扩繁,30 ℃摇床下培养 36 h。
使用分光光度计(UV7558 紫外可见分光光度计,上海佑科仪器仪表有限公司)和灭菌双蒸水将菌液浓度调制 OD600=0.1,使用注射器在每株草木樨的根部注射10 mL 进行接菌。
1.2.3 根瘤切片观察 取野生型Ma389 和突变体Ma62 的根瘤于1.5 mL 离心管中,加入1 mL 1×PBS 液体保存。用1 mL 2.5%~4.0%的沃尔醛溶液替换1×PBS 液体后抽真空30 min,放于4 ℃冰箱过夜,用1×PBS 液体洗3 次,间隔10 min,再依次用10%,20%,30%,50%,70%,80%,90%,100%的酒精进行替换,间隔40 min(用70%的酒精替换后可以保存较长时间,放于 4 ℃冰箱),100%的酒精替换4 h 后,再依次用无水乙醇∶树脂=2∶1,无水乙醇∶树脂=1∶2,全树脂进行替换,间隔2~4 h。
配置树脂∶凝固剂=15∶1 的液体加到包埋板中,每个小孔400 mL,将替换到全树脂的根瘤,转移到包埋板的小孔中,尽可能将其放在底部,封上合适大小的封口膜,等其凝固进行切片后,用 TBO 染色剂染色3 min,用双蒸水冲洗干净(所有切片染色程度应相同),晾干后在体视显微镜(leica zoom 2000TM,上海佑科仪器仪表有限公司)下观察,并拍照记录。
1.3 数据处理
使用Microsoft Excel 2010 对所有数据进行统计分析,以计算平均值和标准误差。
2 结果与分析
2.1 野生型与3 种突变体的结瘤表型鉴定
为了观察白花草木樨野生型Ma389 及其突变体Ma58、Ma61 和Ma62 的结瘤表型,将萌发的种子移至装有蛭石的花盆中,用无氮营养液进行浇灌,生长一周后接种苜蓿中华根瘤菌SM1021。取接种3 d 后的野生型与3 种突变体的根在体视显微镜下观察根毛形态,以未接种根瘤菌的根作为对照。本研究检测到野生型Ma389 接种根瘤菌后的根毛呈显著弯曲变形(图1 A,E),而突变体 Ma58(图1 B,F)中观察到根毛在接种后显示出近端膨胀和轻微弯曲的变形情况,突变体Ma61(图1 C,G)和Ma62(图1 D,H)的根毛在接种后仅显示出顶端稍微肿胀的有限变形情况,结果表明相比于野生型Ma389,突变体Ma58,Ma61 和Ma62 接种根瘤菌后根毛显示出异常和有限的变形情况,推测这3 种突变体的共生信号传导在早期阶段可能就被阻断。
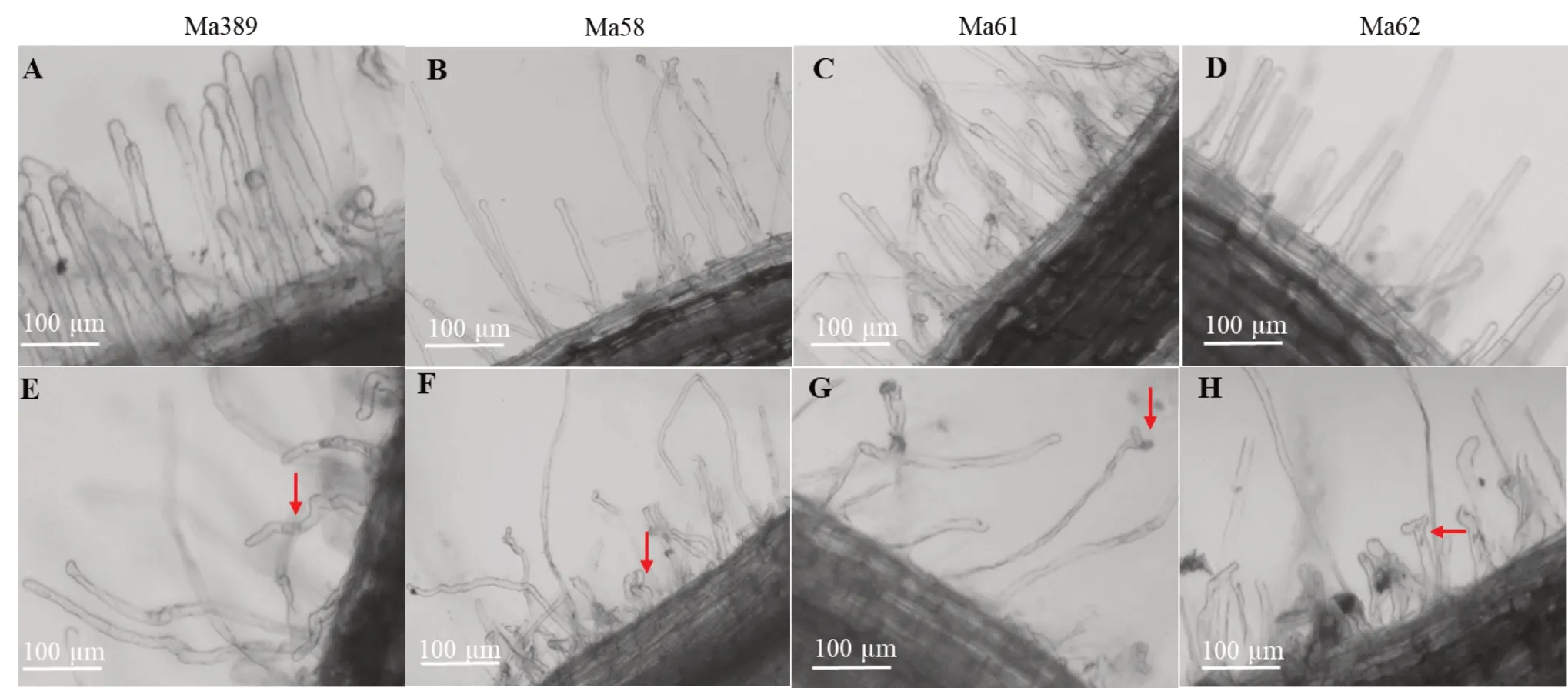
图1 野生型与3 种突变体的根毛接种苜蓿中华根瘤菌后的变化Fig.1 Responses of root hairs from wild-type and three mutants inoculated with SM1021
接种30 d 后观察野生型与3 种突变体的结瘤表型,野生型Ma389 能够形成正常的粉色根瘤(图2 A),平均每株形成21 个根瘤(图3)。突变体Ma58、Ma61 不形成根瘤,甚至不形成根瘤原基(图2 B,C)。突变体Ma62 能够形成少量的白色根瘤(图2 D),平均每株形成5 个根瘤(图3),根瘤大小远小于野生型。
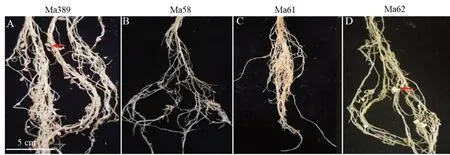
图2 野生型与3 种突变体植株30 d 时的根部结瘤表型Fig. 2 Root nodulation phenotypes of wild-type and three mutants at 30 days

图3 野生型与3 种突变体植株30 d 时的根瘤数Fig. 3 Nodule number of wild-type and three mutants at 30 days
2.2 野生型Ma389 与突变体Ma62 根瘤细胞的切片观察
为了进一步观察野生型Ma389 与突变体Ma62 的根瘤细胞,取Ma389 和Ma62 接种根瘤菌后30 d 的根瘤进行切片染色观察。研究发现与野生型Ma389(图4 A,C)相比,突变体Ma62 的根瘤细胞显示没有根瘤菌的定殖(图4 B,D),表明由于突变体Ma62 在接种后根毛中没有形成侵染线,根瘤菌进入根瘤器官的途径被阻断,导致突变体Ma62 的根瘤细胞中没有定殖根瘤菌,这与前期的研究结果一致[26]。突变体Ma62形成的少数白色根瘤为无效根瘤,无法利用空气中的氮源为植物提供氮素营养。

图4 野生型Ma389 与突变体Ma62 的根瘤切片表型Fig.4 Root nodule section phenotype of wild-type Ma389 and mutant Ma62
2.3 野生型与3 种突变体的生物量分析
在低氮基质中生长的白花草木樨野生型和3 种突变体,在接种根瘤菌后生长至30 d 时,3 种突变体的地上部鲜/干质量、株高和根长与野生型相比无显著差异,而突变体Ma58、Ma61 的根鲜/干质量显著小于野生型(P<0.05),Ma62 的根鲜/干质量与野生型相比无显著差异。植株生长至40 d 以后,3 种突变体的各性状均低于野生型(P<0.05),其中突变体Ma58、Ma61 的地上部鲜/干质量显著低于野生型(P<0.01)(图5)。结果表明,在低氮基质中生长时,由于野生型Ma389 在接种根瘤菌后能够形成正常的根瘤,可以利用空气中的氮气为植物提供氮源来维持植物正常生长,但突变体Ma58,Ma61 和Ma62 在接种根瘤菌后无法形成根瘤或只能形成少量的白色无效根瘤,没有固氮能力,无法提供植株生长所需的氮源(图6)。

图5 低氮基质中野生型与3 种突变体植株的生物量Fig.5 Biomass of wild-type and three mutants in low nitrogen substrate

图6 低氮基质中野生型与3 种突变体植株30 d 时的表型Fig.6 Phenotypes of wild-type and three mutants at 30 days in low nitrogen substrate
在富氮基质中生长的白花草木樨的野生型和3 种突变体,在接种根瘤菌后生长至50 d 时,3 种突变体的地上部鲜/干质量、根鲜/干质量、株高和根长与野生株相比无显著差异(图7)。结果表明,在富氮基质中生长时,虽然突变体Ma58,Ma61 和Ma62 无法通过自身固氮为植株提供氮源,但能够通过根系吸收基质中的氮源来维持植物正常的生长发育,所以突变体植株的各性状与野生型相比无明显差异(图8)。这说明突变体Ma58,Ma61 和Ma62 的突变基因只与根瘤菌的共生固氮机制相关。

图7 富氮基质中野生型与3 种突变体植株的生物量Fig.7 Biomass of wild-type and three mutants in rich nitrogen substrate
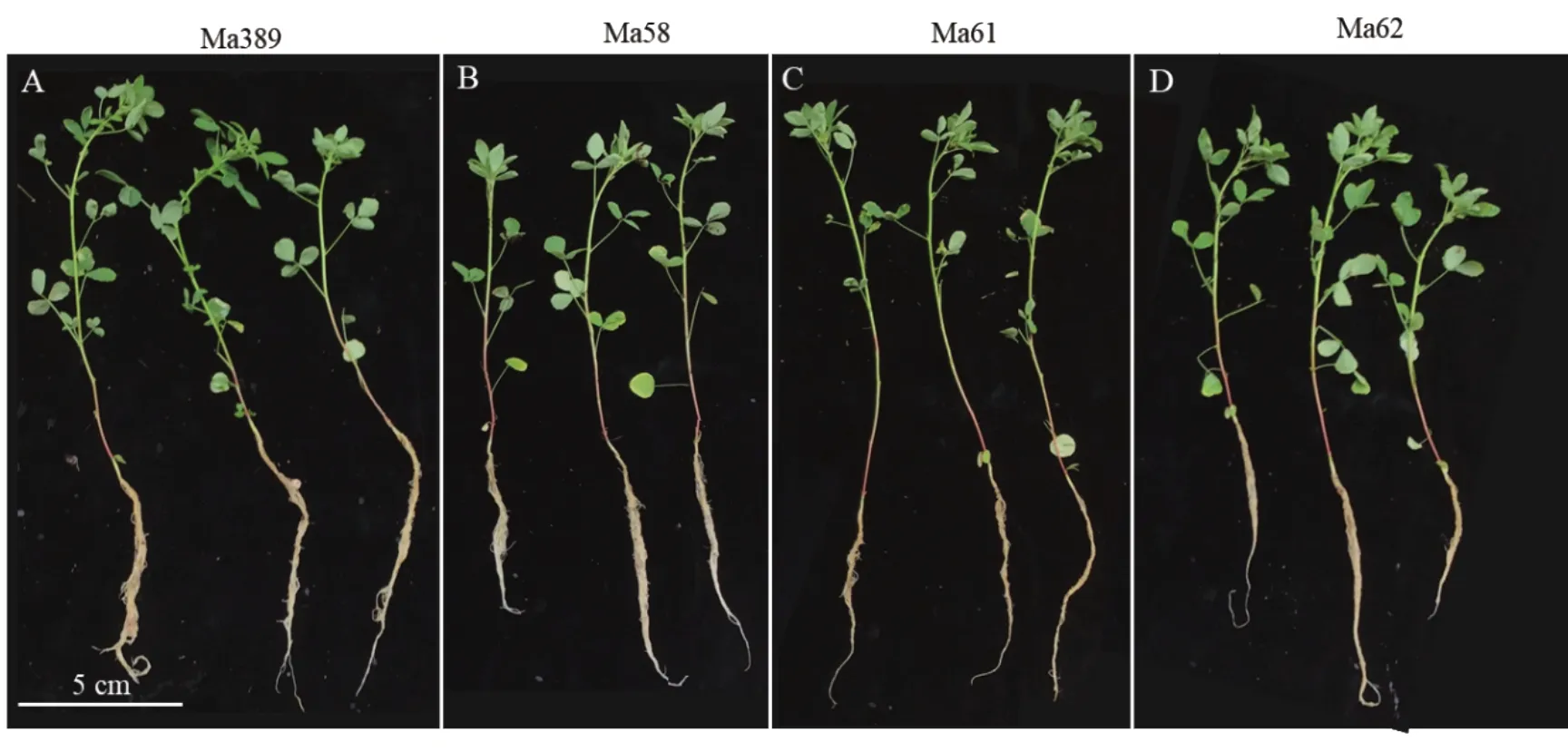
图8 富氮基质中野生型与3 种突变体植株30 d 时的表型Fig.8 Phenotypes of wild-type and three mutants at 30 days in rich nitrogen substrate
3 讨论
3.1 豆科植物与根瘤菌共生固氮关系的建立
豆科植物与根瘤菌之间通过一系列复杂的信号识别和相互作用形成根瘤,这个过程主要分为侵染线的形成和根瘤器官的形成两个部分[12]。首先,在低氮条件下豆科植物根际释放的类黄酮化合物会吸引土壤中的根瘤菌富集在植物根部并诱导其分泌结瘤因子(nod factor,NFs)[27],豆科植物根部表皮细胞膜上的结瘤因子受体(nod factors reception,NFRs)会特异性识别和结合NFs,引起植物根毛顶端肿胀和根毛弯曲变形[28],弯曲的根毛内包裹根瘤菌细胞形成侵染点,并随着局部细胞壁的降解和细胞膜的凹陷不断伸长,形成内含根瘤菌的侵染线[29]。同时,根瘤菌刺激植物根皮层细胞不断分裂,在根毛上局部性体积膨大后形成根瘤原基,根瘤菌通过在侵染线中不断增殖运动到根瘤原基,最终发育为具有固氮能力的成熟根瘤[30]。
目前在豆科模式植物中建立了共生固氮早期信号传导模型[12],豆科植物的NFRs 在感知到根瘤菌释放的NFs 后会将结瘤信号从胞外向胞内传递到下游,NFRs 一般为LysM 型受体样激酶,例如蒺藜苜蓿中的LYK3、NFP和百脉根中的NFR1、NFR5[31],二者形成异源二聚体后共同激活共生受体样激酶基因(MtDMI2和LjSYMRK)的表达[16],并引起钙离子振荡反应,核孔蛋白(LjNUP85、LjNUP133)和阳离子通道蛋白(LjPOLLUX、LjCASTOR、MtDMI1)参与钙离子信号从细胞膜内传递到细胞核内[32],而细胞核内的钙和钙调依赖的蛋白激酶(LjCCaMK、MtDMI3)能够解析钙离子信号[33],再将信号传递到下游的转录因子NSP1 和NSP2,二者相互作用激活nin 的表达[34]。
基于豆科模式植物蒺藜苜蓿和百脉根生长周期短,均为二倍体植物,基因组都较小,并建立了较为完善的遗传转化体系等优势,豆科植物与根瘤菌的共生固氮的理论研究目前主要集中在蒺藜苜蓿和百脉根中[35]。但草木樨作为豆科优良牧草,也拥有生长周期短,二倍体植物等优点,且本课题组已完成了草木樨的基因组测序工作[36],建立了发根农杆菌介导的毛状根转化技术[24],符合基础理论研究的条件,但草木樨与根瘤菌的共生固氮基因的研究尚未见报道。
3.2 正向遗传学筛选的豆科植物共生固氮相关突变体
正向遗传学是指生物通过大规模自发或人工随机诱变,通过从发育异常的突变个体中筛选相关的表型和性状,再寻找到突变的基因并研究其功能[15]。在豆科植物与根瘤菌共生固氮的正向遗传学研究中,豆科模式植物通过EMS 诱变产生的与共生固氮相关的突变体,按照表型主要分为两种根瘤异常突变体,一种是不能形成根瘤原基,或不能形成正常的侵染线,根瘤菌侵染在早期阶段被阻断,称为结瘤缺失型突变体[16],例如symrk突变体在接种根瘤菌后根毛会出现膨大的现象,但是没有根毛弯曲的表型无法形成根瘤[16],nin突变体不能形成侵染线和根瘤原基[37]。一种是该突变体能够形成白色或者绿色等的小根瘤,或形状发育异常的根瘤,根瘤中类菌体不能分化或者已经死亡,导致其固氮能力显著下降或者不能进行固氮,称为固氮缺失型突变体[17],例如mtlin突变体只能形成少数白色的根瘤[14]。豆科植物与根瘤菌的共生固氮早期信号传导模型中起关键作用的基因,大多都是在经过正向遗传学筛选的突变体中发现的[35]。
本研究中,使用白花草木樨野生型Ma389 和经EMS 诱变产生的突变体Ma58,Ma61,Ma62,接种根瘤菌后,突变体Ma58 的根毛出现近端肿胀和轻微变形的现象,突变体Ma61 和Ma62 的根毛仅表现出顶端稍微肿胀的有限变形情况。对3 种突变体接种根瘤菌后30 d 观察到,突变体Ma58,Ma61 不产生根瘤,甚至没有根瘤原基,推测这两种突变体中调控根瘤器官形成的信号通路在开始阶段就被阻断,突变体Ma62 只形成少数白色的小根瘤,根瘤数量和大小远远小于野生型Ma389。对突变体Ma62 的根瘤进行切片观察发现,其根瘤中没有根瘤菌的定殖。前期研究也表明这3 种突变体均没有侵染线的形成[24-25],因此这3 种突变体均属于结瘤缺失型突变体。分别观察在低氮和富氮基质中白花草木樨野生型和3 种突变体的生物性状,发现在低氮基质中突变体Ma58 和Ma61 在生长30 d 时,根鲜/干质量显著小于野生型(P<0.05),而突变体与野生型相比Ma62 无显著差异,推测Ma62 经EMS 诱变程度相比于Ma58 和Ma61 较低,但3 种突变体生长至40 d 以后,地上部鲜/干质量、根鲜/干质量、株高和根长等性状均显著低于野生型(P<0.05)。而在富氮基质中3 种突变体植株的各性状与野生型相比无显著差异,这说明突变体Ma58,Ma61 和Ma62 中的突变基因只与共生固氮相关。
3.3 白花草木樨结瘤缺失型突变体Ma58,Ma61 和Ma62 研究前景分析
本研究表明白花草木樨突变体Ma58,Ma61 和Ma62 为结瘤缺失型突变体,且突变基因只与共生固氮相关。因为突变体Ma58,Ma61 不形成根瘤原基和侵染线,突变体Ma62 不能形成侵染线,且只能形成少数白色根瘤,所以推测这3 种突变体的突变基因在共生固氮信号传导早期阶段。为了进一步确定这3 种突变体中与共生固氮相关的突变基因,后期计划将这3 种突变体分别与白花草木樨野生型Ma389 进行杂交,并以突变体作为母本,野生型Ma389 作为父本,获得F1代后,F1代经自交产生F2代,观察F2代植株的结瘤表型分出突变体表型的单株,采集突变体表型的植株样品和野生型的样品分别构建DNA 混池,以草木樨基因组为参考基因组[36],通过混合群体分离分析法(bulked segregant analysis,BSA)重新测序定位草木樨生物固氮相关基因,筛选候选区域中有SNP 或Indel 的基因,通过发根农杆菌介导的毛状根转化技术进行突变基因的功能验证,以此来挖掘新的共生固氮基因,为进一步阐明共生固氮早期信号传导途径提供新的理论依据。
4 结论
本研究对白花草木樨野生型Ma389 和经EMS 诱变产生的突变体Ma58,Ma61,Ma62 在接种根瘤菌后的表型和生物量进行了观察和分析,结果显示突变体Ma58,Ma61 无法形成侵染线和根瘤原基,突变体Ma62 不能形成侵染线,只形成不固氮的白色小根瘤,表明这3 种突变体均属于结瘤缺失型突变体。在低氮基质中生长的突变体植株在 30 d 开始各生物性状均显著低于野生型,而在富氮基质中则无显著差异,表明突变基因只与共生固氮相关,且突变基因定位在共生固氮信号传导早期阶段。白花草木樨突变体与根瘤菌共生的表型鉴定和生物量分析为豆科植物共生固氮机制研究提供了重要参考,也为草木樨种质利用和遗传育种提供了科学依据。
